





(C) 2010 Rahşen S. Kaya. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A new species of the spider genus Tegenaria Latreille, 1804 is described, based on newly collected specimens from Turkey. Detailed morphological descriptions, diagnosis and figures of the copulatory organs of both sexes are presented. Finally, a checklist and distribution maps for Turkish Tegenaria species are provided.
Agelenidae, new species, Tegenaria, Turkey
The spider family Agelenidae
currently constitutes 514 species in 42 genera and has a global
distribution (
In general, the supraspecific taxonomy of Tegenaria
and the tribe Tegenariini
is poorly resolved. The genus includes species with very different
palpal and epigynal conformations. Recently, Guseinov et al. (2005)
removed 26 species from Tegenaria
and transferred them to Malthonica
Simon, 1898. The taxonomy of Tegenaria
with respect to the Mediterranean fauna is currently being developed by
A. Bolzern (
During our surveys of the Turkish spider fauna, we found an undescribed species in the southern region of the country. This species possesses copulatory organs different from other Turkish and eastern Mediterranean Tegenaria and is described here as a new species.
Material and methodsThe specimens were studied using a Leica M205 C stereomicroscope.
The description of colour was based on live specimens. The epigyne was
macerated in 10% KOH. Measurements were taken with a micrometer eyepiece
from the dorsal aspect of the palps and legs. The morphological
terminology follows
Specimens were photographed using an Olympus Camedia E-520 camera attached to an Olympus SZX16 stereomicroscope. Images were produced using “CombineZP” image stacking software. Photographs were taken in dishes of different sizes with paraffin at the bottom. Holes of different sizes were made in the paraffin in order to keep specimens in the required position.
The following abbreviations are used:
AER anterior eye row; ALE anterior lateral eyes; AME anterior median eyes; PER posterior eye row; PLE posterior lateral eyes; PME posterior median eyes.
All measurements are in millimeters (mm).
Type specimens have been preserved in 70% ethanol and deposited in the Zoological Museum of Uludağ University (ZMUU, Department of Biology, Bursa, Turkey) and the Zoological Museum of the Moscow State University (ZMMU, Moscow, Russia; curator Dr K.G. Mikhailov).
DescriptionGenus Tegenaria Latreille, 1804
urn:lsid:zoobank.org:act:705E551F-CD34-4BAE-9757-48B8AC25B970
Figs 1-16, 20Holotype: male (ZMUU):Turkey: Antalya Province, Manavgat District, Beşkonak Village, Köprülü Canyon, 37°11'N; 31°11'E, 243 m a.s.l., 01.VI.2006, R. S. Kaya.

The general appearance and male palp of Tegenaria bayrami sp. n. 1 male, dorsal view 2 female, dorsal view 3 female prosoma, lateral view, showing long legs 4–5 male palp, ventral and retrolateral views respectively.
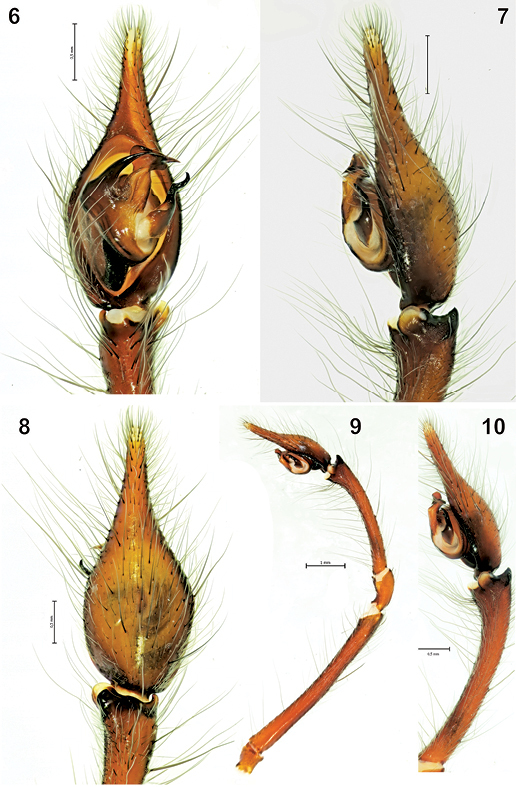
The male palp of Tegenaria bayrami sp. n. 6 ventral view 7 lateral view 8 dorsal view 9 entire palp, retrolateral view 10 tibia and tarsus, retrolateral view.
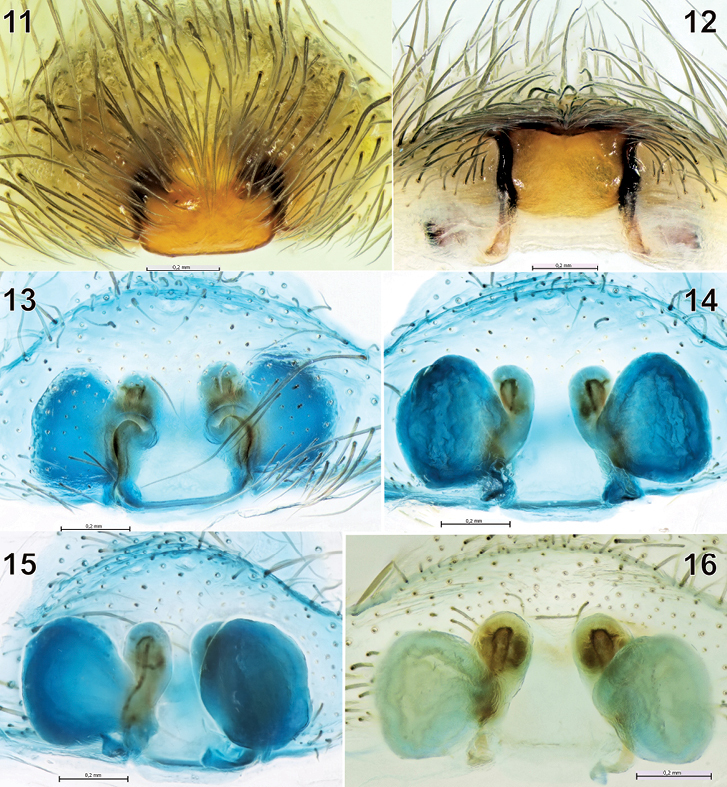
The epigyne and spermathecae of Tegenaria bayrami sp. n. 11 before maceration, ventral view 12 ditto, caudal view 13–14 after maceration, ventral and dorsal views 15 ditto, latero-dorsal view 16 ditto, dorsal view.
One male, one female and three subadult females (ZMUU), one male (ZMMU): same data as for the holotype, 21.VI.2010.
The species is named in honor of Prof. Dr. Abdullah Bayram, who has made an important contribution to Turkish arachnology.
Tegenaria bayrami sp. n. is closely related to Tegenaria longimana Simon, 1898, Tegenaria vignai Brignoli, 1978 and Tegenaria halidi Guseinov, Marusik & Koponen, 2005. Males of the new species can be easily distinguished from the mentioned species by the shape of the large and curved median apophysis and the pointed tip of the conductor (Figs 4, 6). The female of Tegenaria bayrami sp. n. can be separated from all other Tegenaria species by the square-shaped epigynal plate (Figs 11), short ducts and round spermathecae (Figs 14–16).
Male (holotype).Total length9.00.Prosoma: carapace 4.60 long, 3.50 wide. Carapace brownish yellow, with two longitudinal darkened bands (Fig 1), margins not darkened, scarce plumose hairs present. Cephalic region: 1.80 wide, darker and separated from the thoracic region by a distinct, darkened line. PER: 0.9 wide. Diameter of PME: 0.20; PLE: 0.22; AME: 0.10; ALE: 0.22. Distance of PME–PME: 0.10; PME–AME: 0.07. Eye formula: ALE=PLE>PME>AME. Clypeus height (measured from bottom of AME): 0.35, clypeus height (measured from bottom of ALE): 0.30. Clypeus deep reddish brown. Eye rows: AER slightly recurved, PER straight in dorsal view. Eye region darker. Chelicerae: 1.17 long; 0.90 wide. Chelicerae red-brown. Gnathocoxae: 1.32 long; 0.65 wide. Labium: 0.77 long; 0.70 wide. Gnathocoxae and labium brown. Sternum: 2.25 long; 2.22 wide. Sternum heart-shaped, pointed backwards, brown, with a light median band and three pairs of sublateral round spots. Legs light brown, with dark annulations, densely covered with plumose hairs; legs I and II are darker than legs III and IV (Fig. 1). Number of dorsal tarsal trichobothria on tarsi I and III: 10, tarsi II and IV: 9. Leg measurements are given in Table 1, and spine formulae in Table 2. Abdomen: 4.4 long, 2.6 wide; dorsum appears yellowish brown, with a reticulate patter of a series of transverse black lines along the dorsal mid-line and sides. Venter pale brown, with many short hairs and longitudinal black bars between the epigastric furrow and spinnerets.
Male palp as in Figs 4–10; very long (femur 5.10, patella 1.00, tibia 2.70, tarsus 2.20, (total 11.00), longer than body. Femur approximately 1.8 times longer than tibia, tibia 1.2 times longer than cymbium. Retrolateral tibial apophysis with two branches: lateral branch in dorsal view elongated and more or less rectangular, in retrolateral view triangular and tapering off towards the tip; broad and rounded latero-ventral branch with a small protuberance close to lateral branch in retrolateral view. Median apophysis long, in ventral view its base is large and broad, tip is claw-like. Conductor long, beak-shaped in ventral view; embolus thick and short in ventral view.
Female. Total length 11.40. Prosoma: carapace 5.40 long, 4.00 wide. Carapace light yellow, with two longitudinal darkened bands (Figs 2–3), margins not darkened, scarce hairs present. Cephalic region: 2.40 wide, darker and separated from the thoracic region by a distinct, darkened line. PER: 1.17 wide. Diameter of PME: 0.20; PLE: 0.22; AME: 0.12; ALE: 0.22. Distance of PME–PME: 0.20; PME–AME: 0.25. Eye formula: ALE=PLE>PME>AME. Clypeus height (measured from bottom of AME): 0.42, clypeus height (measured from bottom of ALE): 0.40. Clypeus dark brown. Eye rows: AER slightly recurved, PER straight in dorsal view. Chelicerae: 2.50 long; 1.30 wide. Chelicerae brown. Gnathocoxae: 1.50 long; 0.90 wide. Labium: 0.90 long; 0.80 wide. Gnathocoxae and labium brown, labium slightly longer than wide. Sternum: 2.60 long; 2.40 wide. Sternum heart-shaped, pointed towards rear end, brown, with light median band and three pairs of sublateral round spots. Legs light brown, with dark annulations, densely covered by long hairs, plumose hairs present. Number of dorsal tarsal trichobothria on tarsi I and IV: 11, tarsi II and III: 10. Leg measurements are given in Table 1, and spine formulae in Table 2. Abdomen: 6.00 long, 4.10 wide; dorsum yellowish brown, with a reticulate patter of a series of transverse, thick black lines along the dorsal mid-line and sides (Fig. 2). Venter pale brown, with many short hairs and longitudinal black bars between the epigastric furrow and spinnerets.
Leg and palp measurements of the holotype male and paratype female of Tegenaria bayrami sp. n.
| femur | patella | tibia | metatarsus | tarsus | total | ||
|---|---|---|---|---|---|---|---|
| male | palp | 5.1 | 1.0 | 2.7 | - | 2.2 | 11.0 |
| Leg I | 11.1 | 2.1 | 11.6 | 13.6 | 4.2 | 42.6 | |
| Leg II | 9.7 | 1.9 | 9.6 | 12.0 | 3.6 | 36.8 | |
| Leg III | 8.2 | 1.8 | 7.8 | 10.5 | 3.0 | 31.3 | |
| Leg IV | 9.6 | 1.8 | 9.4 | 13.4 | 3.4 | 37.6 | |
| female | palp | 3.3 | 1.2 | 2.2 | - | 2.7 | 9.4 |
| Leg I | 9.7 | 2.1 | 8.8 | 11.4 | 3.7 | 35.7 | |
| Leg II | 8.8 | 2.0 | 7.9 | 9.6 | 3.2 | 31.5 | |
| Leg III | 7.2 | 1.6 | 6.6 | 8.5 | 3.0 | 26.9 | |
| Leg IV | 9.4 | 2.0 | 8.2 | 11.1 | 3.2 | 33.9 | |
Spination of legs and palps of Tegenaria bayrami sp. n. The formula gives the number of spines in the following order: dorsal – prolateral – retrolateral – ventral. The letter ‘p’ indicates a pair of spines that occur at this position.
| femur | patella | tibia | metatarsus | tarsus | ||
|---|---|---|---|---|---|---|
| palp | Paratype (female) | 2-1-1-0 | 2-0-0-0 | 2-2-0-0 | - | many |
| leg I | Holotype (male) | 1-2-3-0 | 1-0-0-0 | 0-1-1-1 | 0-1-0-1p+1+1p | 0-0-0-0 |
| Paratype (female) | 2-3-2-0 | 2-0-0-0 | 1-1-1-1 | 1-1-1-1p+1p+1p | 0-0-0-0 | |
| leg II | Holotype (male) | 2-3-2-0 | 1-0-0-0 | 1-2-1-1+1p | 0-2-1-1p+1+1p | 0-0-0-0 |
| Paratype (female) | 2-3-2-0 | 1-0-0-0 | 1-2-0-1p+1p | 0-2-1-1p+1p+1p | 0-0-0-0 | |
| leg III | Holotype (male) | 2-2-2-0 | 1-0-0-0 | 1-2-1-1p+1p+1p | 1-3-3-1p+1+1+1p | 0-0-0-0 |
| Paratype (female) | 4-5-4-0 | 1-0-0-0 | 1-2-1-1p+1p+1p | 1-4-4-1p+1+1+1p+1p | 0-0-0-0 | |
| leg IV | Holotype (male) | 2-2-3-0 | 1-0-0-0 | 1-2-2+1p+1p | 2-3-4-1+1+1+1p | 0-0-0-0 |
| Paratype (female) | 2-2-3-0 | 2-0-0-0 | 2-3-2-1p+1+1+1p | 2-3-4-1p+1p+1p | 0-0-0-0 | |
Epigyne and spermathecae as in Figs 11-16. Fovea absent, median plate square-shaped; copulatory openings almost invisible on intact epigyne, but readily visible following hair removal. Insemination duct short, spermathecae almost round.
The new species was collected from damp places of rocky areas along the River Köprüçay (Köprülü Canyon, Antalya). The canyon is located on the lower slopes of the West Taurus Mountain ranges. Samples were collected from their big funnel webs during the day. The collection of a male in copula with a female clearly suggests that both sexes described here are conspecific.
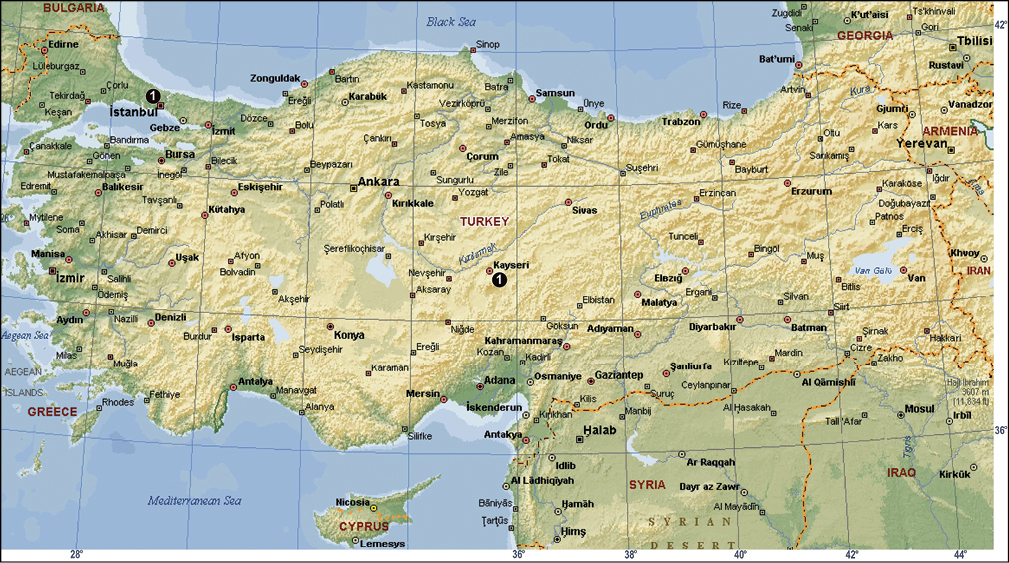
Turkey, known only from the type locality (Fig. 20).
Figs 17-20
Fig. 20
Turkey.
Antalya Province: Döşemealtı District, Mustanini Cave (
Fig. 17
Europe to Central Asia, USA and Canada.
Anatolia, no exact locality. It was reported from Turkey by
Fig. 18
Europe, introduced to North America.
İstanbul Province: Şile District; Kayseri Province: Yeşilhisar District, Harmankaya Cave (Roewer 1959). It is likely that this species was misidentified from Turkish specimens and probably does not occur in the country.
Fig. 20
Turkey.
Mersin Province: Silifke District, Cennet Cave (
Fig. 20
Turkey only.
Antalya Province: Manavgat District, Beşkonak Village, Köprülü Canyon.
Fig. 19
Bulgaria and Turkey.
Bolu Province: Abant (
Fig. 20
Turkey.
Trabzon Province: Maçka District, Sümela Monastery (
Fig. 20
Turkey.
Rize Province: Kalkandere District; Artvin Province: Borçka
District (
Fig. 17
Cosmopolitan, synanthropic in most places.
Hatay Province: Narlıca Town, Narlıca Cave; Urfa Province;
Mardin Province: Midyat District, a hill near Derömer Area (
Fig. 20
Turkey.
Mersin Province: Silifke District, Dilek Cave and Cennet
Cave (
Fig. 20
Turkey.
Hatay Province: Harbiye Town, Harbiye Cave (
Fig. 20
Turkey.
Konya Province: Beyşehir District, Kurucuova Village
(Asarini Cave and İnönüini Cave); Seydişehir District (Ferzene Cave and
fossile sinkhole of Tınaztepe Cave); Çamlık District (Körükini Cave);
Hadım District (Suçıktığı Cave); Antalya Province: Akseki District
(Demirci Dükkanları Cave, a cave in Dikmen Village and from a well) (
Fig. 20
Turkey.
Isparta Province: Eğirdir District (Brignoli 1978b).
Fig. 20
Turkey.
Konya Province: Seydişehir District, Ferzene Cave (
Fig. 19
Turkey, Georgia and Russia.
Rize Province: Fındıklı District (50 km NE of Rize, towards Hopa); Artvin Province: Hopa District; Trabzon Province: Maçka District, Altındere Village, Sümela Monastery (Brignoli, 1978b).
Fig. 20
Turkey.
Artvin Province: Yalnızçam Pass (Brignoli 1978b).
Fig. 20
Turkey.
Diyarbakır Province: Lice Province, Korkha Cave (
Fig. 17
Europe, North Africa to Central Asia, Uruguay and Argentina.
İstanbul Province (
Fig. 20
Turkey.
Isparta Province: Aksu District, Anamas Plateau, Zindan
Cave (
Fig. 19
Rhodes and Turkey.
Konya Province: Beyşehir District, Beyşehir Lake, Island of Hacı Akif; Isparta Province: Eğirdir District (Brignoli 1978b).
Fig. 20
Turkey.
Antalya Province: Around Kaş District (Brignoli 1978b).
Fig. 20
Turkey.
Artvin Province: Borçka District (Brignoli 1978b).
Fig. 20
Turkey.
Trabzon Province: Maçka District (Sümela Monastery) and
Zigana Pass (
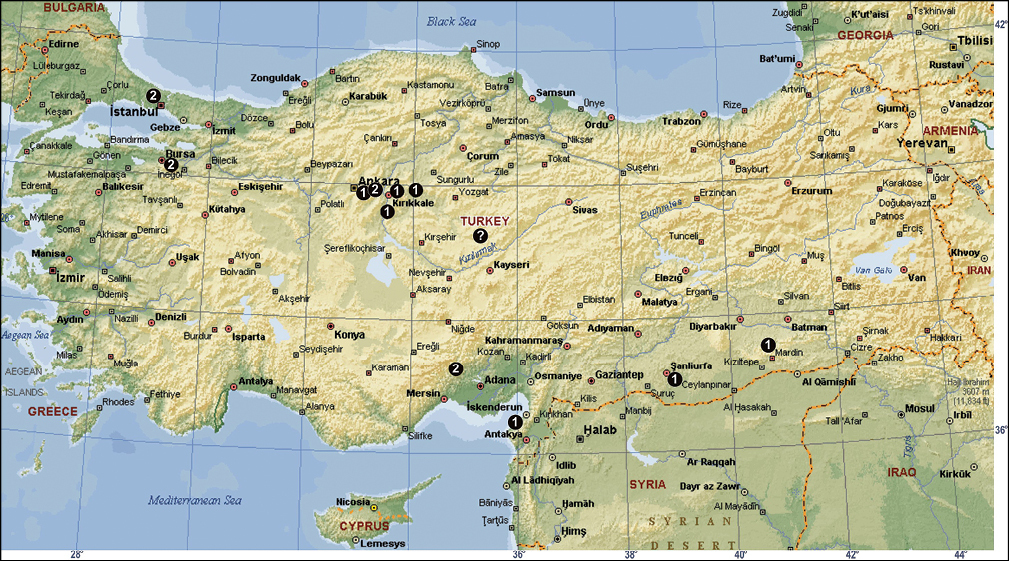
The distribution of Tegenaria agrestis (? = no exact locality in Anatolia; see Caporiacco, 1935), Tegenaria domestica (1) and Tegenaria parietina (2) in Turkey.
The distribution of Tegenaria atrica (1) in Turkey.
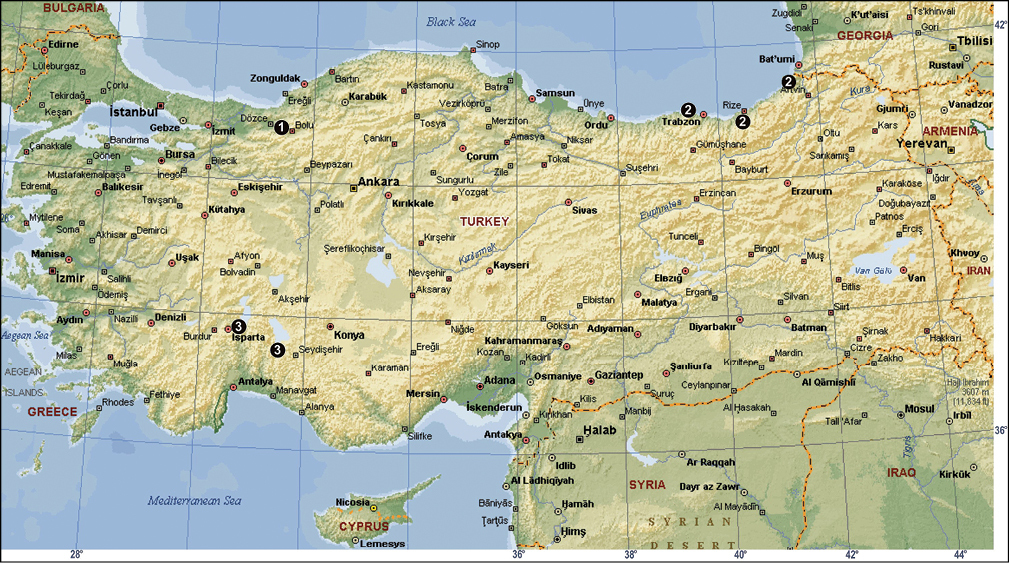
The distribution of Tegenaria bithyniae (1), Tegenaria longimana (2) and Tegenaria rhodiensis (3) in Turkey.
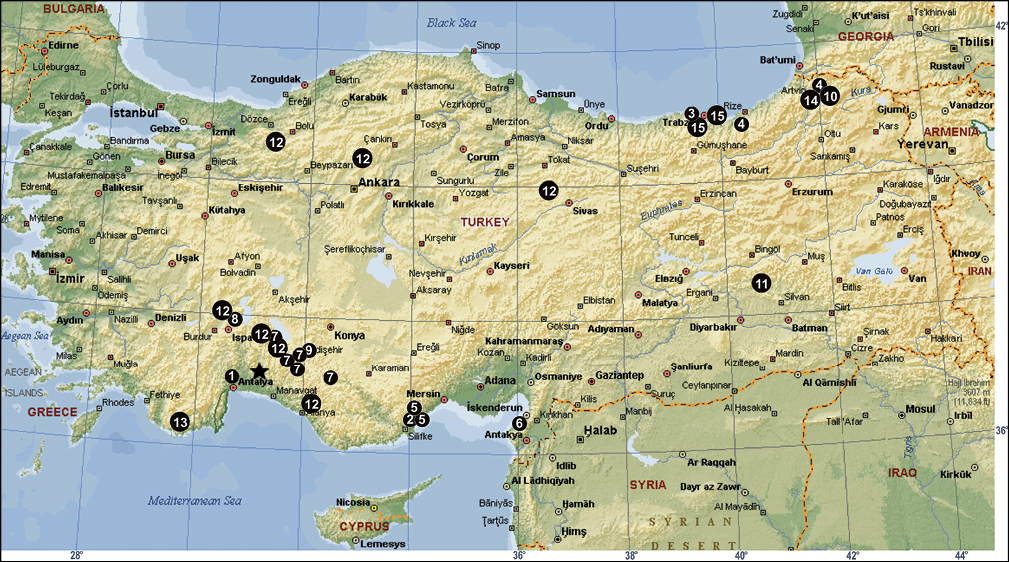
The distribution of Tegenaria agnolettii (1), Tegenaria averni (2), Tegenaria comnena (3), Tegenaria cottarellii (4), Tegenaria elysii (5), Tegenaria faniapollinis (6), Tegenaria forestieroi (7), Tegenaria hamid (8), Tegenaria karaman (9), Tegenaria mamikonian (10), Tegenaria melbae (11), Tegenaria percuriosa (12), Tegenaria tekke (13), Tegenaria vignai (14), Tegenaria xenophontis (15) and Tegenaria bayrami sp. n. (star) in Turkey.
Twenty-three Tegenaria
species have now been reported from Turkey, including the newly
described species. Only four of them (Tegenaria agrestis,
Tegenaria atrica,
Tegenaria domestica
and Tegenaria parietina)
have broad distribution ranges, whereas 16 species are endemic to
Turkey. Among the species restricted to Turkey or to the eastern
Mediterranean, 14 are known exclusively from females and only four
species are known from both sexes. All species known from only one sex
were described by
Although the currently known diversity of Tegenaria and Malthonica in Turkey is already extraordinarily high, the actual diversity may be even higher. Many caves, a favourite habitat for Tegenaria, have never been studied or sampled for spiders in Turkey. Therefore, we expect that more new species will be found in the future.
The authors would like to thank Yaprak Gürkan and biology students (Uludağ University, Science and Art Faculty, Department of Biology) for their valuable assistance during field trips. Also, we would like to thank Prof. Dr. Gökay Kaynak (Uludağ University, Science and Art Faculty, Department of Physics) for allowing us to use Leica M205 C Stereomicroscope (Research Foundation of Uludağ University Project No: F-2005/4). Dr David Penney is thanked for his kind help in editing the English. This work was supported in part by the RFFI grant # 09-04-01365_a.