(C) 2010 Nikita Vikhrev. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The taxonomic reasons for regarding Lispe draperi Séguy, 1933, sp. rev., as a valid species instead of a synonym of Lispe tentaculata (De Geer, 1776) and for treating Lispe quaerens Villeneuve, 1936, syn. n., as a junior synonym of Lispe sericipalpis Stein, 1904 are given. A revised key for the Palaearctic members of the Lispe tentaculata species-group is given. Data on ecology, distribution and feeding preferences are provided.
Lispe tentaculata, Lispe consanguinea, Lispe draperi, Lispe sericipalpis, Lispe orientalis, Lispe quaerens, Muscidae, Diptera, key, Palaearctica, new synonym
The Lispe tentaculata species-group was proposed by
Currently eight species are placed in the Lispe tentaculata species-group, six of which are present in the Palaearctic region. The taxonomic status of Lispe alpinicola Zhong, Wu & Fan, 1981 has not yet been settled (see below). Lispe sericipalpis Stein, 1904 and Lispe orientalis Wiedemann, 1824 are also distributed in the Oriental region. Lispe tentaculata (De Geer, 1776) is recorded from the very north of the Oriental region and widespread in Nearctic. Two more species of this group are found in the Nearctic region.
Material and methodsThe majority of the specimens studied are in the Zoological Museum of Moscow University (not indicated in text). Other material is in Natural History Museum, London (BMNH), Zoological Institute, St Petersburg (ZIN), Zoölogisch Museum, Universiteit van Amsterdam (ZMAN).
The following abbreviations for morphological structures are used: f1, t1, f2, t2, f3, t3 = fore-, mid-, hind- femur or tibia; ac = acrostichal setae; dc = dorsocentral setae; a, p, d, v = anterior, posterior, dorsal, ventral seta(e); prst – presutural, post - postsutural.
Here I suggest a new abbreviation for the tarsi as tar followed by a pair of digits separated by a hyphen: the first digit (1 to 3) gives the leg number and the second digit (1 to 5) the number of the tarsal segment. For example, tar1-4 = 4-th segment of fore tarsus; tar3-1 = hind basitarsus.
Identification key for Palaearctic species of the Lispe tentaculata species-group| 1 | Males | 2 |
| – | Females | 6 |
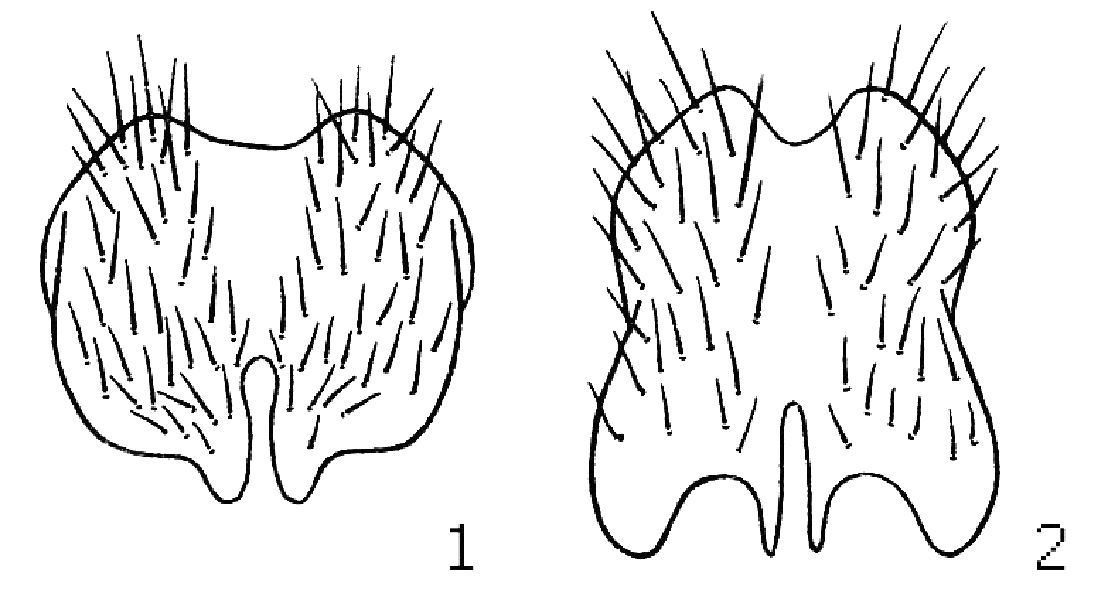
| 2 | Fore tarsus modified: tar1-1 yellow to dark, tar1-2 to tar1-4 yellow, tar1-5 black; tar1-1 half as long as tar1-2, with a dense row of brush-like av setulae and on posterior side with a finger-like yellow process with black apex, this process reaching middle of tar1-2; tar1-2 projecting ventrally. Male cercal plate as Fig. 1 | 3 |
| – | Fore tarsus unmodified. Male cercal plate as Fig. 3 | 5 |
| 3 | Median third of f3 with 2–5 av setae 1.5–2 times longer than femoral width. tar3-1 shortened (slightly more than one third as long as t3); ventral median part of tar3-1 concave; basal 1/3 of tar3-1 with a brash of ventral hairs contrasting with shorter hairs in apical 2/3. Scutellum with some fine hairs below at apex (these hairs may be invisible in old specimens). Cercal plate with 2 pairs of projections apically, distinctly longer than wide (Fig. 1.2). 3 strong post dc setae | 4 |
| – | Median 1/3 of f3 without long av setae (though in basal 1/3 of f3 with 3–4 av subequal to femoral width). tar3-1 straight, not curved or concave, tar3-1 longer (slightly less than half as long as t3); ventral hairs on tar3-1 of uniform length. Scutellum bare below at apex and bare at apex laterally below apical scutellar bristles. Cercal plate with only 1 pair of projections apically, almost as wide as long (Fig. 1.1). Usually 4 post dc setae, anterior 2 post pairs short and fine (but sometimes 3 strong post dc as in tentaculata!). Palearctic, temperate zone | Lispe consanguinea Loew |
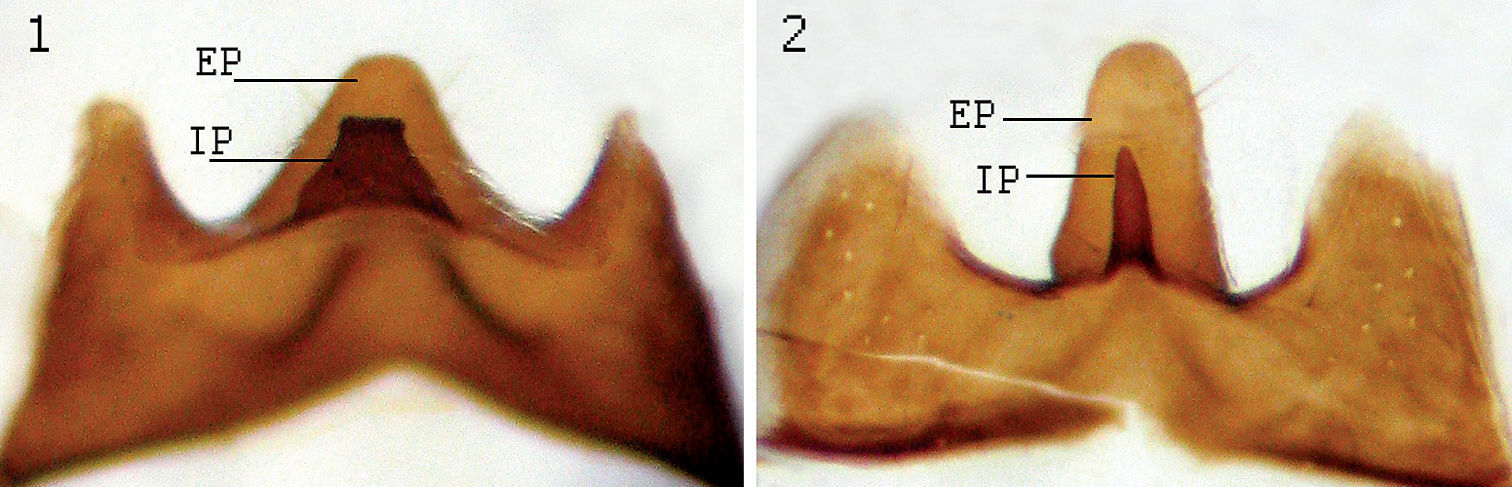
| 4 | Male sternite 5 as in Fig. 2.1. Tibiae dark, yellow only at basal fifth. Median 1/3 of f3 with 3–5 av setae. Basal 1/3 of f3 with 3–4 av setae subequal to femoral width. Holarctic | Lispe tentaculata (De Geer) |
| – | Male sternite 5 as in Fig. 2.2. At least t2 yellow in ground colour, more or less grey dusted, usually both t2 and t3 entirely yellow. Median 1/3 of f3 with 2–3 av setae. Basal 1/3 of f3 with 0–2 av setae subequal to femoral width. North Africa | Lispe draperi Séguy |
| 5 | Palpi black. f2 with 2–7 fine pv setae in basal 1/3. f3 with a long v-pv seta at base, 1–4 av setae in apical 2/5 and at most with a sparse row of 7–8 pv. tar1-2 and tar1-3 ventrally dark like the rest of fore tarsus. Presutural ac in 4–5 irregular rows. Dusting greyish. Length 5–6mm. Male terminalia – Figs 3.3, 4. Southern Palearctic and Oriental | Lispe sericipalpis Stein |
| – | Palpi yellow to brownish. f2 with a full and dense row of 20–30 pv setae and full row of about 20 av (in both rows, setae long in basal half and short in apical half). f3 with full rows of about 15 av and pv setae, the longest setae beyond middle twice as long as femoral width (long v-pv seta at base of f3 also present, but not as conspicuous among other setae). tar1-2 and tar1-3 yellow ventrally. Presutural ac in 6–7 irregular rows. Dusting brownish. Length 6.5–7.5mm. Male terminalia – Figs 3.1, 2. Southern Palearctic and Oriental | Lispe orientalis Wiedemann |
| 6 | Only posterior pair of prst dc present. Presutural ac hairs weak and short, arranged in 5–7 irregular rows | 7 |
| – | Both pairs of prst dc present. Presutural ac hairs stronger, arranged in 3–4 rows | 8 |
| 7 | Palpi black (blackish brown in specimens collected 50–100 years ago). Presutural ac hairs in 5 rows. f3 without av setae in basal 3/5. Length 5–6mm | Lispe sericipalpis Stein |
| – | Palpi yellow to light brown. Presutural ac hairs in 6–7 rows. f3 with a row of 7–9 av in basal 3/5. Length 6–7mm | Lispe orientalis Wiedemann |
| 8 | Scutum with a median pruinose patch between 2-nd and 3-rd post dc setae; 4 strong post dc setae, 2nd and 3rd post dc closely approximated. (Scutellum with some fine hairs below at apex) | 9 |
| – | Scutum without median pruinose patch between 2-nd and 3-rd post dc setae; 3 strong post dc setae or only 2 strong post dc (and 1–2 weak anterior post dc), closely approximated strong post dc absent | 10 |
| 9 | Tibiae dark, only knees yellow. f3 usually with 2–3 long submedian av; basal 1/3 of f3 with 1–4 av subequal to f3 width | Lispe tentaculata (De Geer) |
| – | t2 at least in basal half yellow in ground colour, more or less dusted, usually both t2 and t3 entirely yellow. f3 usually with only 1 long submedian av; basal 1/3 of f3 with 0–3 av hardly longer than half f3 width | Lispe draperi Séguy |
| 10 | Scutellum bare below at apex. t2 and t3 yellow, more or less dusted. 2+4 dc, but the anterior 2 post pairs short and fine. f3 in median 1/3 without av 1.5–2 times longer than femoral width (but 1–3 av setae subequal to f3 width present at basal 1/3 of f3) | Lispe consanguinea Loew |
| – | Scutellum with some fine hairs below at apex (these hairs often invisible on old specimens). 2+3 dc, all strong. f3 in median 1/3 with 1–3 av 1.5–2 times longer than f3 width (females with male chaetotaxy of scutum) | Lispe tentaculata (De Geer) |
Male cercal plates. 1 Lispe consanguinea Loew 2 Lispe tentaculata (De Geer) (from Hennig, 1960).
Male sternite 5, view from inner side. 1 Lispe tentaculata (De Geer) 2 Lispe draperi Séguy; EP external median process IP internal median process.
Male terminalia. 1, 2 Lispe sericipalpis Stein, 1 cercal plate 2 sternite 5; 3, 4 Lispe orientalis Wiedemann, 3 cercal plate 4 sternite 5 (from two points of view). <br/> In Lispe sericipalpis the cercal plate is more pointed, sternite 5 has the median process thinner and longer.
Described from Lhasa (China, 3500–4000 m asl.). Male fore and hind basitarsus modified as in Lispe tentaculata, female scutum with a pruinose patch as in Lispe tentaculata (Xue & Dong, 2005), male cercal plate similar to Lispe tentaculata (Xue & Chao, 1998). According to
Over 130 specimens. Moldova; Russia (European): Arkhangelsk reg., Chuvashia, Krasnodar reg., Moscow reg., Tver reg., Nizhnyi Novgorod reg., Vologda reg., Ulyanovsk reg.; Mongolia; Russia (Asian): Amur reg., Khanty-Mansi reg., Krasnoyarsk reg., Kurgan reg., Novosibirsk reg., Primorsky Kray; Tajikistan: Dushanbe; Turkey: Sakarya prov., Zonguldak prov.
Morocco: east of Marrakech, 1400m, stones on river bank, 22.III.2009, N.Vikhrev, 1♀; west of Marrakech, Oued Nfiss, stones on river bank, 23.III.2009, N.Vikhrev, 2♂♂, 5♀♀; near Essaouira, stones on river bank and pond/pool silt, 24–29.III.2009, N.Vikhrev, 11♂♂, 4♀♀.
The conspecifity of the material listed above with type of Lispe draperi Séguy was kindly confirmed by A.C. Pont (pers. comm.) who examined the holotype of Lispe draperi in the Muséum national d’Histoire naturelle, Paris.
Azerbaijan: Lenkoran env., 38.66°N, 48.79°E, 22–25.X.2008, N.Vikhrev, 16♂♂, 6♀♀.
India: Rajastan, Jaipur, 21.II. 2011, N.Vikhrev, 1♂, 1♀.
Russia: Krasnodarsky Kray, Sochi env., 43.547°N, 39.811°E, 29.IX-24.X.2010, D.Gavryushin, 4♂♂; Primorsky Kray, 42.86°N, 133.62°E, 18.IX.1987, A.Ozerov, 1♂.
Tajikistan: Khatlon div., Farkhor (=Parkhar) env., 37.420°N, 69.352°E, 07.VI.2010, K.Tomkovich, 7♂♂, 5♀♀; Khatlon div., Kulob, 37.909°N, 69.784°E, 07.VI.2010, K.Tomkovich, 19♂♂, 22♀♀.
Thailand: Mae Hong Son prov., 19.57N 98.28E, 650m asl., 21.XI.2010, N.Vikhrev, 1♂.
Turkey: Izmir prov., Dilek Milli Park, 37.68°N, 27.10°E, 20.XII.2006, N.Vikhrev, 1♀; Antalia prov., Silion ruins, 36.989°N, 30.985°E, goat drinking bowl, 25.V.2008, N.Vikhrev, 1♂, 1♀; Hatay prov., Arzus env., 36.407°N, 35.886°E, 14.IV.2010, N.Vikhrev, 8♂♂, 2♀♀; Hatay prov., Samandag env., Çivlek, 17.IV.2010, N.Vikhrev, 6♂♂.
S. Korea: Seoul env., 31.VII.1938, Zhenzhurist, 5♀♀.
Lectotype male of sericipalpis – male (Fig. 4), paralectotypes 1♂, 4♀♀ (ZMAN).
Azerbaijan: Lenkoran reg., 38.65°N, 48.80°E, 25.V.2009, K.Tomkovich, 1♀.
Myanmar: Shan state, Inle Lake env., 20.664°N, 96.966°E, 26–30.XI.2009, N.Vikhrev, 4♂♂, 2♀♀; Kakaw env., 20.64°N, 96.59°E, 03.XII.2009, N.Vikhrev, 45♂♂, 3♀♀.
Nepal: Solukhumbu distr., Janbesi env., 27.581°N, 86.548°E, 2660m asl., 19.III.2010, A.Reshchikov, 2♂♂, 2♀♀.
Tajikistan: Dushanbe division, Ramit env. (38.72N 69.32E), river bank, 15–16.VI.2010, K.Tomkovich, 17♂♂, 30♀♀; Dushanbe env., 13.V.1943, A.Stackelberg, 1♂ (with handwritten label by W.Hennig “Lispe quaerens”) (ZIN); Varzob Canyon, 28–29.VII.1939, L.Zimin, 4♀♀ (ZIN); Varzob Canyon, 04.VII.1937, A.Gussakovsky, 1♂ (ZIN).
Turkey: Antalia prov., Köprü River, 37.075°N, 31.232°E, 06–10.IX.2009, N.Vikhrev, 40♂♂, 29♀♀ (3♂♂, 2♀♀ deposited in BMNH); Mersin prov., 37.194°N, 34.810°E, forest stream, 23.IV.2010, N.Vikhrev, 1♀; Bolu prov., 40.498°N, 31.890°E, forest stream, 1800m asl., 31.VIII.2009, N.Vikhrev, 1♂; Sarakya prov., Karasu env., 41.03°N, 30.79°E, forest stream, 15.VI.2010, N.Vikhrev, 1♀, 28.VIII.2009, N.Vikhrev, 1♂, 1♀; Zonguldak prov., Alaply env., 41.14°N, 31.36°E, forest stream, 21.VI.2010, N.Vikhrev, 3♂♂, 2♀♀.
Lispe sericipalpis Stein, male lectotype (designated by Pont, 1970). Photo by Joke van Erkelens.
Male. Ground colour black. Pollinosity grey, but may be yellowish-grey. Palpus black(ish), but becoming brown in old specimens. Fronto-orbital plate and parafacial whitish, rarely yellowish. Scutum with 3 brown vittae along ac and dc rows, submedian (dc) vittae sometimes almost indistinct. dc 2(1) + 4 (rarely 3), as: presutural: weak to hardly distinct, medium; postsutural: weak, weak, strong, strong. Presutural ac in 4–5 irregular rows. Legs dark. f2 with several (2–7) fine pv-setae in basal 1/3, the longest one (1.5 times as long as femoral width) at base. f3 at base with long (1.5–2 times as long as femoral width) and fine characteristic seta in v-pv position. Other setae on f3 variable: several av-setae present in apical third or slightly more, 1–3 among them longer, from as long as femoral width to twice as long; pv row may consist of 7–8 setae of which 2–3 are as long as femoral width, or may be reduced to 1–2 hardly distinct pv at base; sometimes chaetotaxy of f3 differs on right and left legs of the same specimen. Abdomen whitish-grey to yellowish-grey dusted, with paired L-shaped, more or less extensive dark spots on tergites 3 to 5, divided by dusted median vitta. Female differs from male as follows: parafacial more often yellowish; dc 1+4(3), anterior prst dc always absent; v-pv seta at base of f3 shorter to reduced; other ventral setae on f2 and f3 shorter or reduced.
The type locality of Lispe sericipalpis
is Indonesia, Java. The species has also been recorded from other
Indonesian islands (Bali, Sumatra), Taiwan, Vietnam, Thailand,
Myanmar, Sri Lanka, India, Pakistan, Nepal. The type locality of Lispe quaerens is Turkey, Akshehir prov. This species, as interpreted by
Over 350 specimens from a vast territory from the Iberian to Kamchatka Peninsulas.
Europe: Greece; Latvia; Portugal; Russia (European): Arkhangelsk, Chuvashia, Komi, Krasnodar, Kursk, Moscow, Murmansk, N. Ossetia, Nizhnyi Novgorod, Tula, Vladimir, Ulyanovsk; the Netherlands; Ukraine.
Asia: Abkhazia; Armenia; Azerbaijan; Kazakhstan: Almaty; Mongolia: Uvs prov.; Russia (Asian): Altai, Amur reg., Kamchatka, Khanty-Mansi reg., Krasnoyarsk reg., Magadan reg., Omsk reg., Primorsky Kray, Sakha (Yakutia) reg., Tyumen reg., Yamalo-Nenets reg.; Tajikistan: Dushanbe div., Khatlon div., Gorno-Badakhshan div.; Turkmenistan: Ahal, Mary; Turkey (Asian): Adana prov., Ankara prov., Antalya prov., Bolu prov., Duzce prov., Hatay prov., Isparta prov., Izmir prov., Kayseri prov., Konya prov., Mersin prov., Sakarya prov., Zonguldak prov.; Uzbekistan: Tashkent.
Lispe tentaculata has a variable t3 chaetotaxy: besides the normal strong ad and short pd setae, an additional seta just below the strong ad but in a more a-position is often present and sometimes a second strong ad and a short pd may also be present. About 5% of females have a male-like scutum: with only 3 strong post dc and without a pruinose patch on the scutum.
Lispe tentaculata is a very common species across its natural habitat. The northernmost specimens were collected near Murmansk (forest-tundra at 69°N) and Vorkuta (tundra at 67.5°N), where this species was found on boulders on river banks and was the only Lispe species recorded at these places. In southern Turkey at 36°N Lispe tentaculata was common again on boulders in streams and, in springtime, at various muddy places such as temporary pools. In temperate conditions of the East European Plain Lispe tentaculata is absent from small forest streams and is infrequent on the sandy banks of big rivers but is common on the mud ponds and small lakes. Both in temperate and southern habitats Lispe tentaculata distinctly avoids a salty mud where it is replaced by species from the Lispe caesia-group and the Lispe palposa-group. Lispe tentaculata is common in high mountain areas where it prefers boulders along the shores of mountain lakes; the series from Gorno-Badakhshan in Tajikistan was collected at 3800 m asl.
Lispe draperi was observed in Morocco in late March and seems to have an ecology similar to Lispe tentaculata.
Lispe consanguinea clearly prefers narrow sandy bands along the banks of big rivers. In this habitat Lispe consanguinea is the dominant species in the temperate zone, whereas Lispe tentaculata is uncommon. I had never collected Lispe consanguinea in localities north of the Arctic Circle, but on sandy banks of the Vychegda River at 61.3°N, 46.9°E this species was very common. The most southern records are Tajikistan (Dushanbe env., 1 specimen) and N. Turkey (2 specimens among numerous Lispe tentaculata).
Lispe sericipalpis (Fig. 5.1) may be found on boulders on the banks of rapid streams together with Lispe tentaculata. In N. Turkey this species is uncommon, whilst in S. Turkey Lispe sericipalpis is absent in spring time (late April), but becomes about as common as Lispe tentaculata in September. In Myanmar Lispe sericipalpis was absent in tree-shaded parts of streams, but was found in sunny sites at altitudes of 1100–1300 m asl.
1 Lispe sericipalpis Stein, male (Turkey, Antalya) 2 Lispe orientalis Wiedemann, male (Azerbaijan).
The ecology of Lispe orientalis (Fig. 5.2) is rather unusual: it is a species of dirty, organically-polluted water. Lispe orientalis was collected in Krasnodarsky Kray (Russia) and Antalya prov. (Turkey) around drinking bowls at pools polluted by cattle dung, in Tajikistan (Farkhor) at a very dirty irrigation ditch inside the town and at pools around this ditch polluted by refuse and even carrion, and in Hatay prov. (Turkey) at a strongly dung-polluted pool under a cattle shed.
In Azerbaijan, in late October, I observed Lispe orientalis during cold and bad weather, when it rained every day and usually all day long. In such conditions most flies appeared only after several hours without rain, while during the rain the only active dipteran was Lispe orientalis which hunted Diptera larvae on a wet chicken dunghill. It seems that Lispe orientalis has adapted to endure the rain and to keep on hunting because the larvae have to move up to the surface of the chicken manure and thus the prey becomes more accessible to the predator. In contrast to the related Lispe sericipalpis, Lispe orientalis prefers stagnant water. Only once near Çivlek (Turkey, Hatay prov.), I found Lispe orientalis by a rapid stream but shortly afterwards found a cattle shed in 50 metres upstream, which explained the presence of this species.
Lispe orientalis was regarded as an Oriental species, but the records listed above show that it is widespread in the South Palearctic and rather uncommon in the Oriental region where, for example, my dedicated Lispe collecting in Thailand made over several years has yielded only a single specimen of Lispe orientalis. The record from European Russia (Krasnodarsky Kray, vicinity of Sochi) is the northernmost one (43.4°N) and the first record for Europe, but I suspect that Lispe orientalis may be found in other European countries, being overlooked due to its omission from the keys for European Lispe.
FeedingIt is well known that all Lispe are predators (e.g.
Lispe draperi Séguy, male feeding on a dead Corixidae bug (Morocco, Essaouira).
I thank Adrian Pont (Oxford) for his invaluable help. Due to the kind help of Dutch colleagues Ben Brugge, Joke van Erkelens and Gerard Pennards (Amsterdam) I was able to examine type material of Lispe sericipalpis. I thank Oleg Kosterin (Novosibirsk) and Paul Beuk (Maastricht) who suggested several useful corrections.