





(C) 2010 Angelica Crottini. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
We describe a new mantellid frog of the subfamily Mantellinae from the karstic Bemaraha Plateau, western Madagascar. The new species belongs to the genus Gephyromantis, subgenus Phylacomantis, which previously included Gephyromantis azzurrae, Gephyromantis corvus and Gephyromantis pseudoasper. Gephyromantis atsingy sp. n. has a snout-vent length of 35–43 mm and is a scansorial frog living among the Tsingy de Bemaraha pinnacles and inside the caves present in the area. A morphological analysis and biomolecular comparison revealed the degree of differentiation between these four species of the Phylacomantis subgenus.The new species seems to be endemic to Tsingy de Bemaraha.
Amphibia, Gephyromantis atsingy sp. n., Madagascar, Tsingy de Bemaraha
The intense herpetological activity carried out in
Madagascar during the last decades, together with the wider use of
integrative taxonomic tools has led to the description of an
astonishingly high number of new amphibians species (
Although, the highest species richness of amphibians is typically found along the eastern rainforest belt (
While the species already described from the arid West
mostly belong to radiations of explosive breeders reproducing in
ephemeral ponds, a special attention has been given to species ascribed
to clades that are more typical of humid habitats and rainforest
biomes. This was the case, for example, with the recent discovery of
two new mantellines at the Isalo Massif (
During recent herpetofaunal inventories we discovered a further new species of a rather inconspicuous Gephyromantis frog inhabiting the deciduous forest of the karstic Bemaraha Plateau.
Thirty-six described species are currently ascribed to Gephyromantis that is currently divided in five subgenera, including Phylacomantis. Four species are currently ascribed to this subgenus: Gephyromantis corvus Glaw & Vences, Gephyromantis pseudoasper Guibé, Gephyromantis azzurrae Mercurio & Andreone and the new species described in the present paper. With the exception of Gephyromantis pseudoasper, that mostly inhabits the rainforests of the North, the other species are found only in xeric habitats in the south-western (Gephyromantis corvus and Gephyromantis azzurrae) and western Madagascar (the new species described herein) (Fig. 1).
Due to morphological and external similarities, the new frog was formerly believed to be related to Gephyromantis corvus, a frog endemic of the Isalo Massif.
Unfortunately, the secretive life of this new species
prevented us from obtaining much biological informations and we still
lack information about its acoustic repertoire, breeding behaviour and
larval morphology. Notwithstanding these challenges, the ongoing
collaborative effort generated by the ACSAM (A Conservation Strategy for
the Amphibians of Madagascar,
We present here the formal description of this new Gephyromantis species of the subgenus Phylacomantis, which differs from the other Phylacomantis species by a combination of morphological traits colouration and by a high divergence in mitochondrial DNA sequences.
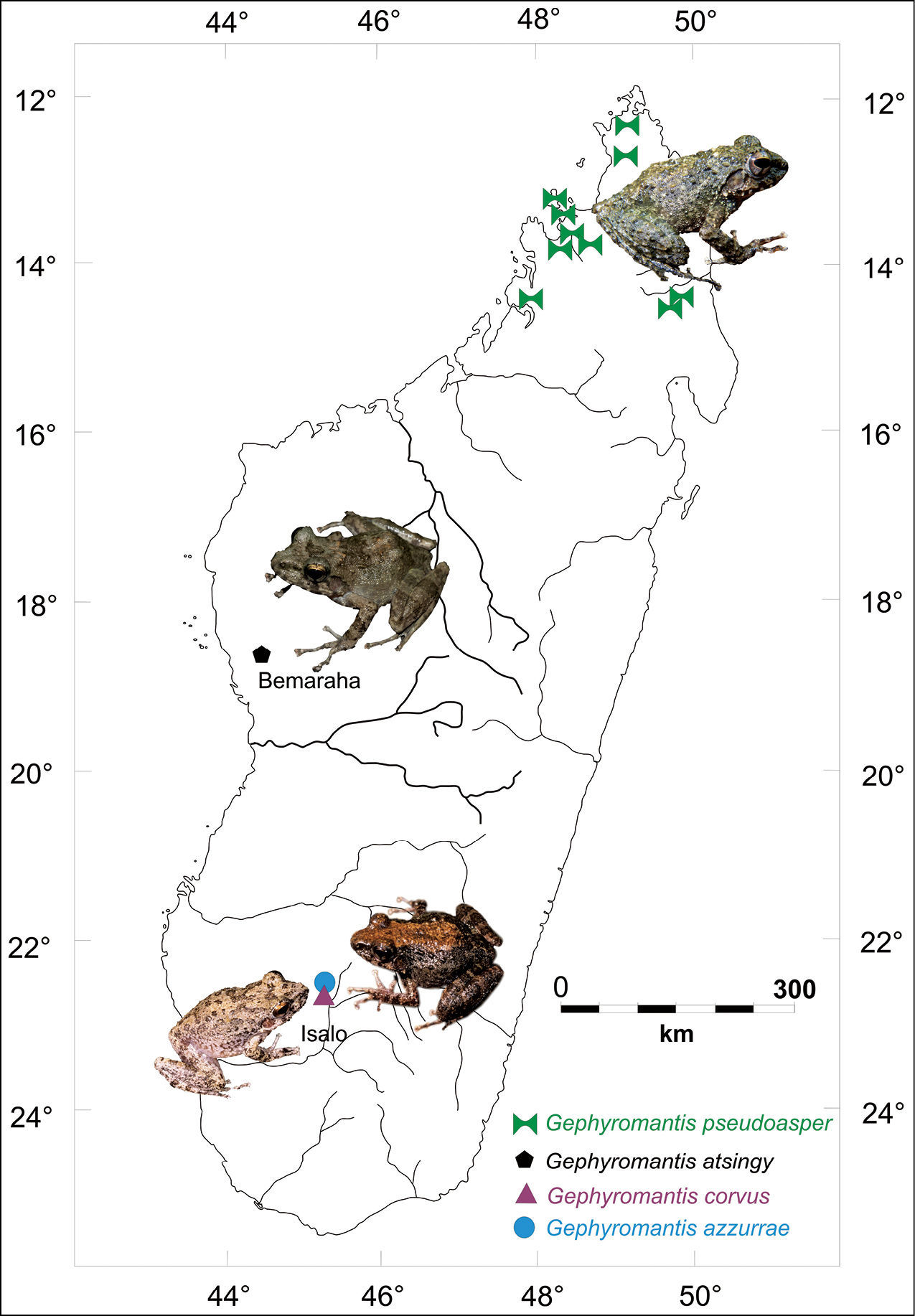
Schematic map of Madagascar with images and distribution of the four described species of the genus Gephyromantis, subgenus Phylacomantis.
The Tsingy de Bemaraha is a karstic plateau in the
Melaky Region, five to 15 km wide and about 100 km long, located in
western Madagascar. Numerous long, sharp pinnacles of rock, that may
reach 45 meters in height, outcrop along the plateau and form the
characteristic landscape (the so-called “tsingy” or “atsingy” in
Malagasy language). Dry, deciduous forest is the most common vegetation
type but humid areas occur within some of the larger canyons. An
extensive area of forest and rock outcrop is included within two
adjacent protected areas (Parc National Tsingy de Bemaraha and Réserve
Naturelle Intégrale du Tsingy de Bemaraha). Savanna grasslands surround
the plateau and there are numerous marshy depressions, caves and gorges
associated with the main outcrop. This area has been object of some
herpetological surveys that led to the discovery and description of
several new species of amphibians (
We searched for frogs at night with the aid of hand
torches and headlamps. Geographic coordinates were taken using a GPS
device. Toponyms often follow the indications by local people, and must
be therefore seen as largely unofficial names. Frogs were collected by
hand and euthanised by immersion in chlorobutanol solution, fixed in 5%
formalin or in 90% ethanol and finally stored in 75% ethanol solution.
Voucher specimens (Tab. 1)
are currently housed at the Museo Regionale di Scienze Naturali di
Torino (MRSN), Zoologische Staatssammlung München (ZSM), and
Université d’Antananarivo, Département de Biologie Animale (UADBA).
Original field numbers are FN and FAZC (Franco Andreone Zoological
Collection), FGZC (Frank Glaw Zoological Collection), BMR (Jasmin E.
Randrianirina), and RBJ (Richard K. B. Jenkins). A few individuals do
not bear any field number (no field number = NFN). The specimens of the
type series were compared with the specimens of the other known species
of the Phylacomantis subgenus (Tab. 1): six specimens of Gephyromantis azzurrae from Isalo, five specimens of Gephyromantis corvus from Isalo, and three specimens of Gephyromantis pseudoasper from Nosy Be (see Tab. 1). The comparative specimens of Gephyromantis pseudoasper
were most probably temporary stored in denaturating solutions, and the
sequencing was therefore not successful. For this reason, the three
specimens here analyzed were only compared morphologically (Tab. 1),
and the required sequences were retrieved from GenBank (DQ987513,
DQ987515, DQ987517, DQ987518; DQ926890; AY848422-AY848424).
Morphological information on Gephyromantis azzurrae specimens were taken from
Morphological measurements were made with a digital
calliper to the nearest 0.1 mm. The following biometric measurements
were taken (according to
A fingertip, or part of the muscle of the tongue,
was cut from each collected individual and stored in 99% ethanol. Total
genomic DNA was extracted from the tissue samples using proteinase K
digestion (10 mg/ml concentration) following
urn:lsid:zoobank.org:act:94995B02-B47C-4275-A6BA-DD4134B51203
Figures 2, A–LThe specific noun “atsingy” (pronounced: “a-tseen-jě”) is a Malagasy word. The terms “atsingy” or “tsingy” are the common names used to refer to the pointed and sharp calcareous lime stone formations and pinnacles originated through rainfall erosion. Although present in several other localities in western Madagascar (e.g.: Ankarana), the outcrops of Bemaraha are typical of this area and the specific name is therefore associated with the locality of provenience of the types.
This species has been referred to as Gephyromantis sp. aff. corvus “Bemaraha” by
MRSN A5487 (NFN), subadult male, collected at Tsingy de Bemahara National Park, western Madagascar, Andamozavaky (Bekopaka commune, Antsalova district, Melaky region, Mahajanga province), 19°01.86'S, 44°46.80'E; 122 m a.s.l., collected by J. E. Randrianirina on 23 May 2003.
MRSN A5486 (BMR 001), subadult male without evident femoral glands, MRSN A5484 (NFN), adult female, MRSN A5482 (BMR 008), MRSN A5483 (BMR 031), MRSN A5485 (BMR 002), three juveniles (sex unknown) sampled from the same locality, collector and date of the holotype (tissue sample taken for genetical analysis for all individuals); ZSM 23/2006 (FGZC 0715), adult female, from Grotte Crystal, close to Andranopasazy, Tsingy de Bemaraha National Park (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°42'31"S, 44°43'08"E, 146 m a.s.l., collected by F. Glaw, J. Köhler, P. Bora and H. Enting on 19 March 2006, fixed in ethanol (tissue sample taken for genetical analysis), individual found at night on limestone cliffs, close to the entrance of the cave; ZSM 37/2006 (FGZC 0746), juvenile (unknown sex) from Grotte Anjohimbazimba, Tsingy de Bemaraha National Park (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°41'34"S, 44°42'36"E, 160 m a.s.l., collected by F. Glaw, J. Köhler, P. Bora and H. Enting on 20 March 2006 (tissue sample taken for genetical analysis), individual found in the cave; ZSM 107/2006 (FGZC 0886), juvenile (sex unknown) from Bendrao Forest (“Camp 3”), Tsingy de Bemaraha National Park (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°47'04"S, 44°51'37"E, 427 m a.s.l., collected by F. Glaw, J. Köhler, P. Bora and H. Enting on 26–27 March 2006; (tissue sample taken for genetical analysis). All these specimens were fixed in 90% ethanol and preserved in 70% ethanol. UADBA 28112 (RBJ 708), female from Ranotsara (Bekopaka commune Antsalova district, Melaky region, Mahajanga province), 19°02'08"S, 44°46'29"E, 65 m a.s.l., collected by R. Andriantsimanarilafy on 18 November 2006; UADBA 28116 (RBJ 792), female from Ankilogoa (Bekopaka commune, Antsalova district, Melaky region, Mahajanga province), 19°07'52"S, 44°48'32"E, 57 m a.s.l., collected by R. Randrianavelona on 13 December 2006; UADBA 28120 (RBJ 791), female from Ankilogoa (Bekopaka commune, Antsalova district, Melaky region, Mahajanga province), 19°07'52"S, 44°48'32"E, 57 m a.s.l., collected by R. Randrianavelona on 13 December 2006; UADBA 28127 (RBJ 718), female from Ranotsara (Bekopaka commune, Antsalova district, Melaky region, Mahajanga province), 19°02'08"S, 44°46'29"E, 65 m a.s.l., collected by R. Randrianavelona on 19 November 2006; UADBA 39057 (RBJ 660), female from Anjaha (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°39'43"S, 44°49'33"E, 403 m a.s.l., collected by J.C. Randrianantoandro, R. Randrianavelona, R.K.B. Jenkins, R.R. Andriantsimanarilafy and E.F. Hantalalaina and Madagascar National Parks personnel on 15–24 February 2006; UADBA 39081 (RBJ 609), female from Andranopasazy (Melaky region, Mahajanga province), 18°42'31"S, 44°43'02"E, 146 m a.s.l. collected by J.C. Randrianantoandro, R. Randrianavelona, R.K.B. Jenkins, R.R. Andriantsimanarilafy and E.F. Hantalalaina and Madagascar National Parks personnel on 13–30 January 2006; UADBA 39082 (RBJ 630), female from Andranopasazy (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°42'31"S, 44°43'02"E, 146 m a.s.l. collected by J.C. Randrianantoandro, R. Randrianavelona, R.K.B. Jenkins, R.R. Andriantsimanarilafy and E.F. Hantalalaina and Madagascar National Parks personnel on 13–30 January 2006; UADBA 39099 (RBJ 627), adult male (with developed glands) from Andranopasazy (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°42'31"S, 44°43'02"E, 146 m a.s.l. collected by J.C. Randrianantoandro, R. Randrianavelona, R.K.B. Jenkins, R.R. Andriantsimanarilafy and E.F. Hantalalaina and Madagascar National Parks personnel on 13–30 January 2006; UADBA 39100 (RBJ 658), female from Anjaha (Antsalova commune, Antsalova district, Melaky region, Mahajanga province), 18°39'43"S, 44°49'33"E, 403 m a.s.l., collected by J.C. Randrianantoandro, R. Randrianavelona, R.K.B. Jenkins, R.R. Andriantsimanarilafy and E.F. Hantalalaina and Madagascar National Parks personnel on 15–24 February 2006.

Images of Gephyromantis atsingy sp. n. A MRSN A5487 (NFN), subadult male (holotype) from Andamozavaky, dorsal view (photo by J. E. Randrianirina) B–C ZSM 23/2006 (FGZC 0715), adult female (paratype) from Grotte Crystal, close to Andranopasazy, dorsolateral views (photos by F. Glaw) D ZSM 23/2006 (FGZC 0715), adult female (paratype) from Grotte Crystal, close to Andranopasazy, ventral view (photo by F. Glaw) E ZSM 37/2006 (FGZC 0746), juvenile (paratype) from Grotte Anjohimbazimba (photo by F. Glaw) F–G UADBA 39099 (RBJ 627), adult male (paratype) from Andranopasazy, dorsolateral and dorsal views (photos by C. Randrianantoandro) H UADBA 39099 (RBJ 627), adult male (paratype) from Andranopasazy, ventral view, with evident and developed femoral glands of “Type 2” (photo by C. Randrianantoandro) I ZSM 107/2006 (FGZC 0886), juvenile (paratype) from Bendrao Forest (“Camp 3”), dorsolateral view (photo by F. Glaw) J MRSN A5483 (BMR 031), juvenile (paratype) from Andamozavaky, dorsolateral view (photo by J. E. Randrianirina) K–L MRSN A5487 (NFN), subadult male (holotype) from Andamozavaky, dorsal and ventral views of the preserved specimen.
A medium sized frog species (adult SVL 35–43 mm), assigned to the genus Gephyromantis (sensu
Morphometric measurements (in mm) of specimens of Gephyromantis atsingy, Gephyromantis corvus, Gephyromantis azzurrae and Gephyromantis pseudoasper. HT (holotype), PT (paratype). M (male). F (female). J (juvenile), SMF (Forschungsinstitut und Naturmuseum Senckenberg, Frankfurt, Germany). Other abbreviations are given in the text.
| Catalogue number | Field number | Species | Locality | GenBank | Rank | SEX | SVL | HW | HL | ED | END | NSD | NND | TD | HAL | HIL | FORL | FOTL | FOL | IMTL | IMTH | FGL | FGW | NG | GD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MRSN A5487 | NFN | G. atsingy | Andamozavaky | HQ640419 | HT | M | 34.8 | 14.1 | 14.5 | 5.7 | 3.8 | 2.6 | 3.6 | 3.3 | 11.6 | 37.5 | 18.6 | 24.8 | 17.7 | 1.1 | 1.1 | - | - | 5 | 0.7 |
| MRSN A5486 | BMR 001 | G. atsingy | Andamozavaky | HQ640421 | PT | M | 31.3 | 12.4 | 13.5 | 5.3 | 3.9 | 1.9 | 3.1 | 2.8 | 10.6 | 33.1 | 16.2 | 23.6 | 15.5 | 1.7 | 1.2 | - | - | - | - |
| MRSN A5484 | NFN | G. atsingy | Andamozavaky | HQ640418 | PT | F | 43.4 | 15.9 | 17.3 | 6.4 | 4.5 | 3.1 | 3.9 | 3.6 | 13.3 | 43.2 | 20.1 | 30.9 | 20.4 | 2.2 | 1.0 | - | - | - | - |
| ZSM 23/2006 | FGZC 0715 | G. atsingy | Grotte Crystal, close to Andranopasazy | HQ640414 | PT | F | 38.5 | 14.3 | 15.2 | 6.1 | 4.3 | 2.2 | 3.6 | 2.7 | 11.7 | 39.9 | 18.0 | 26.0 | 16.7 | 1.6 | 0.9 | - | - | - | - |
| MRSN A5482 | BMR 008 | G. atsingy | Andamozavaky | HQ640420 | PT | J | 22.0 | 8.4 | 9.6 | 3.8 | 2.6 | 1.6 | 2.5 | 2.2 | 7.4 | 23.7 | 11.1 | 17.1 | 9.9 | 1.1 | 0.6 | - | - | - | - |
| MRSN A5483 | BMR 031 | G. atsingy | Andamozavaky | HQ640417 | PT | J | 19.6 | 7.3 | 8.3 | 3.5 | 2.2 | 1.1 | 2.1 | 1.7 | 6.3 | 17.9 | 9.4 | 13.3 | 8.1 | 0.4 | 0.2 | - | - | - | - |
| MRSN A5485 | BMR 002 | G. atsingy | Andamozavaky | HQ640415 | PT | J | 23.6 | 8.9 | 10.5 | 4.1 | 2.9 | 1.4 | 2.6 | 2.3 | 8.1 | 25.5 | 13.2 | 18.4 | 11.6 | 0.6 | 0.2 | - | - | - | - |
| ZSM 107/2006 | FGZC 0886 | G. atsingy | Bendrao Forest | HQ640416 | PT | J | 22.5 | 7.8 | 9.5 | 3.1 | 2.4 | 0.9 | 2.1 | 1.8 | 6.5 | 22.4 | 10.9 | 16.5 | 10.0 | 1.0 | 0.3 | - | - | - | - |
| ZSM 37/2006 | FGZC 0746 | G. atsingy | Grotte Anjohimbazimba | - | PT | J | 24.6 | 8.2 | 10.1 | 3.2 | 2.7 | 1.5 | 2.3 | 1.9 | 6.9 | 24.2 | 11.1 | 17.1 | 10.8 | 1.1 | 0.5 | - | - | - | - |
| UADBA 39099 | RBJ 627 | G. atsingy | Andranopasazy | HQ640413 | PT | M | 36.6 | 11.2 | 18.6 | 4.2 | 3.6 | 1.9 | 2.9 | 3.4 | 11.5 | 62.1 | 16.4 | 25.5 | 17.5 | 1.1 | 0.8 | 7.5 | 3.1 | 70 | 0.5 |
| UADBA 39081 | RBJ 609 | G. atsingy | Andranopasazy | - | PT | F | 38.4 | 14.8 | 16.8 | 4.3 | 3.6 | 1.9 | 3.4 | 3.3 | 11.9 | 63.5 | 17.4 | 27.3 | 19.3 | 1.4 | 0.8 | - | - | - | - |
| UADBA 28120 | RBJ 791 | G. atsingy | Ankilogoa | - | PT | F | 35.9 | 11.1 | 17.1 | 4.5 | 3.3 | 1.6 | 2.6 | 3.2 | 11.1 | 64.0 | 16.9 | 27.1 | 18.1 | 1.1 | 0.5 | - | - | - | - |
| UADBA 39100 | RBJ 658 | G. atsingy | Anjaha | - | PT | F | 35.1 | 12.1 | 17.8 | 4.4 | 3.9 | 1.7 | 2.7 | 3.3 | 11.0 | 62.2 | 17.3 | 26.8 | 17.3 | 1.3 | 0.7 | - | - | - | - |
| UADBA 28127 | RBJ 718 | G. atsingy | Ranotsara | - | PT | F | 39.1 | 12.7 | 17.9 | 4.6 | 3.7 | 1.6 | 3.2 | 3.3 | 12.1 | 64.0 | 18.4 | 28.4 | 18.7 | 1.4 | 0.6 | - | - | - | - |
| UADBA 39082 | RBJ 630 | G. atsingy | Andranopasazy | - | PT | F | 38.1 | 12.6 | 16.9 | 3.9 | 4.1 | 1.6 | 2.9 | 2.9 | 11.6 | 63.9 | 17.8 | 27.4 | 17.6 | 1.5 | 0.5 | - | - | - | - |
| UADBA 28112 | RBJ 708 | G. atsingy | Ranotsara | - | PT | F | 38.4 | 11.8 | 16.8 | 4.2 | 4.0 | 1.1 | 2.5 | 3.0 | 11.4 | 61.9 | 18 | 26.3 | 16.4 | 1.2 | 0.5 | - | - | - | - |
| UADBA 39057 | RBJ 660 | G. atsingy | Anjaha | - | PT | F | 35.3 | 11.4 | 17.4 | 4.3 | 3.6 | 1.4 | 2.9 | 2.8 | 11.6 | 63.9 | 16.6 | 27.5 | 18.4 | 0.9 | 0.5 | - | - | - | - |
| UADBA 28116 | RBJ 792 | G. atsingy | Ankilogoa | - | PT | F | 33.9 | 11.0 | 16.3 | 3.4 | 3.7 | 1.2 | 2.2 | 2.5 | 11.0 | 58.6 | 16.4 | 25.0 | 15.9 | 1.0 | 0.5 | - | - | - | - |
| MRSN A5373 | FAZC 12859 | G. corvus | Isalo, Tsiombivositra | HQ640423 | - | M | 39.8 | 16.1 | 15.3 | 5.8 | 4.8 | 2.9 | 4.3 | 3.7 | 11.8 | 38.7 | 16.3 | 28.6 | 19.6 | 2.4 | 1.4 | 9.1 | 4.1 | 96 | 0.6 |
| MRSN A5325 | FAZC 13000 | G. corvus | Isalo, Ambovo | - | - | F | 40.0 | 15.2 | 16.2 | 6.4 | 4.3 | 2.6 | 4.1 | 3.2 | 10.7 | 40.3 | 18.2 | 30.1 | 20.0 | 1.6 | 0.6 | - | - | - | - |
| MRSN A5323 | FAZC 12661 | G. corvus | Isalo, Malaso | HQ640422 | - | F | 39.0 | 15.0 | 15.7 | 6.1 | 4.4 | 2.8 | 4.2 | 3.6 | 11.9 | 39.3 | 17.6 | 29.7 | 20.3 | 1.6 | 0.8 | - | - | - | - |
| MRSN A5324 | FAZC 12758 | G. corvus | Isalo, Zahavola | HQ640424 | - | F | 40.1 | 15.0 | 16.3 | 6.1 | 4.4 | 2.8 | 4.2 | 3.3 | 10.8 | 37.2 | 16.9 | 28.0 | 19.8 | 1.3 | 0.7 | - | - | - | - |
| MRSN A2786 | FAZC 11964 | G. corvus | Isalo, Andranomena | HQ640425 | - | F | 40.8 | 15.0 | 15.8 | 6.3 | 4.7 | 2.7 | 4.3 | 3.6 | 11.1 | 40.5 | 17.7 | 30.2 | 20.4 | 1.9 | 0.7 | - | - | - | - |
| MRSN A5310 | FAZC 12568 | G. azzurrae | Isalo, Andriamanero | EF222301 | HT | M | 41.1 | 16.9 | 13.4 | 6.1 | 3.9 | 2.5 | 4.0 | 4.0 | 12.1 | 41.1 | 20.0 | 30.0 | 18.8 | 2.0 | 1.1 | 6.3 | 2.0 | 45 | 0.5 |
| MRSN A5309 | FAZC 12567 | G. azzurrae | Isalo, Andriamanero | EF222300 | PT | M | 38.5 | 15.3 | 12.8 | 5.2 | 4.3 | 2.2 | 3.7 | 3.7 | 11.1 | 41.1 | 19.9 | 26.7 | 18.9 | 1.9 | 1.3 | 6.5 | 2.7 | 38 | 0.5 |
| MRSN A5311 | FAZC 12569 | G. azzurrae | Isalo, Andriamanero | EF222302 | PT | M | 40.2 | 15.8 | 14.1 | 6.0 | 4.0 | 2.7 | 4.0 | 4.1 | 11.2 | 41.0 | 19.9 | 27.7 | 19.9 | 2.1 | 1.1 | 6.7 | 2.7 | 40 | 0.6 |
| MRSN A5312 | FAZC 12910 | G. azzurrae | Isalo, Iambahatsy | EF222304 | PT | M | 23.3 | 8.8 | 8.8 | 4.1 | 2.8 | 1.4 | 2.2 | 2.5 | 8.8 | 24.5 | 11.1 | 17.7 | 12.1 | 1.1 | 0.5 | _ | _ | _ | _ |
| SMF 85859 | NFN | G. azzurrae | Isalo, Sakamalio | EF222305 | PT | M | 42.7 | 16.4 | 14.3 | 5.4 | 3.7 | 2.6 | 3.9 | 3.5 | 13.4 | 41.1 | 21.0 | 29.9 | 20.0 | 1.9 | 1.1 | 7.0 | 3.0 | 42 | 0.6 |
| SMF 85860 | NFN | G. azzurrae | Isalo, Sakamalio | EF222303 | PT | M | 43.7 | 16.4 | 13.5 | 5.7 | 4.0 | 2.5 | 3.8 | 4.0 | 12.2 | 42.3 | 21.1 | 27.7 | 20.0 | 1.9 | 1.0 | 7.5 | 2.7 | 42 | 0.5 |
| MRSN A3415 | NFN | G. pseudoasper | Nosy Be | - | - | M | 33.3 | 12.1 | 13 | 4.9 | 3.8 | 2.6 | 2.8 | 3.1 | 10.4 | 52.4 | 15.7 | 23 | 17.5 | 2.1 | 1.1 | 6.1 | 2.5 | 43 | 0.3 |
| MRSN A3416 | FN 6696 | G. pseudoasper | Nosy Be | - | - | M | 37.4 | 12.9 | 15 | 4.9 | 4.4 | 3 | 3.8 | 4.4 | 10.3 | 54.2 | 16.4 | 25 | 15.8 | 2.4 | 1.1 | 7.2 | 2.9 | 39 | 0.4 |
| MRSN A3417 | FN 6423 | G. pseudoasper | Nosy Be | - | - | F | 33.1 | 11.9 | 14 | 5.3 | 3.7 | 2.7 | 2.9 | 3 | 9.9 | 56.5 | 16.5 | 24 | 18.2 | 1.9 | 1 | - | - | - | - |
Subadult male in mediocre state of preservation,
with the belly opened for gonadal inspection and part of the ventral
surface of thighs cut and opened to check the glands. SVL 34.8 mm; for
other measurements see Tab. 1.
Body slender; head longer than wide, in line with the body; snout
slightly pointed in dorsal view, rather rounded in lateral view;
nostrils directed laterally, much nearer to tip of snout than to eye;
canthus rostralis well defined; tympanum distinct, rounded, its
horizontal diameter about 50% of eye diameter; supratympanic fold well
distinct, regularly curved; tongue distinctly bifid posteriorly. Arms
slender; subarticular tubercles single; outer and inner metacarpal
tubercles paired; fingers without webbing; finger disks triangular
distinctly enlarged; nuptial pads absent. Hind limbs slender;
tibiotarsal articulation reaching the nostril when hindlimbs are
adpressed along body; lateral metatarsalia partly connected; inner
metatarsal tubercle distinct, outer metatarsal tubercle small but
recognizable; webbing of foot 1(1), 2i(1), 2e(1), 3i(2), 3e(1),
4i(2), 4e(2), 5(1). Skin slightly granular on dorsum and belly,
ventral skin smooth on throat and chest. Femoral glands cluster ("Type
2", according to
We based the current description of variability
upon some specimens (paratypes and complementary individuals), part of
which (ZSM 23/2006, 37/2006, 107/2006, MRSN A5486 and MRSN A5483)
were also photographed in nature, and thus provided more diagnostic
characters. The female ZSM 23/2006 (Fig. 2; C–E),
shows a back with sparse larger warts. Its colouration appears light
brown with greyish shadings, darker dots and transversal bands on the
back and legs. These are more evident in the preserved individual,
where a pattern of darker spots is visible on the back, suggesting the
presence of a darker X-shaped drawing. These spots are visible in two
other individuals, MRSN A5484 (a female) and in the holotype MRSN A5487
(Fig. 2; A, K),
although for the former specimen we do not have photographs taken in
life. The tympanum is uniformly brownish, and the iris is yellowish
with darker reticulations. The belly is comparatively smooth, with
fewer warts on its lateral parts. The throat is quite smooth. The
central part of the belly is lighter than the flanks and the ventral
sides of thighs, whitish on breast and thorax, with sparse darker
spots. The inguinal part appears yellowish. The throat is darker than
the belly, with a median lighter (although not so contrasted) line. The
lateral borders of the lower jaw bear darker spots. After preservation,
the colouration appears substantially similar, although faded. The
juvenile ZSM 37/2006 (Fig. 2; E)
presents a rather smooth back and flanks with sparse and barely evident
warts. The colouration is brownish shading to the grey on the flanks
and lateral parts of the back, with darker spots, extending around the
flanks. The central part of the back is crossed by a longitudinal light
(almost beige) band which enlarges on the head to cover the upper
eyelids. The posterior part of such a band narrows to shade almost
totally at the level of the vent. A thin, almost continuous whitish
longitudinal line runs from the tip of the snout until the groin. The
juvenile ZSM 107/2006 (Fig. 2; I)
also shows a rather smooth back. The colouration is much darker, and
the markings and spots are less visible. The tympanum is lighter than
the surrounding areas, and the upper ridge is entoured by black
pigment. Both these juveniles after about four years of preservation
present a similar pattern of colouration as in life. In ZSM 107/2006 the
central part of the back appears quite lighter than the surrounding
areas, with a sort of arrow pattern. An interesting comparison is with
the only mature available male (SVL 36.6 mm) photographed in life, the
individual labelled UADBA 39099 (Fig. 2; F–H).
This male appears quite slim in the photographs (either in dorsal or
ventral view), with rather uniform light brown shading to greenish in
life, and a moderately glandular skin texture (Fig. 2; F–G). The belly appears rather smooth in life, with the whole venter and thorax whitish (Fig. 2; H).
The throat is darker with a rather indistinct central whitish band and
vocal sacs are not recognizable. Lower parts of arms and thighs are
pinkish, while tibiae are more whitish pigmented. The plantar surfaces
are also reddish-pink. In this male, the glands are well visible and
yellowish, and appear similar to those observed in Gephyromantis azzurrae, Gephyromantis corvus and Gephyromantis pseudoasper. In particular, they clearly belong to the gland “Type 2”, sensu
According to our observations, the species
lives in habitats that retain some humidity, such as rock cavities and
along the walls of the canyon-like formations. One important notation
comes from the fact that several of the collectors, independently (JER,
FG, JCR) found this species within the caves which are typical of the
area. We suspect that the species uses caves because these sites
presumably have a higher humidity than the surrounding areas. In such a
sense it behaves similarly to Gephyromantis corvus at Isalo, which is known to frequent narrow canyons and cave-like canyons (
Only known from the localities of the type specimens within the Tsingy de Bemaraha National Park.
Gephyromantis atsingy sp. n. differs from Gephyromantis pseudoasper, Gephyromantis azzurrae and Gephyromantis corvus by the lack of paired blackish skin folds (vocal sacs) along the lower jaws in adult males, and from Gephyromantis azzurrae also by details of colouration (see below). Following our measurements, adult males of Gephyromantis atsingy can also be differentiated among each other by the number of granules in the femoral glands: 70 granules in Gephyromantis atsingy; 96 granules in Gephyromantis corvus; 38–45 granules in Gephyromantis azzurrae and 39–43 granules in Gephyromantis pseudoasper. In addition, the new species differs from all three species by substantial genetic differentiation (see below).
All the described species of Gephyromantis, subgenus Phylacomantis, show similarities with Gephyromantis atsingy (Tab. 2).
The dorsal pattern is similar in all species, showing an assemblage of
darker spots and reticulations on the lighter background, and barred
legs and arms. The dorsal colouration in Gephyromantis atsingy is usually light brown-beige, with a somehow greenish shading, while in Gephyromantis corvus
it is uniformly grey or dark grey with sparse darker (uniformly-sized)
warts and dots. Notwithstanding, the examined specimens of Gephyromantis atsingy have a much more contrasted X-shaped dark spot on the back. This is less evident in Gephyromantis corvus,
where the dark-light pattern is more confuse and irregular. We
observed a longitudinal repetition of lighter elements, a longitudinal
light band or a middorsal light line only in Gephyromantis atsingy. The belly in both species is light, but in Gephyromantis atsingy
we detected more frequently the darker drawing with a lighter central
area on the throat and chest. According to the original description and
subsequent papers (
Distribution, habitats and diagnostic characters of the nominal species in the genus Gephyromantis, subgenus Phylacomantis.
| Species | Distribution | Habitat | SVL | Vocal Sacs | Dorsal colouration | Ventral colouration | Dorsal texture |
|---|---|---|---|---|---|---|---|
| Gephyromantis atsingy | Tsingy de Bemaraha | Karst pinnacles and caves | 35–43 mm | Non-evident | Light brownish with greenish shadings | Whitish | Slightly warty |
| Gephyromantis azzurrae | Isalo Massif | Open canyons and permanent rivers | 23–44 mm | Double and brownish | Brownish, sometimes with wide light band | Whitish, with reddish shadings | Warty with heterogeneous warts |
| Gephyromantis corvus | Isalo Massif | Close canyons and cave-like canyons | 39–41 mm | Double and blackish | Greyish with darker spots | Whitish | Warty |
| Gephyromantis pseudoasper | Sambirano, N, NW and NE | Rainforests, karstic areas | 33–37 mm | Double and blackish | Brownish | Whitish | Slightly warty |
The molecular data confirm the attribution of Gephyromantis atsingy to the subgenus Phylacomantis (
This species appears to be restricted to the Bemaraha Plateau, where it has been found in seven localities within the Tsingy de Bemaraha National Park. It may also occur in the Réserve Naturelle Intégrale, which forms the northerly limit of the Bemaraha Plateau, but survey data are lacking from this site. Within the national park, some areas of forest are damaged by conversion to agriculture and charcoal production, but the humid canyons where G. atsingy occur are generally well protected. We therefore recommend assigning a category of Near Threatened because the species nearly qualifies for listing as Vulnerable under D2: the species is confined to a single site, the Bemaraha Massif (1, 577 km2), with a plausible threat that could impact the species in the near future. If the threat became operational, the species would be eligible for listing as Endangered since its extent of occurrence is well within the 5, 000km2 threshold under the B criterion and it would occur at a single location (where the threat is habitat loss from agricultural activities and charcoal production) and there would be a continuing decline in the quality and area of habitat, qualifying the species for the criteria B1ab(iii).
The Bemaraha plateau is one of the most peculiar areas of western Madagascar in terms of amphibian richness and endemicity (
One powerful tool is the application of an integrated
taxonomy approach, which includes aspects of direct field surveys,
behavioural assessment, molecular screening and bioacustic analysis.
This is what allowed
As already stressed by
We expect that other forest blocks in western Madagascar may host further undescribed species of Gephyromantis and we highlight the need of conservation actions in Madagascar’s dry forests due to the increasing deforestation rate and changing climatic scenarios.
The accelerated species discovery in Malagasy amphibians
points to the importance of taxonomic surveys (see www.sahonagasy.org),
and we like to consider Gephyromantis atsingy as another “forceps delivered” species, according to the attractive definition given by
Many friends and colleagues helped us providing samples that have been used in this study. Thanks to S. Federici and M. Barbuto for their help in the lab. Special thanks to M. Vences, P. Bora, J. Köhler and H. Enting for the help in the field and for providing information on the herpetofauna of Bemaraha. We thank N. Kaffenberger for sharing unpublished information on molecular relationships, and A. Angulo and M. Hoffmann for their precious help on analysing the conservation status of this newly described species. This work has been carried out in the framework of collaboration agreements of the authors’ institutions with UADBA. The Malagasy authorities kindly granted research and export permits. A. Crottini was supported by a Dote Ricercatore of Regione Lombardia fellowship. Fieldwork by RKBJ and JCR was funded by the Darwin Initiative.