





(C) 2010 William A. Shear. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The genus Trilasma Goodnight & Goodnight, 1942 is reinstated for Mexican ortholasmatines, and Cladolasma Suzuki, 1963 is reinstated for two species from Japan and Thailand, Cladolasma parvula Suzuki, comb. n. and Cladolasma angka (Schwendinger & Gruber), comb. n. Eight new species in the subfamily Ortholasmatinae Shear & Gruber, 1983 are described, as follows: Ortholasma colossus sp. n. is from California, Trilasma tempestado sp. n., Trilasma hidalgo sp. n., Trilasma trispinosum sp. n., Trilasma ranchonuevo sp. n., Trilasma petersprousei sp. n. and Trilasma chipinquensis, sp. n. are from México, and Trilasma tropicum sp. n. from Honduras, the farthest south for a dyspnoan harvestman in the New World. A new distribution record for Martensolasma jocheni Shear 2006 is given. The recently described Upper Cretaceous amber fossil Halitherses grimaldii Giribet & Dunlop 2005 is not a member of the Ortholasmatinae, but is likely a troguloidean of an undiagnosed family.
Ortholasma, Dendrolasma, Trilasma, Cladolasma, Halitherses, amber, fossil, California, Sierra Nevada, Nuevo León, Tamaulipas, Hidalgo, Veracruz, Honduras, Japan, Thailand, new species, new combination
The harvestman subfamily Ortholasmatinae was monographed by
Since 1983, there has been little activity in the study
of these most unusual harvestmen, but a few years after the 1983
monograph’s publication,
Ortholasmatines have received some attention in studies of harvestman phylogeny.
The first Mexican species of ortholasmatine, previously Ortholasma (Trilasma) bolivari (Goodnight & Goodnight 1942), was placed by
I also describe in this paper a new Californian species from the Sierra Nevada, including Sequoia National Park. Ortholasma colossus, new species, occurs at the southern end of the Sierra Nevada distribution of Ortholasma levipes Shear & Gruber 1983, and a single specimen of the new species was mentioned and briefly characterized in Shear and Gruber’s monograph. The collection of much additional material makes possible the description of this, the largest of all ortholasmatines, nearly half again as long as the previously known largest species.
Though
Ortholasmatines are the most southerly occurring members of the harvestman suborder Dyspnoi, which appears to be limited to the northern hemisphere. In many cases these harvestmen are clearly associated with boreal regions or relatively high elevations, and indeed such is the case with ortholasmatines, though they seem tolerant of drier habitats so long as temperatures are moderate to low. With the exception of Ortholasma pictipes Banks, 1911, and the species of Dendrolasma, which are found in moist coniferous forests from northern California to Alaska, North American Ortholasma species are often characteristic of sclerophyll vegetation (though collecting notes suggest they are found even there in moister microhabitats, such as caves and canyons). There may be additional species awaiting discovery in such habitats in southern California, and the diversity of the subfamily in México may be significantly greater than presently documented. Found near sea level in northwestern North America, this proclivity for cool temperature regimes carries ortholasmatines to high elevations in México, where Trilasma species are found in coniferous forests mostly above 2100 m (7000’) elevation, or in cloud forests. Even at these elevations, collection data suggest that some tropical and subtropical species are troglophiles, seeking out the damper, cooler conditions in caves. In the Sierra Nevada of California, individuals are often taken in caves, but show no troglomorphic adaptations, and while two of the new species described below from México are from caves and are conspicuously long-legged, they do not appear to be troglobionts. The caves have been much more fully explored than their surface environs, so the number of records of ortholasmatines from caves in California and México is probably more an artifact of collecting than an indicator of habits.
Is the fossil Halitherses grimaldii an ortholasmatine?The cuticular ornamentation of Halitherses grimaldii
consists of low, rather rounded, densely scattered tubercles quite
unlike the anvil-shaped, T-shaped, or multi-armed tubercles found in
ortholasmatines, and indeed are more like the ornament of the
ischyropsalidoid genus Hesperonemastoma.
There is no organization of the ornamentation into keel cells, as seen
in all ortholasmatines, even the single species lacking the median
hood process. Likewise, lateral hood processes, universal in all
ortholasmatines, are not seen in the fossil species. It is not clear
what the authors mean by “fused meso- and metapeltidia” since it is
obvious from their illustrations that at least the metapeltidium is
free. The condition of the specimen and the angles of observation
available make it difficult to see. No troguloideans have a free
mesopeltidium, this “first thoracic segment” always being fused to the
carapace, or propeltidium, and the free metapeltidium with scutum parvum is found in several troguloideans (
Giribet and Dunlop included Halitherses grimaldii in the morphological dataset of Giribetet al. (2002), and the result in a strict consensus cladogram was simply to group the species with all other included troguloids in a “comb” (fig. 6 of Giribet & Dunlop 2005). This was undoubtedly due to much missing data for the fossil species, but also showed no close relationship to Nemastomatidae.
All the evidence suggests to me that Halitherses grimaldii is not an ortholasmatine. The large eyes and eye tubercle are unique, and among troguloids, only the cavernicolous nemastomatids have such extraordinarly long legs. I think it highly likely that this species represents at the least a new, probably extinct, family within Troguloidea.
Palpal setaeTwo distinct types of palpal setae in the Dyspnoi,
not necessarily homologous, have been confused in some recent
literature, where both are referred to as “clavate” hairs or setae. In
the superfamily Troguloidea, the palpal setae are of the type described in great detail by
However, the palpal setae in some species of the superfamily Ischyropsalidoidea,
illustrated by me in 1986 using scanning electron micrographs, are
quite different. In these setae, fine tubules occupy at least the
distal half of the seta, are not arranged radially, and may or may not
be the source of a secretion. Similar setae are often found on the
palpi of both juvenile and some adult Eupnoi (
Details need to be examined before these setae are used further as characters in phylogenetic reconstructions. For example, both types of hairs appear to be absent at least in adults of the ischyropsalidoid genera Ischyropsalis, Ceratolasma and Acuclavella. Crosbycus dasycnemus plumose setae appear different from those of Taracus or Sabacon; the former are truncate, not pointed, and the tubules appear not to extend all the way to the tip.
Chemical defensesSeveral specimens of Ortholasma pictipes and Ortholasma rugosum
were dropped alive into small amounts of methanol, and the extract
analyzed for volatile components. This method has proven itself as a way
of determining the presence and nature of the chemical defenses of
harvestmen (
Recently,
Measurements of the appendages of single specimens were taken from material temporarily mounted in glycerine on microscope slides, using a calibrated ocular micrometer in an Olympus BX-50 compound microscope. Lengths and widths of appendage segments were taken from their greatest lengths and greatest widths; length of first cheliceromere includes the apodeme, that of the second cheliceromere the fixed finger. Total leg lengths do not include coxae or trochanters, and are the summed lengths of the femora, patellae, tibiae, metatarsi and tarsi. Body dimensions were measured under an Olympus SZH stereomicroscope, using an external scale and camera lucida attachment, and photomicrographs were taken using this microscope and a mounted Luminera Infinity 1 digital camera; photographs were manipulated for clarity in the programs PhotoShop and IPhoto. The animals in these photographs appear red or orange rather than their field color of brown or black because the use of transmitted light in taking the photographs enhanced the clarity of the keel cells, but incidentally produced the colors; removing the color results in less clarity.
Total length was measured from the distal tip of the median hood process to the posterior margin of the abdominal scute, excluding the extended keel tubercles; total width at the widest point of the body (usually near the posterior margin of the scute). Length of the median hood process was measured in dorsal view from the keel just posterior to the eye tubercle to the distal tip, and width was measured at the widest point. All measurements are in millimeters. Tarsal article counts vary, sometimes on the left and right sides of the same individual, but usually only by one or two articles. The counts given in the descriptions below should be taken as possibly varying by one or two in either direction.
Map coordinates and approximate elevations were obtained from Google Earth when the data did not appear on collection labels.
Depositories:TMM Texas Memorial Museum
AMNH American Museum of Natural History
CAS California Academy of Sciences.
TaxonomyFamily Nemastomatidae Simon, 1872
Nemastomatid Opiliones with the anvil-shaped tubercles on the dorsum exaggerated, with long horizontal arms overlapping in patterns to form distinctive keel cells (Figs 2, 3); lateral anterior margins of the carapace with at least one (up to three) forward-projecting lateral hood process on either side of the eye tubercle; eye tubercle in all but one genus with long, forward-projecting median hood process bearing large T-shaped tubercles laterally and sometimes dorsally. All these characters are absent from nemastomatines.
.
| 1. | With a forward-projecting process on the eye tubercle, bearing elongate T-shaped tubercles laterally | 2 |
| – | Without such a process, eye tubercle with small, blunt dorsal spine; Aguascalientes, Puebla, México | Martensolasma jocheni Shear |
| 2. | Frontal border of the carapace with a single long process on either side of the eye tubercle | 3 |
| – | Frontal border of the carapace with two or more such processes | 4 |
| 3. | Metapeltidium separated from both carapace and abdominal scutum; keel cells small, not in transverse rows; southern Japan and SE Asia | Cladolasma Suzuki |
| – | Metapeltidium fused to abdominal scutum, partially fused to carapace; keel cells larger, arranged in approximate transverse rows; northern Pacific coast of North America | Dendrolasma Banks |
| 4. | Dorsal hood process with median armed tubercles; scute with rows of small cells between rows of large ones; males without a gland on first cheliceromere; central and northwestern México to Honduras | Trilasma Goodnight & Goodnight |
| – | Dorsal hood process without median armed tubercles; only large cells on scute; males with dorsal gland on first cheliceromere; western North America from southern California to northern British Columbia | Ortholasma Banks |
Martensolasma jocheni
Ortholasmatines lacking a median hood process on the eye tubercle and having a simple pattern of keel cells; scutum magnum present.
24 km north of Xicotepec de Juarez, 1070 m asl, oak forest litter, Puebla, México, collected 17 June 1983 by R. Anderson, male (AMNH).
This specimen is virtually identical to the holotype and paratype males from Aguascalientes, Aguascalientes, México, despite having been collected about 480 km (300 miles) east-southeast of there. The Aguascalientes type specimens came from a garden, and I speculated that they might have been introduced, but then only considered introductions from outside México. While it is possible (though unlikely) that the species has a natural range of this size, the Aguascalientes specimens could have been brought to that major urban center with garden plants from Puebla. Future collecting may solve the dilemma, and may also produce the presently unknown females.
Cladolasma parvula
Ortholasmatines with single lateral hood processes on each side of eye tubercle (two or more such processes in Ortholasma and Trilasma), two-thirds or more length of median hood process; metapeltidium free from abdominal scutum (fused to scutum in Dendrolasma); scute ornamentation of small keel cells only (large or small cells in Dendrolasma); leg femoral microsculpture of broad denticles. Penis with short stylus, two lateral rows of macrosetae.
In addition to the type, Cladolasma angka (Schwendinger & Gruber 1992).
Southern Japan; northern Thailand.
While the male of Cladolasma angka is unknown,
With regard to Dendrolasma, the two species now included there are quite different from one another. Dendrolasma mirabile
Ortholasma rugosum
Ortholasmatines in which the median hood process arises rostrally and is distinctly spoon-shaped, widest past its midlength, and lacks a median, dorsal row of T-shaped tubercles (Trilasma species have parallel-sided median hood processes with a median row of tubercles); two lateral hood processes; males with glands dorsally on the first cheliceromere (glands lacking in Trilasma); male first cheliceromere with or without distal inner tooth; males of some species with palpal femoral and patellar glands (needs verification for all species using scanning electron microscopy); keel cells large, not subdivided into smaller cells (some cells subdivided in Trilasma; sometimes in Dendrolasma).
Ortholasma species are best distinguished from one another by characters of the dorsal ornamentation, in particular the numbers of lateral T-shaped tubercles on the median hood process and the elevation of the median paired spines of the scute. The proportions of the appendages and their microsculpture are also valuable, but the form of the penis is so similar in all species that it is of little systematic value. The absence of an inner tooth on the basal articles of the chelicerae of males may be synapomorphic for at least some Ortholasma; however, Ortholasma rugosa has a small tooth, and the tooth is not present in all species of Trilasma.
As delimited here, Ortholasma is found almost exclusively in the United States and Canada, though Ortholasma coronadense extends into Baja California, México, on Islas Coronados, offshore and just south of the border with the United States. This species has not been reported from mainland Baja California, but likely occurs there.
.
| 1. | Large animals, males 4.5 mm or longer, females 6 mm or longer; southern Sierra Nevada in Tulare and Kern Cos., California | Ortholasma colossus sp. n. |
| – | Smaller animals, males less than 4 mm, females 4.5 mm or less | 2 |
| 2. | Leg femora banded, body with median light stripe; northern California north to Vancouver Island | Ortholasma pictipes Banks |
| – | Leg femora not banded, body uniform in color | 3 |
| 3. | Leg femora without false articulations; San Francisco Bay area | Ortholasma rugosum Banks |
| – | At least second femora with false articulations | 4 |
| 4. | All femora with false articulations; San Francisco Bay area south to Los Angeles, Sierra Nevada foothills, caves in higher Sierras | Ortholasma levipes Shear & Gruber |
| – | False articulations only in femora 2 and 4; southern California and Coronado Islands | Ortholasma coronadense Cockerell |
urn:lsid:zoobank.org:act:78E03B21-192C-47C3-B964-C179BC06EF60
Figs 1– 8, 10, 20, 30, 31Male holotype and two female paratypes collected in Bear Den Cave, Sequoia National Park, Tulare Co., California, 1 May 2004, by J. Krejcaet al. deposited in California Academy of Sciences (CAS); male and female paratypes from same locality, collected 17–18 July 2003 by J. Krejca et al. deposited in Field Museum of Natural History (FMNH).

Ortholasma colossus sp. n., live female specimen in habitat. Photo courtesy of Marshal Hedin.
The notably larger size of the females distinguishes this species of Ortholasma from all others.
The species name, a Latin noun in apposition, refers to a gigantic statue, or by implication, anything outlandishly large for its type.
Male holotype: total length, 4.5, width, 2.25. Color uniform blackish brown, legs somewhat lighter brown. Carapace arcuate, about twice as wide as long, with complete lateral and posterior submarginal keels; median keels connect eye tubercle and innermost lateral hood process, lateral keels also arising on innermost lateral hood process. Two acute lateral hood processes each about half as long as median hood process. Circumocular keels absent, only subocular portion present. Median hood process arising dorsally on eye tubercle, length 1.6, width 1.0; bearing 24 lateral T-shaped tubercles, all connected. Metapeltidium free, complete keel along anterior margin. Scute 2.6 long, 2.25 wide. Pattern of scute keels typical, paired median scute spines low, scarcely standing above level of keels; posterior marginal keel with complete fenestrations (Fig. 2).
Chelicerae (Figs 4, 5, 10) with basal article 0.9 long, 0.25 wide, prominent epigamic gland with dense vestiture of small, fine setae (Fig. 5), otherwise sparsely setose; second article 0.75 long, 0.21 wide, with small, curved mediobasal tooth. Palpus (Figs 6, 20) with dense vestiture of clavate setae; trochanter with several ventral seta-tipped tubercles; tibia swollen; dimensions given in Table 1. Legs in order of length, 2 (11.95), 4 (9.7), 3 (6.3), 1 (5.8); metatarsus 2 with 8 false articulations, otherwise false articulations lacking; tarsi 1–4 with 10, 11, 10, 11 articles respectively. Lengths of leg segments given in Table 1. Leg femora with typical ornamentation (Fig. 7).
Genital operculum broadly rounded, marginate, suture faintly indicated. Penis (Fig. 30) 1.0 long, typically thin; glans (Fig. 31) sinuate with hooked tip, subtended by small setae.
Female paratype: total length, 6.6, width, 4.2. Nonsexual characters as in male (Fig. 3); median hood process 1.7 long, 0.9 wide. Scute 4.5 long, 4.2 wide. Chelicera without gland on basal article, distal article lacking tooth, basal article 1.05 long, 0.26 wide; distal article 0.90 long, 0.24 wide. Palpal tibia not swollen, dimensions given in Table 2. Legs in order of length, 2 (10.6), 4 (7.34), 3 (5.7), 1 (4.87); metatarsus 2 with 6 false articulations; tarsi 1–4 with 7, 8, 9, 11 articles respectively. Lengths of leg segments given in Table 2. Genital operculum broadly rounded, marginate, without suture. Ovipositor typical.
Appendage article measurements (mm) of Ortholasma colossus male.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 1.10 | 0.76 | 0.70 | - | 0.52 |
| Leg 1 | 1.90 | 0.60 | 1.10 | 1.10 | 1.10 |
| Leg 2 | 4.00 | 0.75 | 3.50 | 2.50 | 1.20 |
| Leg 3 | 1.90 | 0.40 | 2.00 | 0.90 | 1.10 |
| Leg 4 | 3.20 | 1.00 | 3.00 | 1.30 | 1.20 |
All specimens deposited in CAS unless otherwise noted. CALIFORNIA: Fresno Co., 3 mi. south of Trimmer, 4 June 1967, under rhyolite in oak forest, T. Briggs, female (AMNH). Tulare Co., Johnsondale, Kern River, 4 July 1956, V. Roth, W. Gertsch, female (AMNH); Lost Soldier’s Cave, December 1977, A. Grubbs, female (AMNH); 21 July 2003, J. Krejcaet al., female, 23 July 2003, molt fragment, 9 November 2003, female; Lightening Cave, 30 June 1952, A. Lange, female (used for SEM; AMNH); Sequoia National Park, Ash Mtn., 26 April 1951, E. Schlinger, male (AMNH); Paradise Cave, 30 April 2004, J. Krejca et al., male, juveniles; Lange Cave, 6 May 2004, J. Krejca et al., male, 2 females, 6 May 2004, male, female, 7 May 2004, late instar juvenile; Carmoe Crevice, 5 July 2003, J. Krejcaet al., male; May’s Cave, 16 May 2004, J. Krejca et al., female, early juvenile; Hidden Cave, 15 November 2003, J. Krejca, V. Loftin, male; Highway 245, 14 mi. north of Woodlake, near Cottonwood Creek and Rattlesnake Creek confluence, 26 March 2009, M. Hedin et al., female (pictured alive and in the field in Fig. 1).
Appendage article measurements (mm) of Ortholasma colossus female.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 1.10 | 0.90 | 0.70 | - | 0.46 |
| Leg 1 | 1.50 | 0.52 | 1.20 | 1.00 | 1.10 |
| Leg 2 | 3.50 | 0.80 | 2.60 | 2.10 | 1.60 |
| Leg 3 | 1.80 | 0.52 | 1.52 | 0.90 | 0.96 |
| Leg 4 | 2.20 | 0.80 | 2.20 | 1.10 | 1.04 |
The preponderance of cave records is
probably misleading; the species shows no signs of troglomorphosis and
specimens from caves are nearly identical to the few surface-collected
specimens. The cool, moist environment of caves probably attracts
individuals (troglophily). Caves have been much more thoroughly
collected than surface habitats, and in this case most of the records
come from a biological survey of the caves of Sequoia and Kings Canyon
National Parks. The caves mentioned in the Distribution section are
located on a map in
The size of the species, at least 50% longer than the next largest, is striking when seen side-by-side with congeners; only Ortholasma rugosum approaches it with some 4.5-mm-long males. However, most males of Ortholasma colossus sp. n. are longer than 4.5 mm, and the largest females of Ortholasma rugosum are only 4.8 mm long, compared to the usual 6–7 mm in colossus sp. n.
Trilasma bolivari Goodnight & Goodnight, 1942, by monotypy. Of Ruaxphilos, Ruaxphilos petrunkevitchou Goodnight & Goodnight, 1945, by monotypy.
Ortholasmatines in which the median hood process arises dorsally, projects anteriorly in a shallow curve, and is parallel-sided or nearly so, with a dorsal row or rows of tubercles (spoon-shaped and lacking dorsal tubercles in Ortholasma); two or three lateral hood processes; males without cheliceral glands (cheliceral glands present in Ortholasma); male first cheliceromere with distal inner tooth, dorsal tooth or sharp tubercle present or absent, curved mediobasal tooth on second cheliceromere (not verified for species known only from females); palpal patella with only two seta-bearing tubercles (Ortholasma with several); large keel cells separated by transverse rows of much smaller keel cells (Ortholasma lacks rows of smaller cells; Dendrolasma has either large or small cells but not both).
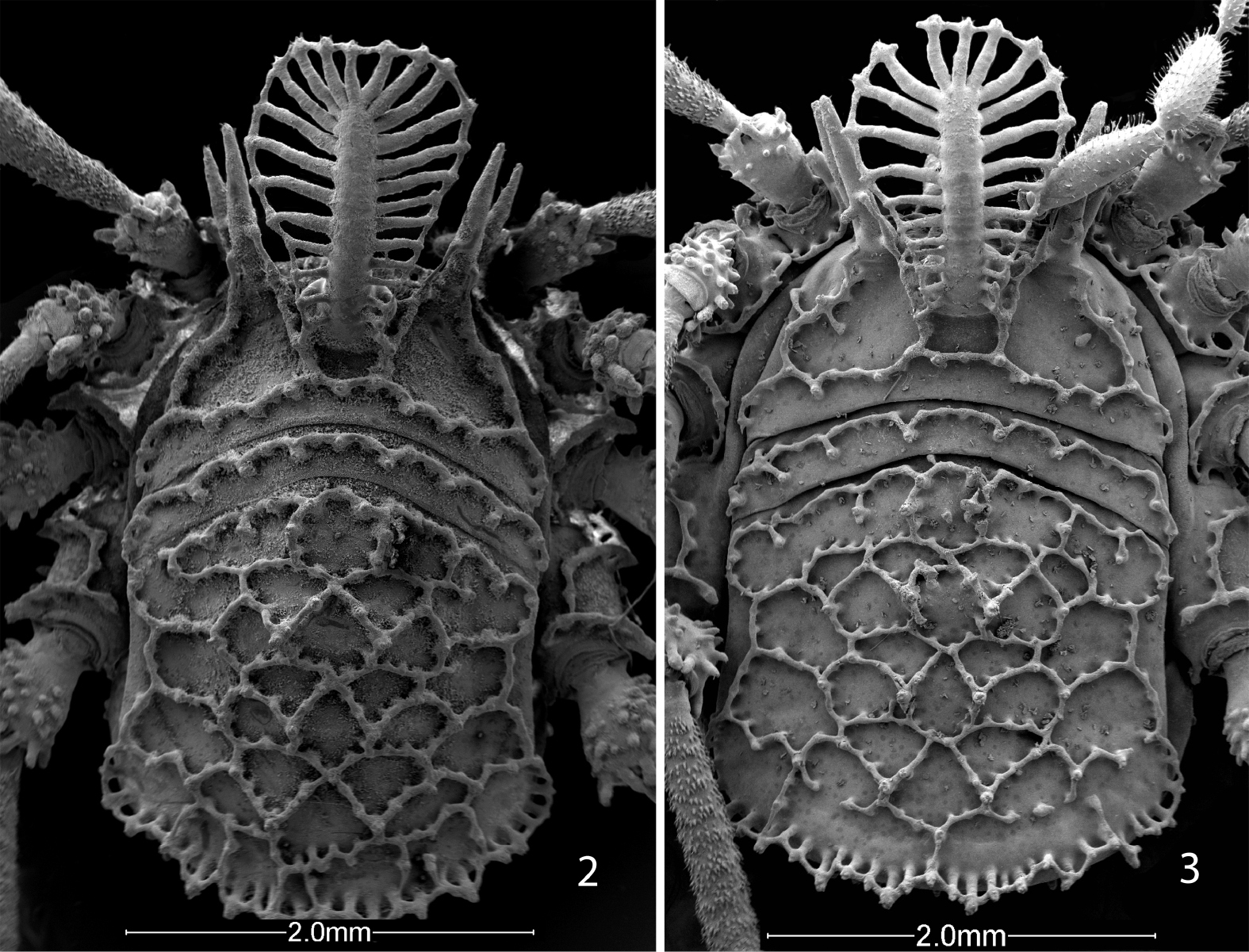
Scanning electron micrographs of Ortholasma colossus sp. n., dorsal views. 2 male 3 female.
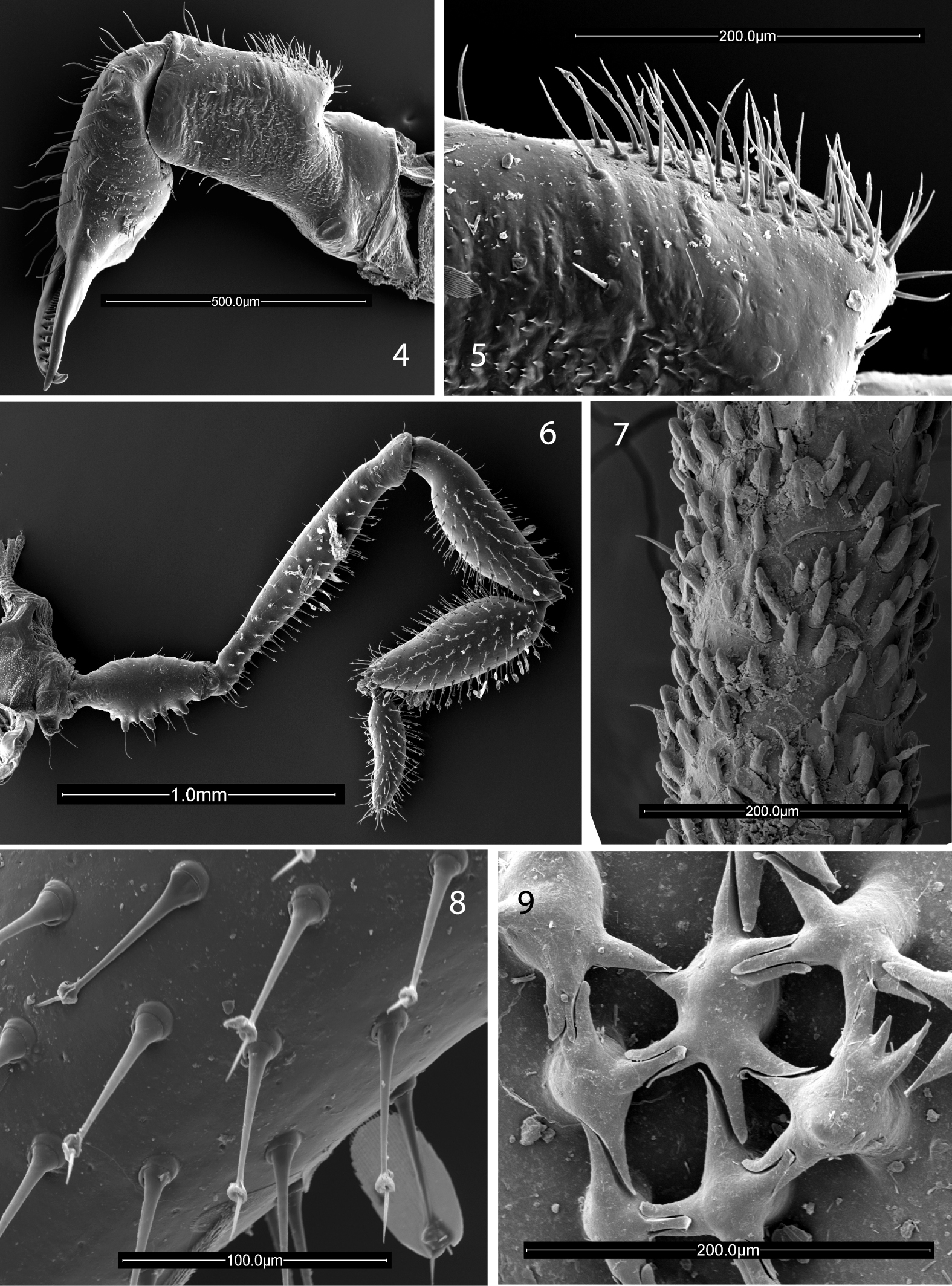
Scanning electron micrographs. 4–8 Ortholasma colossus sp. n. 4 right chelicerae of male, mesal view 5 glandular area of basal cheliceral article of male, mesal view 6 right palpus of male, mesal view 7 sculpture of second femur of female 8 kugelhaare of female palpus 9 Trilasma trispinosum sp. n., branched tubercles forming small keel cells of scute.
Trilasma consists of seven species extending from Nuevo León, México, in the north to Honduras in the south. Not much ecological information is available. One species, sbordonii, is troglomorphic, but while other cave collections exist, none of the other species seems especially adapted for life underground (although both petersprousei sp. n. and tempestado sp. n. have unusually long, thin legs and palpi) and probably are at most troglophilic. Surface collections are from cloud forest litter, and from under the bark of pines. Except for chipinquensis, collected at an altitude of about 1525 m (5000’), all species come from altitudes ranging from 2100–3050 m (7000–10000’) asl.
Trilasma species are immediately separable from Ortholasma species due to the presence of areas of small keel cells on the abdominal scutum (compare Figs 42, 45, and 54 with 1, 2). It appears that the small cell groups are at the anterior borders of the scute areas because in some species the first group is to be found at the very anterior margin of the scute (of area 1) and succeeding groups appear anterior to the paired median spines of the succeeding areas. The maximum number of groups is thus five, but the anteriormost is absent in some species, and the posteriormost is generally broken up into two groups on either side of the midline and consists of just two or three small cells on each side, or may be absent as well. The small cell groups of areas 2, 3 and 4 are the most marked and are successively wider, with the group of area 4 occupying about two-thirds the width of the scute and consisting of up to 20 small cells. In addition, Trilasma species have dorsal tubercles on the median hood process of the eye tubercle; these may be few (trispinosum sp. n., Fig. 43) or very numerous (tempestado sp. n., Fig. 44) and bearing branches that connect complexly with each other and with the usual lateral tubercles. Instead of the typically spatulate dorsal hood processes of Ortholasma species, Trilasma species have dorsal hood processes with nearly parallel sides that are obviously longer and narrower than in the former genus, reaching an extreme in the very narrow hood of Trilasma trispinosum sp. n. (Fig. 43). In several species the process tapers apically to a point.
Trilasma species are on average significantly smaller than Ortholasma species, ranging from a minimum of 2.1 mm in length to just 3.4 mm, including the hood. As in other members of the subfamily, the order of legs in length is 2, 4, 3, 1. In some species false articulations are present in at least some femora and metatarsi, but sometimes only in males. The detailed ornamentation of the leg femora, of taxonomic value in Ortholasma, because it differs between species, is quite uniform in Trilasma, consisting of numerous, short, club-shaped setae, among which are dispersed a few blunt, rod-shaped setae on elevated sockets. This character was studied for all species using scanning electron microscopy. The short, club-shaped setae are clearly seen to be hollow in some of the photomicrographs, but there appear not to be any pores communicating with the outside. In species of both Ortholasma and Dendrolasma, the longer setae on elevated sockets are acute and tapered, not blunt and rod-shaped.
Sexual dimorphism in Trilasma species is more pronounced than in Ortholasma species. Males are significantly smaller than females, with relatively longer legs, especially legs 2 and 4, which may also have increased numbers of tarsal segments, and in some species the males have false articulations in the fourth leg femora that are not present in the females. In addition to the basal, median tooth on the second cheliceral segment and the glands in the patellae and tibiae of the palpi, males of Trilasma species may have somewhat reduced dorsal ornamentation when compared to females, expressed in fewer small cells and lower paired tubercles of the dorsal areas. This dimorphism is especially notable at the posterior margin of the scute, where in females there is an obvious “post and rail fence” with long “posts” (Shear and Gruber, 1983); in males of the same species, this feature may be hardly noticeable.
The following characters seem to be useful in defining and separating the species of Trilasma: the relative length and robustness of the legs, the presence or absence and the number (if present) of false articulations in the femora and metatarsi of the legs (this may be sexually dimorphic in some species), the range of numbers of tarsal articles (difficult to evaluate because of small sample size; the counts given should be taken as variable by one or two articles in either direction), the relative prominence of the ocular keels, the patterns of small keel cells on the scute, the height of the paired area tubercles, and the shape and size of the dorsal hood process. In addition, there are autapomorphies of single species, such as the very narrow dorsal hood process and three lateral hood processes in Trilasma trispinosum sp. n.
While I am still certain that Ruaxphilos is a synonym of Trilasma, it is no longer clear that Ruaxphilos petrunkevitchou Goodnight & Goodnight, 1945 is a synonym of Trilasma bolivari, as placed by
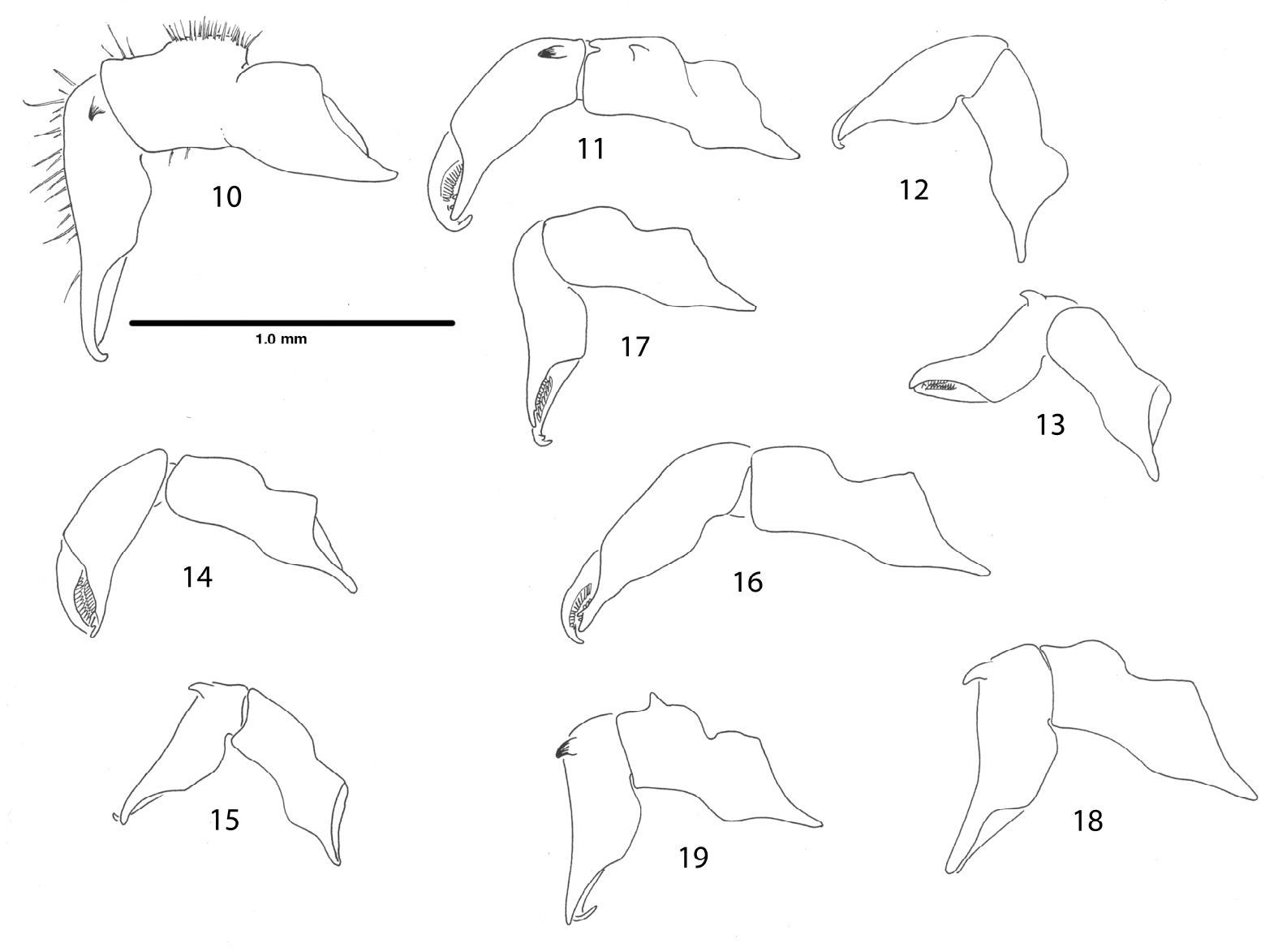
Right chelicerae, mesal views. 10 Ortholasma colossus sp. n., male 11 Trilasma ranchonuevo sp. n., male 12 Trilasma trispinosum sp. n., female 13 Trilasma trispinosum sp. n., male 14 Trilasma tempestado sp. n., female 15 Trilasma tempestado sp. n., male 16 Trilasma petersprousei sp. n., female 17 Trilasma chipinquensis sp. n., female 18 Trilasma hidalgo sp. n., male 19 Trilasma tropicum sp. n., male. All drawings to the same scale.
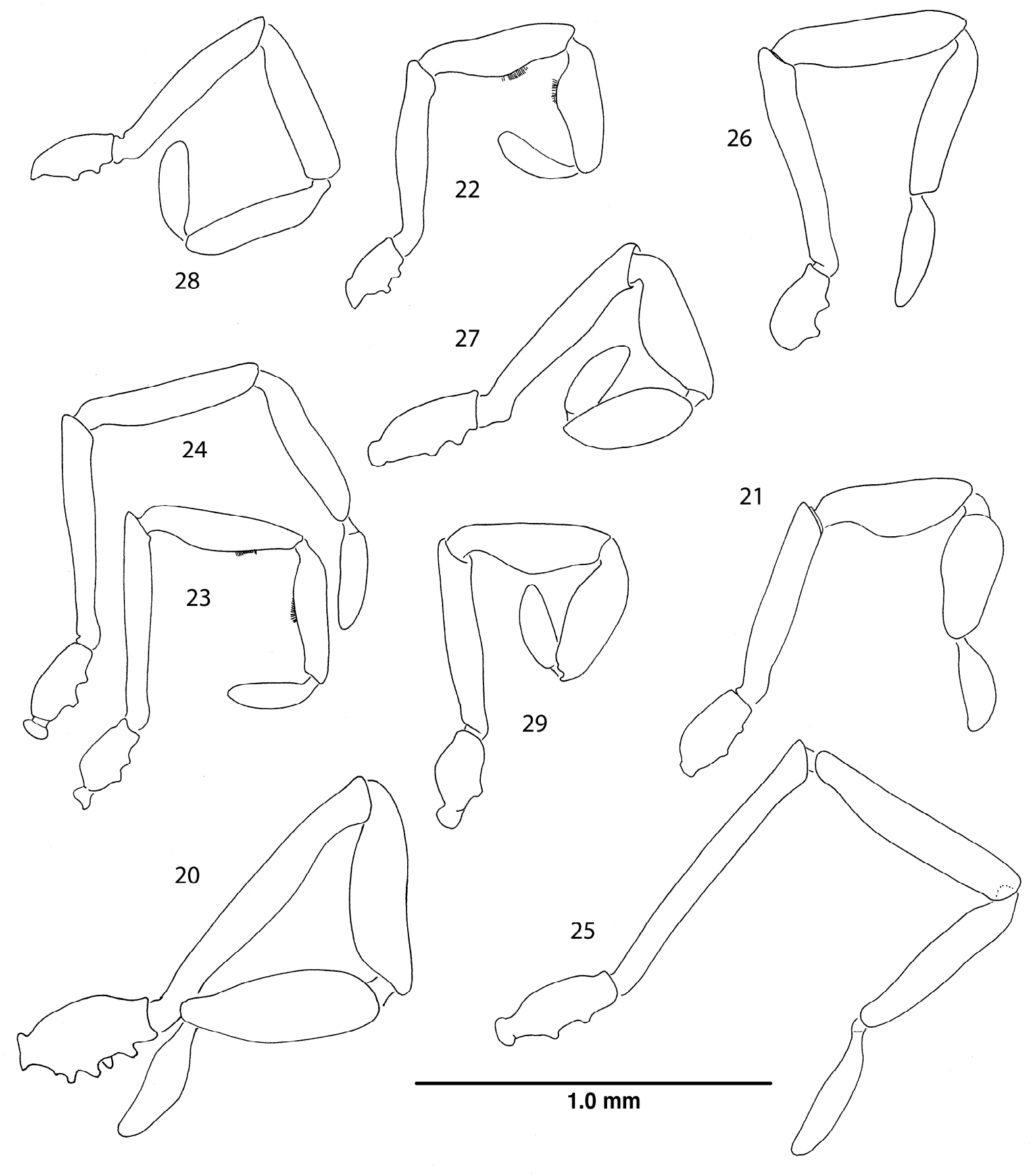
Right palpi, lateral views. 20 Ortholasma colossus sp. n., male 21 Trilasma ranchonuevo sp. n., male 22 Trilasma trispinosum sp. n., male 23 Trilasma tempestado sp. n., male 24 Trilasma tempestado sp. n., female 25 Trilasma petersprousei sp. n., female 26 Trilasma chipinquensis sp. n., female 27 Trilasma hidalgo sp. n., male 28 Trilasma tropicum sp. n., female 29 Trilasma tropicum sp. n., male. All drawings to the same scale.
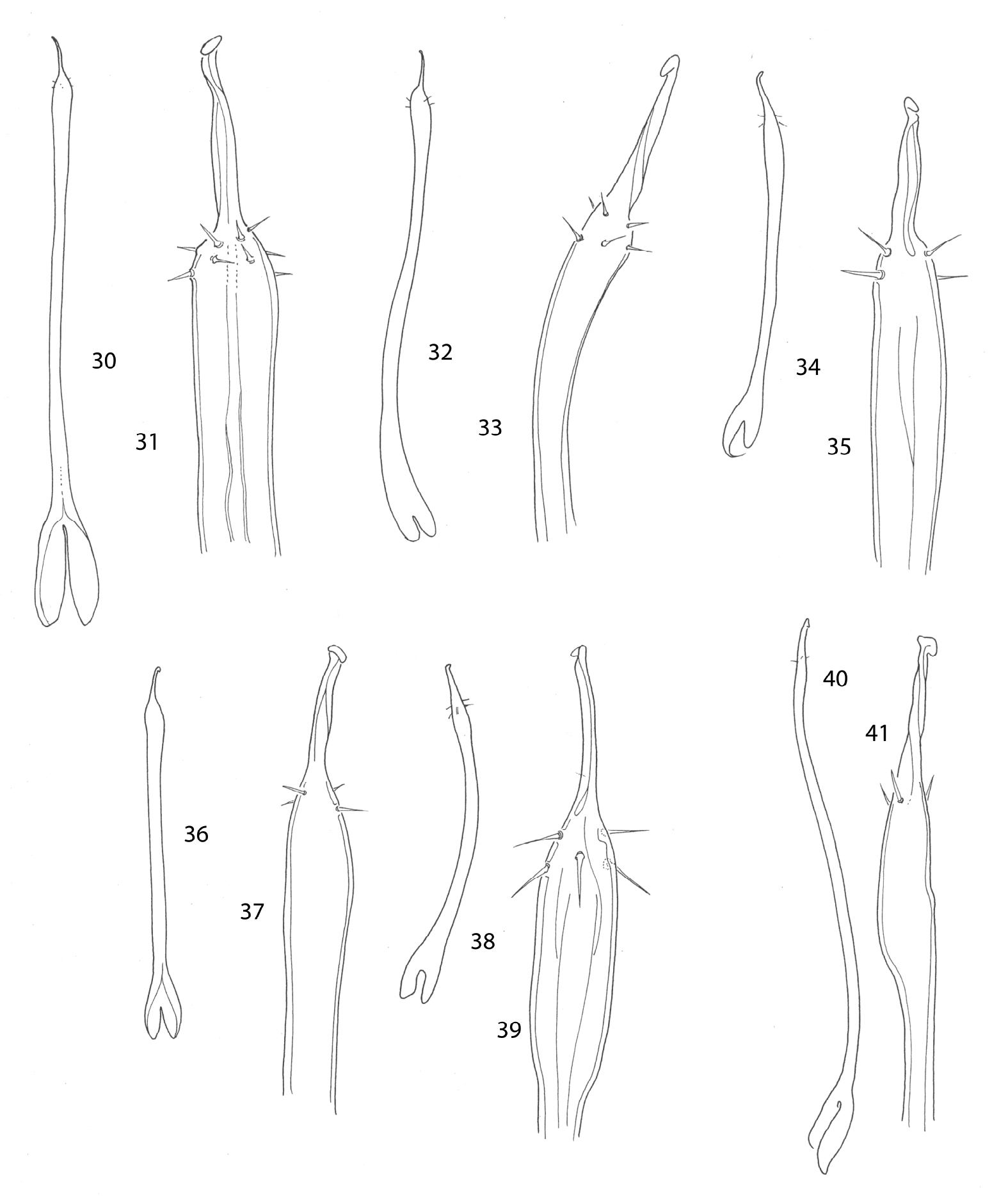
Penes. 30, 31 Ortholasma colossus 30 dorsal view 31 glans 32, 33 Trilasma ranchonuevo sp. n. 32 dorsal view 33 glans 34, 35 Trilasma trispinosum sp. n.34 dorsal view 35 glans 36, 37 Trilasma tempestado sp. n. 36 dorsal view 37 glans 38, 39 Trilasma tropicum sp. n. 38 dorsal view 39 glans 40, 41 Trilasma hidalgo sp. n.40 lateral view 41 glans. All drawings to the same scale; penis of Ortholasma colossus is 1 mm long.
.
| 1. | Troglomorphic facies, depigmented animals with reduced eyes and elongate legs and palpi; caves in Tamaulipas | Trilasma sbordonii (Šilhavý) |
| – | Medium tan or dark brown to black animals with normal eyes | 2 |
| 2. | Two lateral hood processes | 4 |
| – | Three lateral hood processes | 3 |
| 3. | Femora 2 and 4 of males each with a false articulations (females unknown); median hood process normally broad, with 5 or 6 dorsal tubercles; Tamaulipas | Trilasma ranchonuevo sp. n. |
| – | All femora of males with false articulations; median hood process extremely narrow, with only 1 or 2 dorsal tubercles; Veracruz | Trilasma trispinosum sp. n. |
| 4. | Femur of second leg longer than or equal to scute length | 5 |
| – | Femur of second leg shorter than scute | 6 |
| 5. | Fourth femur with two false articulations; second femur about as long as scute; Nuevo Léon | Trilasma tempestado sp. n. |
| – | All femora without false articulations; second femur longer than scute; San Luis Potosí | Trilasma petersprousei sp. n. |
| 6. | Fourth femur with a basal false articulation; Nuevo Léon | Trilasma chipinquensis sp. n. |
| – | All femora lacking false articulations | 7 |
| 7. | Second metatarsus with two or three false articulations; Distrito Federal, Puebla and adjacent states | Trilasma bolivari Goodnight & Goodnight |
| – | Second metatarsus without false articulations | 8 |
| 8. | Males about 3.0 mm long; about 15 dorsal tubercles on median hood process; Hidalgo, México | Trilasma hidalgo sp. n. |
| – | Males about 2.5 mm long; about 8 dorsal tubercles on median hood process; Honduras | Trilasma tropicum sp. n. |
urn:lsid:zoobank.org:act:7BCAB326-0BF9-414A-8E16-BC5773077567
Figs 11, 21, 32, 33, 47Male holotype (TMM) from Rancho Nuevo, Tamaulipas, México, collected 10 April 1982 by Terri Treacy.
Like Trilasma trispinosum sp. n., ranchonuevo sp. n. has three lateral hood processes, but the median hood process of ranchonuevo sp. n. is much broader and has more dorsal tubercles. There are more small keel cells on the scute of trispinosum sp. n. than in ranchonuevo sp. n.The male of ranchonuevo sp. n. has a single false articulation in each of the second and fourth leg femora; such are present in all legs of trispinosum sp. n. males. Tarsal counts of the single available specimen are 3, 4, 4, 4, the lowest for any Trilasma.
The species epithet, a noun in apposition, refers to the type locality.
Male holotype: total length, 2.7, width, 1.6. Color pale tan to yellowish brown. Carapace arcuate, about 1.5X as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connecting eye tubercle and innermost lateral hood process, lateral keels arising both on innermost and middle lateral hood processes. Three blunt lateral hood processes each about one-half as long as median hood process. Circumocular keels suppressed, subocular portion vaguely indicated, eyes relatively large, bulging. Median hood process arising dorsally on eye tubercle, with nearly parallel sides, then converging distally, length 0.9, width 0.3; median keels of carapace continuing as rows of lateral tubercles on median hood process, about 18–20 lateral tubercles, linearly connected; 5 or 6 dorsal tubercles present, connected linearly to one another but not obviously to lateral tubercles. Metapeltidium free, complete keel along anterior margin, 6 tubercles posterior to keel, connected to it by single branch each. Scute 1.6 long, 1.6 wide. All keels relatively low. Small keel cells present only on areas 2–4, as single transverse rows of 6–8 cells. Paired median scute spines prominent, on areas 4, 5 larger than adjacent keel tubercles (Fig. 47).
Chelicerae (Fig. 11) with basal article 0.62 long, 0.22 wide, sparsely setose, with small, median distal tooth; second article 0.62 long, 0.20 wide, with dark, median basal tooth. Palpus (Fig. 21) with dense vestiture of clavate setae; patella and tibia swollen, but glands not marked by patches of small setae, trochanter with ventral seta-tipped tubercles very low, almost obselete; dimensions given in Table 3. Legs in order of length, 2 (6.66), 4 (4.74), 3 (3.12), 1 (3.0); metatarsus 2 with 2 false articulations, femora 2, 4 with single basal false articulation; tarsi 1–4 with 3, 4, 4, 4 articles respectively. Lengths of leg segments given in Table 3. Length/width ratios of femora, in order: 3.64, 8.33, 3.46, 7.0. Leg femora with typical ornamentation.
Genital operculum broadly rounded, marginate, notched. Penis (Figs 32, 33) typical of genus.
Appendage article measurements of Trilasma ranchonuevo sp. n. male.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.70 | 0.50 | 0.50 | - | 0.31 |
| Leg 1 | 0.80 | 0.40 | 0.60 | 0.80 | 0.40 |
| Leg 2 | 1.50 | 0.60 | 1.20 | 1.94 | 1.60 |
| Leg 3 | 0.90 | 0.44 | 0.66 | 0.72 | 0.40 |
| Leg 4 | 1.40 | 0.52 | 1.00 | 1.00 | 0.82 |
The male holotype of Trilasma ranchonuevo sp. n. is significantly larger than the male paratype of Trilasma trispinosum sp. n. and has longer legs. The pattern of small cells on the scute is quite different, and the median hood process is much broader in relation to its width, with many more dorsal tubercles. On the left side of the body of the holotype, the innermost lateral hood process is irregularly developed, suggesting that the innermost process is the “extra” one and that its presence or absence might be subject to variation.
Rancho Nuevo is in the Sierra Nevada Oriental in western Tamaulipas, near the Nuevo Léon border and about 22 miles northwest of Ciudad Victoria. Coordinates: 23°51'50.40N; 99°27'07.43W, elevation 8600’ (2650 m).While the region around Rancho Nuevo is famous for its caves, this specimen was collected on the surface and shows no signs of troglobiosis.
urn:lsid:zoobank.org:act:F39EF9AA-DD3B-48F2-8C8E-B0EC3E17BF6D
Figs 9, 12, 13, 22, 34, 35, 42, 43Female holotype, male and female paratypes (TMM) from Puerto del Aire, Veracruz, México, collected 6 January 1966 by J. Richter, in cloud forest oak litter.
Differing from all other Trilasma except ranchonuevo sp. n.in the three, rather than two, lateral hood processes of the carapace, but distinct from ranchonuevo sp. n. in the extremely narrow median hood process, with only a few dorsal tubercles.
The species epithet refers to the three lateral hood processes.
Female holotype: total length, 2.6, width, 1.7. Color uniform chestnut brown, legs somewhat lighter brown, proximal parts of femora whitish yellow. Carapace arcuate, about 1.5X as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connect eye tubercle and innermost lateral hood process, lateral keels also arising on innermost lateral hood process. Three blunt lateral hood processes each about one-fourth as long as median hood process. Circumocular keels absent, subocular portion vaguely indicated, eyes relatively large, bulging. Median hood process arising dorsally on eye tubercle, narrow, length 0.9, width 0.11; median keels of carapace continue as rows of reduced lateral tubercles on median hood process, about 18–20 lateral tubercles, linearly connected; only 2 or 3 dorsal tubercles present. Metapeltidium free, complete keel along anterior margin, 6 tubercles posterior to keel, connected to it by single branch each. Scute 1.5 long, 1.7 wide. All keels relatively low. Small keel cells of scute area 1 absent, small keel cell rows progressively wider posteriorly, widest on area 4; small cells of area 5 in two groups either side of midline. Paired median scute spines small, on areas 4, 5 no larger than adjacent keel tubercles (Figs 42, 43).
Chelicerae (Fig. 12) with basal article 0.65 long, 0.18 wide, sparsely setose; second article 0.52 long, 0.18 wide. Palpus with dense vestiture of clavate setae; trochanter with ventral seta-tipped tubercles; dimensions given in Table 4. Legs in order of length, 2 (5.45), 4 (4.99), 3 (3.10), 1 (3.05); metatarsi without false articulations, femur 4 with single basal false articulation; tarsi 1–4 with 4, 5, 4, 5 articles respectively. Lengths of leg segments given in Table 4. Length/width ratios of femora, in order: 3.5, 7.0, 3.75, 5.25. Leg femora with typical ornamentation.
Genital operculum broadly rounded, marginate, with suture. Ovipositor typical of subfamily.
Male paratype: total length, 2.3, width, 1.2. Nonsexual characters as in female, but dorsal ornament reduced, obscured in paratype by secretion; median hood process 0.79 long, 0.2 wide. Scute 1.2 long, 1.2 wide. Chelicera (Fig. 13) without gland on basal article, second article with forward-projecting tooth, basal article 0.60 long, 0.20 wide; distal article 0.51 long, 0.15 wide. Palpal patella and tibia swollen, small ventral glandular areas indicated by patches of fine setae (Fig. 22), dimensions of palpus given in Table 5. Legs in order of length, 2 (5.76), 4 (4.96), 3 (3.32), 1 (3.22); all femora with single, basal false articulation, metatarsi without false articulations; tarsi 1–4 with 4, 5, 4, 5 articles respectively. Lengths of leg segments given in Table 5. Length/width ratios of femora, in order: 3.9, 9.8, 4.0, 6.1. Genital operculum broadly rounded, marginate, with two small, lateral notches. Penis typical (Figs 34, 35).
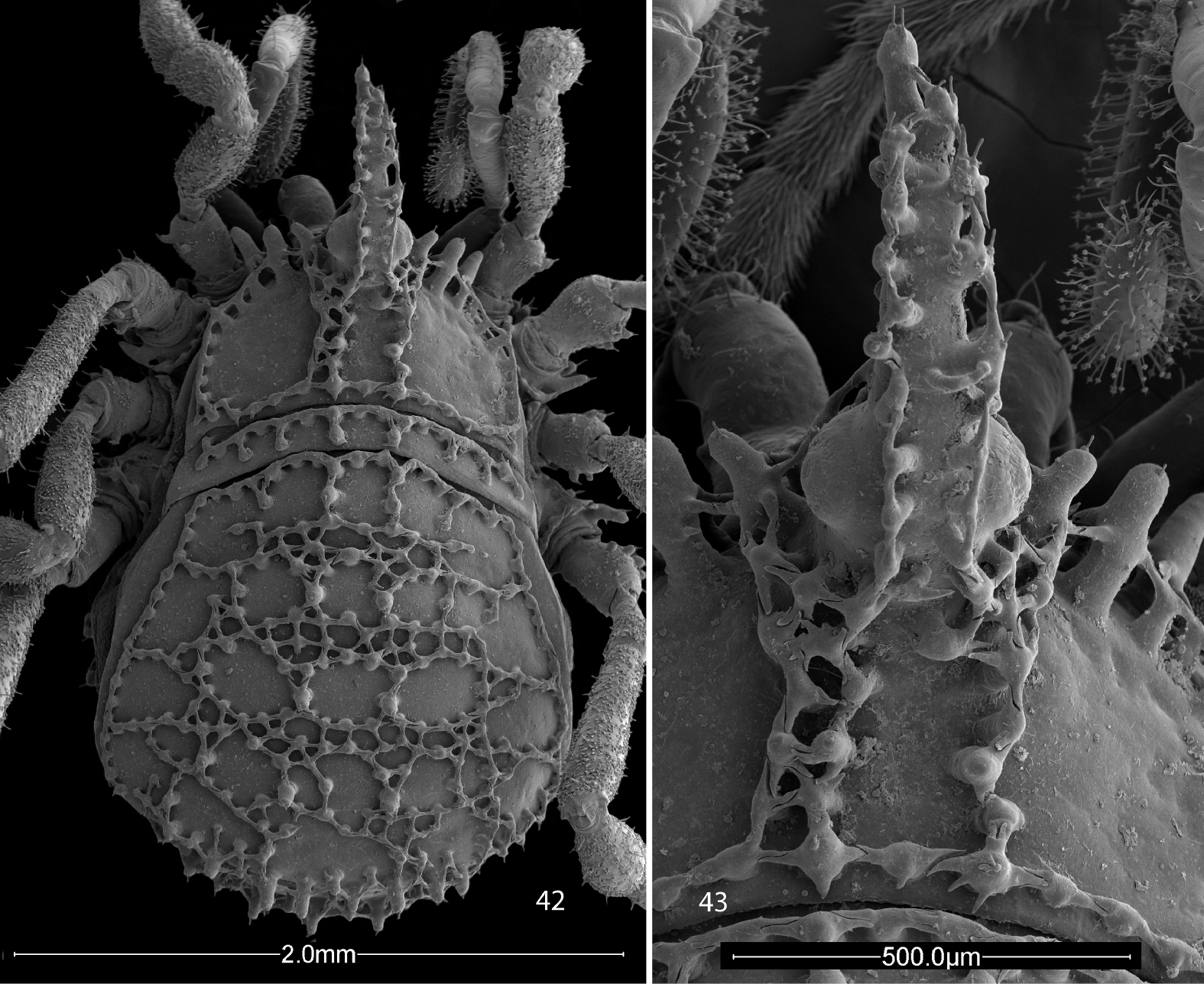
Scanning electron micrographs of female Trilasma trispinosum sp. n. 42 body, dorsal view 43 hood, dorsal view.
Appendage article measurements of Trilasma trispinosum sp. n. female.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.60 | 0.57 | 0.50 | - | 0.25 |
| Leg 1 | 0.70 | 0.40 | 0.50 | 0.80 | 0.65 |
| Leg 2 | 1.05 | 0.55 | 0.70 | 1.60 | 1.55 |
| Leg 3 | 0.75 | 0.40 | 0.50 | 0.80 | 0.65 |
| Leg 4 | 1.05 | 0.40 | 0.74 | 1.00 | 1.80 |
Appendage article measurements of Trilasma trispinosum sp. n. male.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.65 | 0.51 | 0.49 | - | 0.30 |
| Leg 1 | 0.78 | 0.40 | 0.50 | 0.80 | 0.74 |
| Leg 2 | 1.18 | 0.44 | 0.74 | 1.70 | 1.70 |
| Leg 3 | 0.80 | 0.38 | 0.60 | 0.74 | 0.80 |
| Leg 4 | 1.10 | 0.46 | 0.80 | 1.30 | 1.30 |
This species is the most distinctive of all Trilasma species due to the very short and narrow median hood process, which has only one or two dorsal armed tubercles. The female holotype (Figs 42, 43) is anomalous in that the keel extending from the eye tubercle to the posterior margin of the carapace is doubled on the left side. The pattern of keel cells is very distinct in the female holotype and paratype, but the male paratype has a much-reduced ornament. However, this is uncertain because of the thick secretion covering the dorsum and median hood process in the single available specimen; this material could not be cleaned off using ultrasonics, or methanol as a solvent. Notable also is the presence in the male of basal false articulations in all the femora. In the female, only the fourth femur has a basal false articulation. The presence of palpal glands in the male palpal patellae and tibiae is indicated by the patch of fine setae on the ventral side, and in still-adhering secretion on the tibia.
Puerto del Aire is a small village west of the larger city of Acultzingo, Veracruz, and southeast of Morelos Canadá, Puebla, virtually at the Puebla-Veracruz border (18°42'13.5N; 97°21'29.6W). The elevation is about 2556 m (7500 ft.) asl.
urn:lsid:zoobank.org:act:2755568D-A57D-4B19-80BF-62EE1547B1DC
Figs 14, 15, 23, 24, 36, 37, 44, 45, 48Holotype female and male paratype (TMM) from Cueva de Polvo Tempestado, 1–2 km south of San Josecito, Nuevo Léon, México, collected 1 March 1989 by George Veni and Allan Cobb; female paratype (TMM) from Sótano de las Tres Ventanas, Purifacición area, Cuauhtémoc, Nuevo Léon, México, collected 29 November 1981 by Paul Fambro; male paratype (TMM) from Pozo de las Pantaletas, Santa Marta de Arriba, 20 km southeast of Zaragoza (UTM 433010/2656459), collected 26 November 1999 by Peter Sprouse. See Notes, below, for a detailed discussion of these localities.
A long-legged, pale species most similar to Trilasma petersprousei sp. n., which is about half again as large. Trilasma tempestado sp. n.has the second femur about the same length as the scute, and has two false articulations in the fourth femora of both males and females, while petersprousei sp. n. lacks femoral false articulations. With the male paratype only 2.1 mm in length, this species is the smallest known Trilasma, about the same size as Martensolasma jocheni, the smallest known ortholasmatine.

Scanning electron micrographs of female Trilasma tempestado sp. n. 44 hood, dorsal view 45 body, dorsal view.
The species epithet is a noun in apposition, referring to the type locality.
Female holotype: total length, 2.6, width, 1.5. Color pale tan to yellowish brown. Carapace arcuate, about 1.5X as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connecting eye tubercle and innermost lateral hood process, lateral keels arising both on innermost and middle lateral hood processes. Two blunt lateral hood processes each about one-third or less as long as median hood process. Circumocular keels suppressed, but subocular portion visible, eyes relatively small. Median hood process arising dorsally on eye tubercle, with nearly parallel sides, then converging distally, widest at about midlength, length 0.90, width 0.40; median keels of carapace continuing as rows of lateral tubercles on median hood process, about 20 lateral tubercles, linearly connected; 11–12 dorsal tubercles present, connected complexly to one another and to lateral tubercles. Metapeltidium free, complete keel along anterior margin, 4 tubercles posterior to keel, connected to it by single branch each. Scute 1.6 long, 1.5 wide. All keels relatively high. Small keel cells present on areas 1–5; area 1 with 3–4 small cells in midline, area 2 with about 10 small cells in transverse row less than ½ width of scute, area 3 with about 16 small cells in midline, area 4 with 20–23 small cells in transverse row about 2/3 width of scute, area 5 with two paramedian groups of 3–4 small cells. Paired median scute spines relatively prominent, on all areas distinctly larger than adjacent keel tubercles (Figs 44, 45).
Chelicerae (Fig. 14) with basal article 0.60 long, 0.20 wide, sparsely setose; second article 0.56 long, 0.18 wide. Palpus (Fig. 24) slender, with dense vestiture of clavate setae; trochanter with two prominent seta-bearing ventral tubercles; dimensions given in Table 6. Legs in order of length, 2 (8.80), 4 (6.12), 3 (4.48), 1 (4.32); metatarsus 2 with 3 false articulations, femora 4 with 3 false articulations; tarsi 1–4 with 5, 6, 7, 8 articles respectively. Lengths of leg segments given in Table 6. Length/width ratios of femora, in order: 7.33, 16.67, 8.0, 12.14. Leg femora with typical ornamentation.
Genital operculum broadly rounded, separated from sternite by suture. Ovipositor typical of genus.
Male paratype: total length, 2.1, width, 1.3. Color uniform light chestnut brown. Nonsexual characters as in female (see Figs 44, 45, 48 and description, above), but dorsal ornament somewhat reduced; median hood process 0.70 long, 0.25 wide. Scute 1.3 long, 1.3 wide. Chelicera (Fig. 15) without gland on basal article, second article with strong, anteriodorsal, slightly hooked, conical protuberance, basal article 0.50 long, 0.17 wide; distal article 0.52 long, 0.16 wide. Palpal patellae and tibiae swollen, epigamic glands present and marked by patches of fine setae (Fig. 23), dimensions of palpus given in Table 7. Legs in order of length, 2 (8.68), 4 (6.00), 3 (4.48), 1 (4.26); metatarsus 2 with 2 false articulations, femur 4 with 3 false articulations; tarsi 1–4 with 5, 5, 6, 6 articles respectively. Lengths of leg segments given in Table 7. Length/width ratios of femora, in order: 7.43, 20.0, 6.89, 10.5. Genital operculum broadly rounded, marginate, with two small, lateral notches. Penis typical (Figs 36, 37).
Appendage article measurements of Trilasma tempestado sp. n. female.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.80 | 0.65 | 0.56 | - | 0.34 |
| Leg 1 | 1.10 | 0.40 | 0.70 | 1.12 | 1.00 |
| Leg 2 | 2.00 | 0.60 | 1.50 | 2.90 | 1.80 |
| Leg 3 | 1.12 | 0.40 | 0.80 | 1.16 | 1.00 |
| Leg 4 | 1.70 | 0.40 | 1.28 | 1.50 | 1.24 |
Appendage article measurements of Trilasma tempestado sp. n. male.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.72 | 0.60 | 0.46 | - | 0.30 |
| Leg 1 | 0.90 | 0.46 | 0.64 | 0.90 | 0.90 |
| Leg 2 | 1.50 | 0.56 | 1.08 | 2.00 | 1.30 |
| Leg 3 | 0.84 | 0.50 | 0.66 | 0.80 | 0.96 |
| Leg 4 | 1.40 | 0.56 | 1.28 | 1.00 | 1.10 |
Like Trilasma petersprousei sp. n., this species is pale and long-legged, yet does not appear to be a troglobiont. It occurs in at least two distinct karst areas. Cueva de Polvo Tempestado is located 1–2 km south of the village of San Josecito (23°58'12.68N; 99°54'21.18W, elev. ca. 2300m (7570’)). This is a small, vertical cave, one of several in the area. Sótano de las Tres Ventanas is a pit in the well-known Río Purificación karst region located in Nuevo Léon and Tamaulipas between Zaragoza, NL, and Ciudad Victoria, Tamps. The coordinates are 23°53’30.583N; 99°28’20.445W, at an elevation of 2210 m asl. The entrance pit drops 35 m to a rubble pile, which slopes down to a plug at -45 meters. Temperature measured at this point was 8.6° C during exploration on 29 November 1981 (P. Sprouse, pers. comm.) Sistema Purificación, located in this area, is one of the deepest and longest cave systems in México. Pozo de las Pantaletas is located near the village of Santa Marta at coordinates of 23°51’40.16N; 99°41’23.982W and an elevation of 2800 m asl. This is a vertical cave with multiple rope drops leading to a depth of 140 m. All of these records are rather tightly clustered in a mountainous region of Nuevo Léon that makes an easterly salient into Tamaulipas 35–45 km northwest of Ciudad Victoria. It would be no surprise to see specimens from the adjoining part of Tamaulipas.
urn:lsid:zoobank.org:act:A8DA8437-E2B4-4F4E-8434-05EEAFAC56BC
Figs 16, 25, 46Female holotype (TMM) from Hoya de las Guaguas, 10 km south of Aquismón, San Luis Potosí, México, collected 29 August 1986 by Peter Sprouse.
Trilasma petersprousei sp. n. is a long-legged species, with long, thin pedipalps. The high tarsal count of the holotype (6, 12, 6, 8) and the presence of 12 false articulations in the second metatarsi also distinguish it from other species.
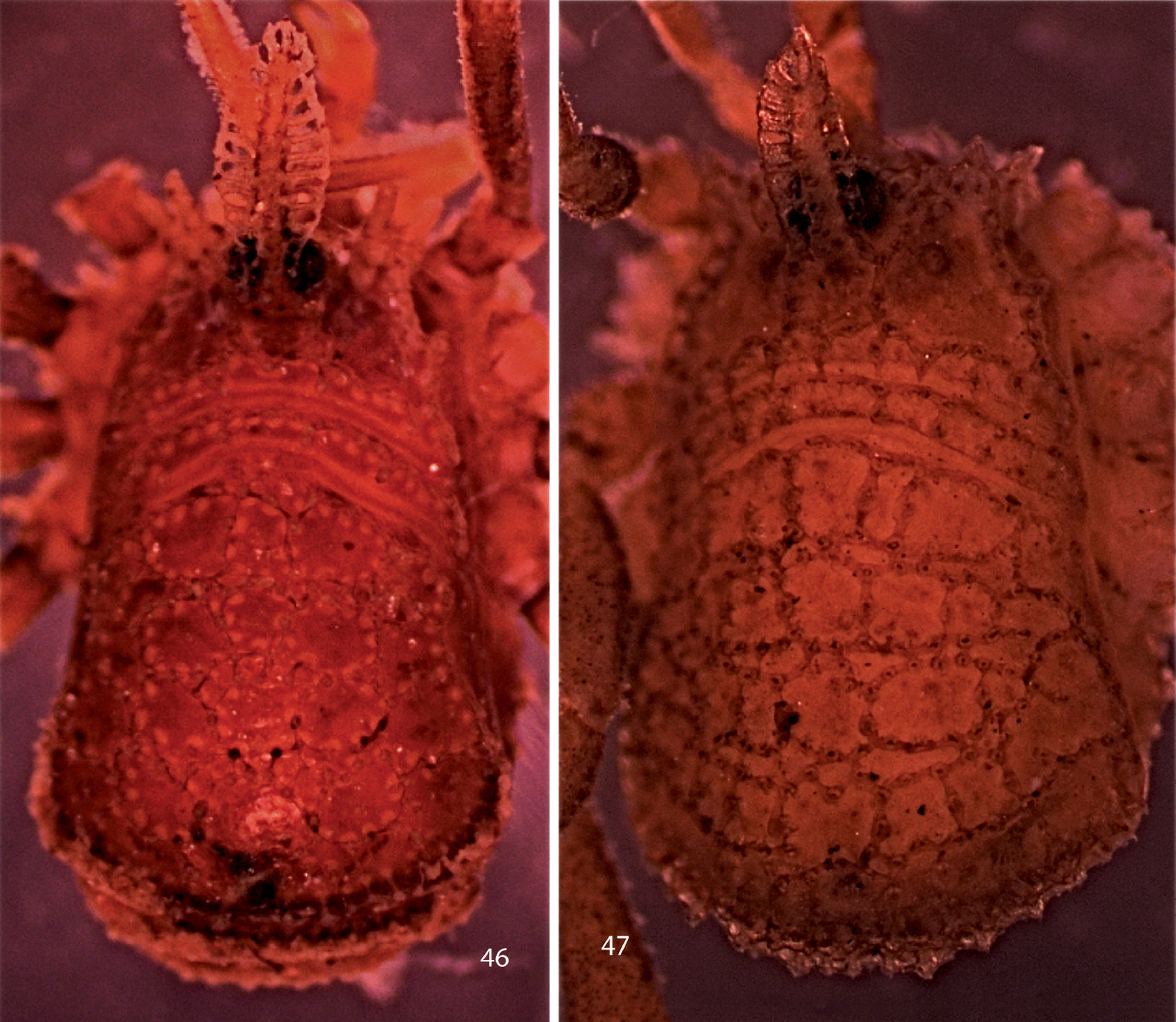
Photographs of bodies, dorsal views. 46 male Trilasma petersprousei sp. n.47female Trilasma ranchoneuvo sp. n.
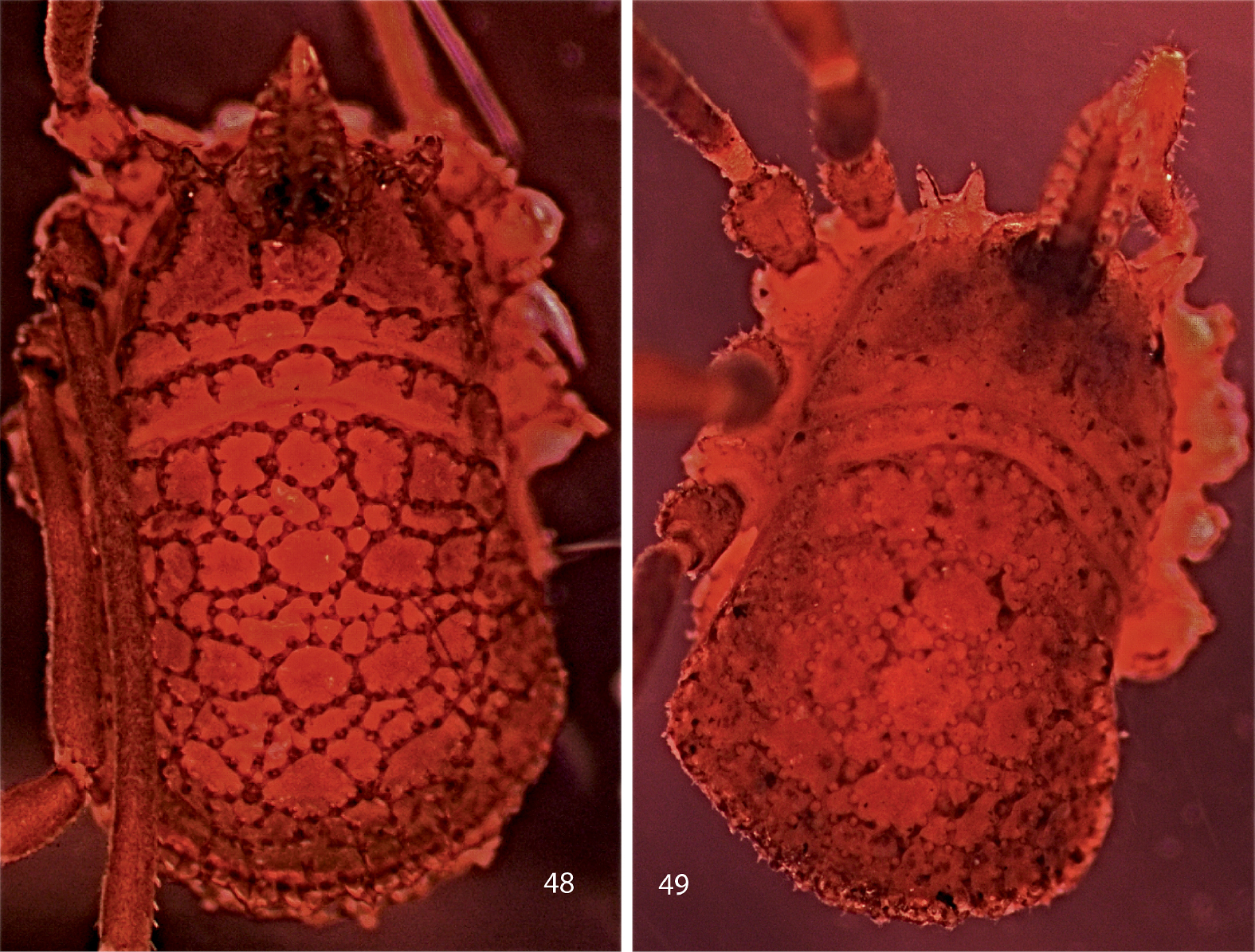
Photographs of bodies, dorsal views. 48 male Trilasma tempestado sp. n. 49 male Trilasma tropicum sp. n.
The species is named for the collector, Peter Sprouse of Zara Environmental, LLC, a noted explorer of Mexican caves who has collected many new species of troglobionts and troglophiles.
Female holotype: total length, 3.4, width, 1.9. Color pale tan to yellowish brown. Carapace arcuate, about 1.5X as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connecting eye tubercle and innermost lateral hood process, lateral keels arising both on innermost and middle lateral hood processes. Two blunt lateral hood processes each about one-half as long as median hood process. Circumocular keels suppressed, but subocular portion obvious, eyes relatively large, bulging. Median hood process arising dorsally on eye tubercle, with nearly parallel sides, then converging distally, widest slightly beyond midlength, length 1.10, width 0.45; median keels of carapace continuing as rows of lateral tubercles on median hood process, about 20 lateral tubercles, linearly connected; about 15 dorsal tubercles present, connected complexly to one another and to lateral tubercles. Metapeltidium free, complete keel along anterior margin, 6 tubercles posterior to keel, connected to it by single branch each. Scute 1.8 long, 1.9 wide. All keels relatively low. Small keel cells present on areas 1–4; area 1 with 2–3 small cells in midline, area 2 with 4–6 small cells in transverse row less than ½ width of scute, area 3 with 5–6 small cells in midline, area 4 with 10–12 small cells in transverse row about ½ width of scute. Paired median scute spines not prominent, on areas 4, 5 only slightly larger than adjacent keel tubercles (Fig. 46).
Chelicerae (Fig. 16) with basal article 0.70 long, 0.23 wide, sparsely setose; second article 0.72 long, 0.20 wide. Palpus (Fig. 25) slender, with dense vestiture of clavate setae; trochanter with two prominent seta-bearing ventral tubercles; dimensions given in Table 8. Legs in order of length, 2 (14.05), 4 (9.43), 3 (5.90), 1 (5.73); metatarsus 2 with about 12 false articulations, femora without false articulations; tarsi 1–4 with 6, 12, 6(7), 9 articles respectively. Lengths of leg segments given in Table 8. Length/width ratios of femora, in order: 8.25, 36.8, 9.0, 28.2. Leg femora with typical ornamentation.
Genital operculum broadly rounded, separated from sternite by suture. Ovipositor typical of genus.
Appendage article measurements of Trilasma petersprousei sp. n. female.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 1.00 | 0.86 | 0.70 | --- | 0.44 |
| Leg 1 | 1.65 | 0.60 | 1.08 | 1.15 | 1.25 |
| Leg 2 | 4.05 | 0.90 | 3.10 | 3.50 | 2.50 |
| Leg 3 | 1.80 | 0.50 | 1.30 | 1.05 | 1.25 |
| Leg 4 | 3.10 | 1.10 | 2.40 | 1.25 | 1.58 |
While Trilasma petersprousei sp. n. has long, thin legs and palpi, it does not appear in any other way to be a troglobiont; the eyes are large and well-pigmented and it lacks the exaggerated median hood process of Trilasma sbordonii, a true troglobiont.
Hoya de las Guaguas (sometimes spelled Huahuas in Spanish) is an immense pit, 478 m (1565’) deep, developed in limestone of the Sierra Huasteca, and located at 21°31'56'N; 99°02'01.15W, about 460 m (1500’) asl. The entrance is large enough that sufficient light reaches the floor to sustain a plant community (P. Sprouse, pers. comm.).
urn:lsid:zoobank.org:act:6C2FDB74-23B9-4266-92F9-FE96DD129CB6
Figs 17, 26, 50Female holotype (AMNH) from Chipinque Mesa, Monterrey, Nuevo Léon, México, collected 24 June 1969 by Stewart B. Peck.
The paired median area tubercles are strongly developed in this species and project well above the level of the keels. Like trispinosum sp. n., there is a single false articulation in female femur 4, but trispinosum sp. n. has 3 lateral hood processes, while chipinquensis sp. n. has 2.

Photographs of bodies, dorsal views. 50 female Trilasma chipinquensis sp. n. 51 male Trilasma hidalgo sp. n.
The species epithet, an adjective, refers to the type locality.
Female holotype: total length, 2.7, width, 1.5. Color dark brown, nearly black (possibly artifact of preservation). Carapace arcuate, about 1.5X as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connecting eye tubercle and innermost lateral hood process, lateral keels arising both on innermost and middle lateral hood processes. Two blunt lateral hood processes each about one-half as long as median hood process. Circumocular keels strongly developed, but subocular portion easily seen in dorsal view, eyes relatively small. Median hood process arising dorsally on eye tubercle, with nearly parallel sides, only slightly converging distally, length 0.95, width 0.40; median keels of carapace continuing as rows of lateral tubercles on median hood process, about 16 lateral tubercles, linearly connected; about 6 dorsal tubercles present, connected in a single row to one another but not to lateral tubercles. Metapeltidium free, complete keel along anterior margin, 6 tubercles posterior to keel, connected to it by single branch each. Scute 1.7 long, 1.5 wide. All keels well elevated above dorsum. Small keel cells present on areas 2–4; areas 2, 3 with 5–8 small cells in midline, area 4 with 10–12 small cells in transverse row about 2/3 width of scute. Paired median scute spines prominent, on all areas significantly larger than adjacent keel tubercles, standing well above keels; pair of spines also present on metapeltidium (Fig. 50).
Chelicerae (Fig. 17) with basal article 0.64 long, 0.21 wide, sparsely setose; distal article 0.60 long, 0.19 wide. Palpus (Fig. 26) relatively slender, with dense vestiture of clavate setae; trochanter with two prominent seta-bearing ventral tubercles; dimensions given in Table 9. Legs in order of length, 2? (-), 4 (5.62), 3 (4.66), 1 (4.20); metatarsi without false articulations, femora 4 without single false articulation; tarsi 1–4 with 5, -, 7, 6 articles respectively (tarsi 2 missing). Lengths of leg segments given in Table 9. Length/width ratios of femora, in order: 5.56, 12.86, 5.56, 7.60. Leg femora with typical ornamentation.
Genital operculum broadly rounded, separated from sternite by suture. Ovipositor typical of genus.
Appendage article measurements of Trilasma chipinquensis sp. n. female.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.80 | 0.32 | 0.30 | --- | 0.40 |
| Leg 1 | 1.00 | 0.40 | 0.64 | 1.20 | 0.96 |
| Leg 2 | 1.80 | 0.60 | 1.10 | --- | --- |
| Leg 3 | 1.00 | 0.46 | 1.00 | 1.20 | 1.00 |
| Leg 4 | 1.52 | 0.50 | 1.00 | 1.40 | 1.20 |
The holotype female was mentioned and briefly described by
Chipinque Mesa is a ridge of the Sierra Madre Oriental overlooking the city of Monterrey, to the north. Approximate coordinates are 25°36'29.43N; 100°21'18W; elevation at the top of the ridge is 1524 m (5000’). Chipinque Mesa is now a part of the Parc Nacional Cumbre and is a popular sight-seeing destination for visitors to Monterrey. It is densely forested in pines.
I no longer include Ruaxphilos petrunkevitchou Goodnight & Goodnight in the synonymy of Trilasma bolivari, as
I provide some new illustrations (Figs 52, 53) for comparison with the other species described here as new. The illustrations are of a female specimen collected by Fred Coyle at the pass between Toluca and México City, Distrito Federal, on Rt. 15, elev. 3000 m (9800’) asl, on 4 June 1982.
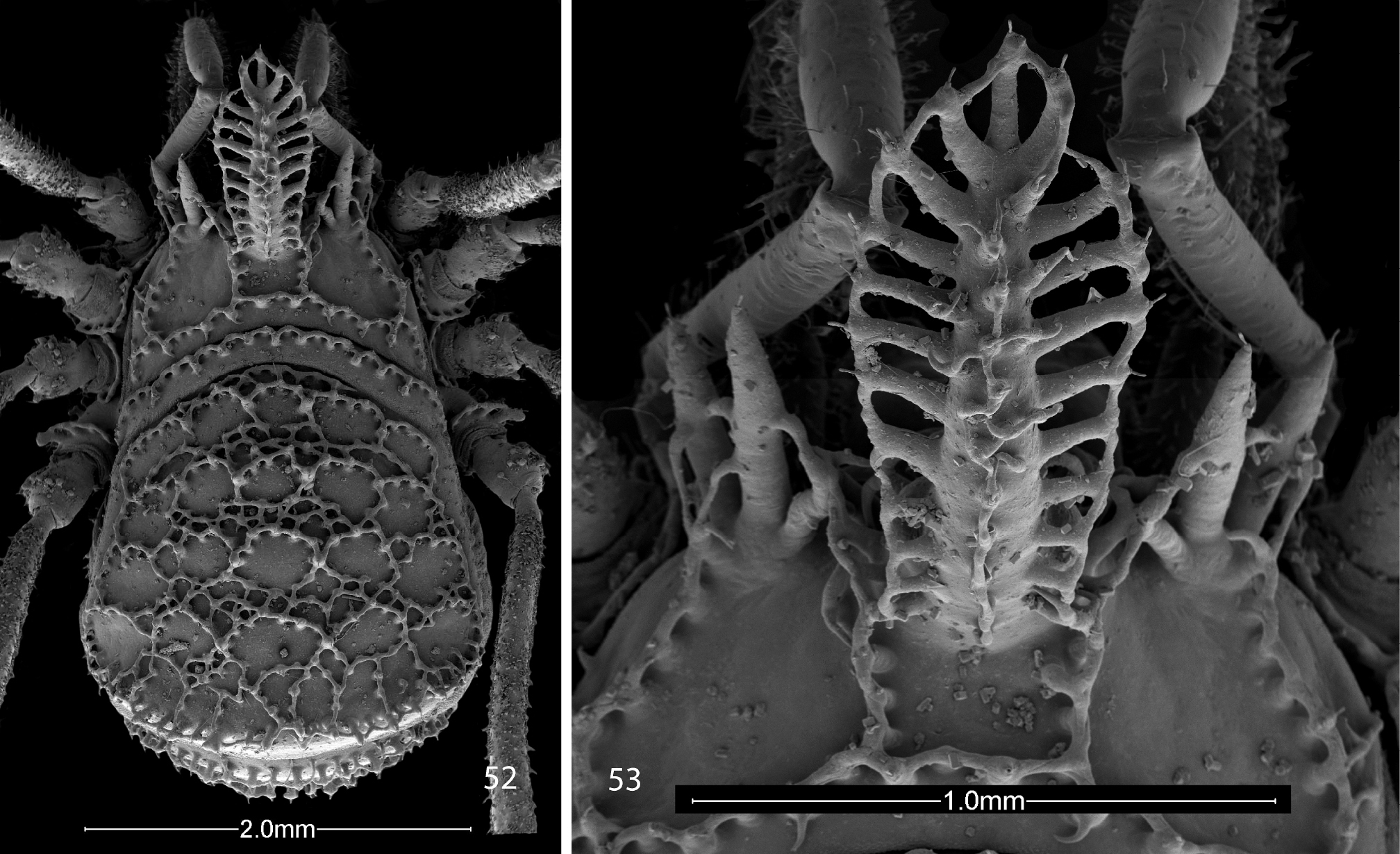
Scanning electron micrographs of female Trilasma bolivari
urn:lsid:zoobank.org:act:1CA2B4A7-FC11-471B-A1DF-BF4236BDB5B2
Figs 18, 27, 40, 41, 51Male holotype (TMM) from El Chico, Pachuco, Hidalgo, México, collected 1 January 1976, no collector named.
This species is closest to Trilasma bolivari sp. n., but differs from it in having fewer small cells on the scute, shorter, stouter legs, and less prominent scute area tubercles. In addition the dorsal tubercles on the median hood process of Trilasma bolivari sp. n. are much more numerous and are scattered over the stem of the process, with connections to the lateral tubercles; Trilasma hidalgo sp. n. has fewer dorsal tubercles which are arrayed in a line and only rarely connected to the lateral tubercles.
The species epithet is a noun in apposition, referring to the Mexican state of Hidalgo.
Male holotype: total length, 3.2, width, 1.3. Color dark brown, nearly black. Carapace arcuate, about 1.5X as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connecting eye tubercle and innermost lateral hood process, lateral keels arising both on innermost and middle lateral hood processes. Two blunt lateral hood processes each about one-third as long as median hood process. Circumocular keels strongly developed, subocular portion especially prominent. Median hood process arising dorsally on eye tubercle, widest point past midline of length, length 1.0, width 0.4; median keels of carapace continuing as rows of lateral tubercles on median hood process, about 30 lateral tubercles, linearly connected; about 15 dorsal tubercles present, connected in a single row to one another but not to lateral tubercles. Metapeltidium free, complete keel along anterior margin, 8 tubercles posterior to keel, connected to it by single branch each. Scute 1.3 long, 1.3 wide. All keels well elevated above dorsum. Small keel cells present on areas 2–5; area 2 with 5 or 6 small cells in single transverse row; area 3 with 10 to 12 small cells in single transverse row, but row is slightly wider than row on area 2; area 4 with two paramedian groups of 2 or 3 small cells each; area 5 similar, but only 1 or 2 small cells. Paired median scute spines prominent, significantly larger than adjacent keel tubercles on areas 2–5 (Fig. 51).
Chelicerae (Fig. 18) with basal article 0.82 long, 0.28 wide, sparsely setose; second article 0.65 long, 0.22 wide. Palpus (Fig. 27) stout, tibia, patella crassate, with dense vestiture of clavate setae; trochanter with two prominent seta-bearing ventral tubercles; dimensions given in Table 10. Legs in order of length, 2 (7.44), 4 (5.70), 3 (4.01), 1 (3.98); no false articulations; tarsi 1–4 with 4, 5, 5, 6 articles respectively. Lengths of leg segments given in Table 10. Length/width ratios of femora, in order: 4.58, 12.50. 4.58, 8.0. Leg femora with typical ornamentation.
Genital operculum broadly rounded, not separated from sternite by suture. Penis typical of genus (Figs 40, 41).
Appendage article measurements of Trilasma hidalgo sp. n. male.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.80 | 0.56 | 0.44 | --- | 0.37 |
| Leg 1 | 1.10 | 0.60 | 0.80 | 0.70 | 0.78 |
| Leg 2 | 2.00 | 0.70 | 1.70 | 1.60 | 1.44 |
| Leg 3 | 1.10 | 0.56 | 0.90 | 0.65 | 0.80 |
| Leg 4 | 1.60 | 0.60 | 1.60 | 0.90 | 1.00 |
“El Chico” doubtless refers to what is
now Parque Nacional El Chico, located north of the city of Pachuca. The
Parque is extensive but approximate coordinates are 20°12'26N; 98°43'52W;
elevations within the park range from 2300–3090 m (7540–10131’) asl.
The mountains are covered with a dense pine forest, with fir at the
higher elevations.
urn:lsid:zoobank.org:act:58A227AB-0922-4C73-A739-510A54F7732F
Figs 19, 28, 29, 38, 39, 49, 54, 55Male holotype and female paratype from “Las Ventas, Honduras, ” collected 11 February 1939 by R. V. Chamberlin (AMNH).
The short, relatively crassate legs and the extremely prominent subocular keels (Fig. 55) separate this species from others. Trilasma ranchonuevo sp. n. (Tamaulipas, México) has false articulations in femora 2 and 4 that are absent in Trilasma tropicum sp. n. The male has an acute dorsal knob on the basal article of the chelicerae, as well as the tooth on the second article.

Scanning electron micrographs of female Trilasma tropicum sp. n. 54 body, dorsal view 55 eye tubercle, dorsal view.
The species epithet refers to the occurrence of the species in the Neotropics.
Male holotype: total length, 2.5, width, 1.4. Color uniform light chestnut brown, metatarsi and tarsi of legs 3, 4 whitish yellow. Nonsexual characters as in female (see Figs 54, 55, and description, below), but dorsal ornament reduced, partially obscured in holotype by secretion; median hood process 0.80 long, 0.3 wide. Scute 1.4 long, 1.4 wide.
Chelicera (Fig. 19) without gland on basal article but with dorsal, conical protuberance, second article with forward-projecting tooth, basal article 0.50 long, 0.21 wide; distal article 0.55 long, 0.19 wide. Palpal patellae and tibiae swollen, epigamic glands probably present but not marked by patches of fine setae (Fig. 29), dimensions of palpus given in Table 11. Legs in order of length, 2 (6.44), 4 (5.34), 3 (3.76) , 1 (3.80); legs lacking false articulations; tarsi 1–4 with 5, 5, 6, 7 articles respectively. Lengths of leg segments given in Table 11. Length/width ratios of femora, in order: 4.5, 10.7, 4.2, 7.8.
Genital operculum broadly rounded, marginate, with two small, lateral notches. Penis typical (Figs 38, 39).
Female paratype: The female paratype (Figs 54, 55) is mounted on a SEM stub, therefore it was not possible to get accurate measurements of the appendages. Total length, 2.9, width, 1.8. Carapace arcuate, about twice as wide as long, with complete lateral and posterior submarginal keels; pair of median keels connect eye tubercle and innermost lateral hood process, lateral keels also arising on innermost lateral hood process. Two blunt lateral hood processes each about one-fourth as long as median hood process. Circumocular keels prominent, subocular portion strongly developed, eyes of usual size. Median hood process arising rostrally on eye tubercle, relatively narrow, length 0.8, width 0.3, ten fenestrations on each side, sides nearly parallel; median keels of carapace continue as rows of reduced lateral tubercles on median hood process, about 20–22 lateral tubercles, linearly connected; about 8 dorsal tubercles present, connected to each other but not to lateral tubercles. Metapeltidium free, complete keel along anterior margin, 10 tubercles posterior to keel, connected to it by single branch each. Scute 1.7 long, 1.7 wide. All keels relatively low. Small keel cells of scute area 1 present, small keel cell rows progressively wider posteriorly, widest on area 4; small cells of area 5 few, in two small groups near midline. Paired median scute spines moderately developed, on areas 4 distinctly larger than adjacent keel tubercles.
Chelicerae as described for male, but lacking dorsal protuberance on basal article, tooth on second article. Palpus (Fig. 28) as in male but patellae and tibiae not swollen. Legs in order of length, 2, 4, 3, 1; leg articles without false articulations. Leg femora with typical ornamentation.
Genital operculum broadly rounded, marginate, with suture.
Appendage article measurements of Trilasma tropicum sp. n. male.
| Femur | Patella | Tibia | Metatarsus | Tarsus | |
|---|---|---|---|---|---|
| Palpus | 0.72 | 0.60 | 0.46 | --- | 0.30 |
| Leg 1 | 0.90 | 0.46 | 0.64 | 0.90 | 0.90 |
| Leg 2 | 1.50 | 0.56 | 1.08 | 2.00 | 1.30 |
| Leg 3 | 0.84 | 0.50 | 0.66 | 0.80 | 0.96 |
| Leg 4 | 1.40 | 0.56 | 1.28 | 1.00 | 1.10 |
I mounted the only female on an SEM stub under the impression there was a second female in the collection, but the second specimen turned out to be a male. Thus measurements of the appendages of the female became impossible, but there is no reason not to suspect that the typical sexual dimorphism in appendage lengths is characteristic of this species.
Las Ventas is not listed for Honduras by the U. S. Board on Geographical Names (http://geonames.nga.mil/ggmagaz), nor could it be found by Google Earth. There are four places named Las Ventanas and one called Las Ventanillas, scattered in four different departments of Honduras. There is a village of Las Ventas in El Salvador (13°35'58.55N; 88°21'00W; 365 m (1050’) asl) but it is in the central part of the country, not close to the border with Honduras. Otherwise the name evidently is not used for a populated place in Central America. The type locality is therefore in doubt, but because the collection was made 70 years ago, it is possible “Las Ventas” was in existence then in Honduras, having since disappeared from geographic databases, or the label is a lapsus for one of the Las Ventanas. In any case, if this species is really from Honduras, it occurs farther south than any New World dyspnoan, at approximately the same latitude as Cladolasma angka in Thailand.
I am grateful to Jean Krejca, Stewart Peck, James Reddell, Lorenzo Prendini and Darrell Ubick for loans of material, and to Fred Coyle for specimens of Trilasma bolivari sp. n. from México. Marshal Hedin provided the photograph in Fig. 1, and additional specimens of Ortholasma colossus sp. n. I especially thank Peter Sprouse for detailed information on cave localities in México. Detailed comments from Peter Schwendinger, Jürgen Gruber, Adriano Kury and an anonymous referee materially improved the manuscript. Asa Kreevich assisted with laboratory preparations. Work on this project was financially supported by grants from the Professional Development Committee of Hampden-Sydney College.