





(C) 2011 Natalia Pereira Benaim. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
As a secondary result of oil prospecting in Brazil, samples from the Campos Basin continental slope became available. In these samples, specimens of the genera Microgloma Sanders Allen, 1973 and Pristigloma Dall, 1900 were found. This contribution provides the southernmost record of the genus Microgloma, the first record of Microgloma mirmidina (Dautzenberg & Fischer 1897) from the western Atlantic, the descriptions of Microgloma macaron sp. n. and Microgloma nhanduti sp. n. as new species, and the shallowest record of Pristigloma alba Sanders & Allen 1973.
Pristiglomidae, Nuculanidae, Pelecypoda, deep-sea, biodiversity, Campos Basin slope, Rio de Janeiro
Knowledge of the deep-sea mollusks from the Campos Basin has increased in the last ten years, and several new species have been described (
Microgloma and Pristigloma comprise a few species that appear represented only in the Atlantic Ocean, each genus being represented by no more than five species. Their systematic affinities have been the subject of debate for almost 30 years (
The five species of Microgloma described until now [Microgloma tumidula (Monterossato, 1880), Microgloma mirmidina (Dautzenberg & Fischer, 1897), Microgloma yongei Sanders & Allen, 1973, Microgloma pusilla Jeffreys, 1979, and Microgloma guilornadi Hoeksema, 1993] were recorded from Western Europe (Iberian Peninsula, Mediterranean Sea, Azores and Canaries), West Africa (Cape Verde and Angola), Surinam, and North America. Prior to the report of
Pristigloma is a genus widespread in the entire Atlantic Ocean and is characterized by smooth, fragile shells, lamellar hinge teeth with an unequal number of teeth on both plates, and a large, internally elongated ligament which is opisthodetic (
Here, we present the first records of the genus Microgloma from the southwestern Atlantic, as well as descriptions of two new species belonging to this genus. We also present new points in the geographical distribution of Pristigloma alba Sanders & Allen 1973.
Material and methodsThe samples used in the present study were collected by means of a box corer in the Campos Basin, off Rio de Janeiro State (22°S, 41°W), Brazil, from the research vessel Astro–Garoupa, as part of the programs “Environmental Characterization of Campos Basin, RJ, Brazil" during the years 2002 and 2003, and “Habitats Project – Campos Basin Environmental Heterogeneity" in 2008 and 2009. Both programs were sponsored by the Brazilian oil company Petrobras S.A. Of the material obtained we observed 260 samples taken between the isobaths of 400 and 2500 m. The list of localities with Microgloma and/or Pristigloma specimens is given in Tables 1 and 2. Most of the shells were found in a good state of preservation, with valves attached, ligaments intact, and often with the mass of the animal body inside the shell. Unfortunately, there were no preserved organs in these cases. Each specimen was examined under stereoscopic microscope (Nikon SMZ 800), and selected specimens were photographed with a scanning electron microscope (ZEISS EVO 40), at the Gerência de Bioestratigrafia e Paleoecologia Aplicada (BPA), of the Petrobras Research Center (Centro de Pesquisas da Petrobras, CENPES).
Table of the localities sampled as part of the project Environmental Characterization of Campos Basin.
Table of the localities sampled as part of the Habitats Project – Campos Basin Environmental Heterogeneity.
| Cruise | Station (#) | Depth (m) | Latitude / Longitude | Date | |
|---|---|---|---|---|---|
| HAB 4 | D11 | R1 | 2449 | 22°52'15.30"S, 40°05'10.40"W | 22/5/2008 |
| HAB 4 | G12 | R1 | 3236 | 22°12'19.50"S, 38°35'52.00"W | 25/5/2008 |
| HAB 6 | D07 | R1 | 698 | 22°36'27.10"S, 40°22'29.60"W | 25/6/2008 |
| HAB 6 | D07 | R2 | 700 | 22°36'27.30"S, 40°22'29.30"W | 25/6/2008 |
| HAB 6 | D07 | R2 | 700 | 22°36'27.30"S, 40°22'29.30"W | 25/6/2008 |
| HAB 6 | A7 | R1 | 694 | 23°39'20.10"S, 41°18'30.30"W | 23/6/2008 |
| HAB 6 | A7 | R2 | 692 | 23°39'19.80"S, 41°18'30.20"W | 23/6/2008 |
| HAB 6 | A7 | R2 | 692 | 23°39'19.80"S, 41°18'30.20"W | 23/6/2008 |
| HAB 6 | A7 | R3 | 733 | 23°39'19.90"S, 41°18'30.50"W | 24/6/2008 |
| HAB 6 | CANAC7 | R1 | 758 | 21°47'26.70"S, 40°02'13.30"W | 28/6/2008 |
| HAB 6 | CANAC7 | R2 | 753 | 21°47'26.60"S, 40°02'13.70"W | 28/6/2008 |
| HAB 6 | I07 | R1 | 694 | 21°11'12.20"S, 40°12'52.00"W | 29/6/2008 |
| HAB 6 | D07 | R2 | 700 | 22°36'27.30"S, 40°22'29.00"W | 25/6/2008 |
| HAB 6 | D07 | R2 | 700 | 22°36'27.30"S, 40°22'29.30"W | 25/6/2008 |
| HAB 6 | D07 | R1 | 698 | 22°36'27.10"S, 40°22'29.60"W | 25/6/2008 |
| HAB 6 | D07 | R2 | 700 | 22°36'27.30"S, 40°22'29.30"W | 25/6/2008 |
| HAB 7 | D06 | R1 | 396 | 22°33'35.70"S, 40°26'38.90"W | 08/7/2008 |
| HAB 7 | D06 | R3 | 393 | 22°33'33.80"S, 40°26'40.30"W | 11/7/2008 |
| HAB 7 | H7 | R1 | 700 | 21°41'12.30"S, 40°02'20.20"W | 07/7/2008 |
| HAB 7 | H7 | R2 | 699 | 21°41'11.70"S, 40°02'20.70"W | 07/7/2008 |
| HAB 7 | H7 | R3 | 700 | 21°41'11.80"S, 40°02'20.40"W | 07/7/2008 |
| HAB 7 | I07 | R3 | 792 | 21°11'02.60"S, 40°12'18.20"W | 05/7/2008 |
| HAB 8 | D06 | R2 | 401 | 22°33'35.10"S, 40°26'37.50"W | 31/1/2009 |
| HAB 8 | D07 | R2 | 696 | 22°36'25.30"S, 40°22'30.60"W | 29/1/2009 |
| HAB 8 | C10 | R3 | 1953 | 23°08'23.80"S, 40°36'37.90"W | 27/1/2009 |
| HAB 8 | A07 | R2 | 701 | 23°39'20.60"S, 41°18'28.20"W | 28/1/2009 |
| HAB 8 | A07 | R3 | 693 | 23°39'21.90"S, 41°18'33.10"W | 28/1/2009 |
| HAB 9 | CANAC7 | R2 | 780 | 21°47'26.60"S, 40°01'55.30"W | 06/2/2009 |
| HAB 9 | CANAC7 | R3 | 775 | 21°47'26.70"S, 40°01'55.50"W | 06/2/2009 |
| HAB 9 | CANG–7 | R2 | 720 | 21°56'11.90"S, 39°57'45.30"W | 07/2/2009 |
| HAB 9 | H07 | R2 | 702 | 21°41'12.60"S, 40°01'56.10"W | 06/2/2009 |
Taxonomic identifications were made through comparison with the figures of type specimens [Microgloma pusilla (Jeffreys, 1879)] and descriptions available in the literature (
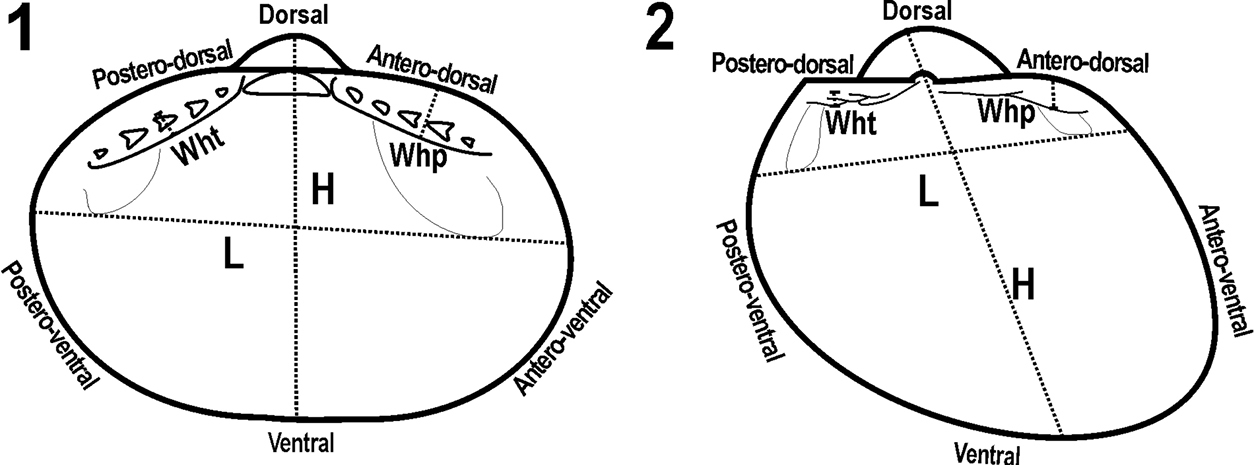
Scheme of the measurements and abbreviations used in the descriptions of the species A Microgloma B Pristigloma H height L length, wht – width of hinge teeth, whp – width of hinge plate.
The material analyzed in this study is deposited in the Mollusca collections of the following institutions: Departamento de Zoologia, Instituto de Biologia, Universidade Federal do Rio de Janeiro (IBUFRJ); Museu Nacional do Rio de Janeiro (MNRJ); Museu de Zoologia da Universidade de São Paulo (MZUSP); Museu Oceanográfico da Fundação Universitária de Rio Grande (MOFURG); National Museum of Natural History, Smithsonian Institute (USNM); and Muséum National d’Histoire Naturelle, Paris (MNHN). The following abbreviations are used: # – station; IBUFRJ – Instituto de Biologia da Universidade Federal do Rio de Janeiro; MNRJ – Museu Nacional do Rio de Janeiro; MZUSP – Museu de Zoologia da Universidade de São Paulo; MOFURG – Museu Oceanográfico da Fundação Universitária de Rio Grande; MNHN – Muséum National d’Histoire Naturelle, Paris; USNM – National Museum of Natural History, Smithsonian Institute; MCZ – Museum of Comparative Zoology, Harvard University, Cambridge, U.S.A.
Systematicshttp://species-id.net/wiki/Pristigloma_alba
Figs 3–8MCZ 271976. We tried to find this lot in the MCZ with the help of Mr. Cleo Oliveira, but the curators could not find it. We made contact with Dr John A. Allen who sent us live specimens from Surinam Basin #293 (08°58'N54°04'W, 1518 m) to compare with Campos Basin specimens. Furthermore, using the good description in
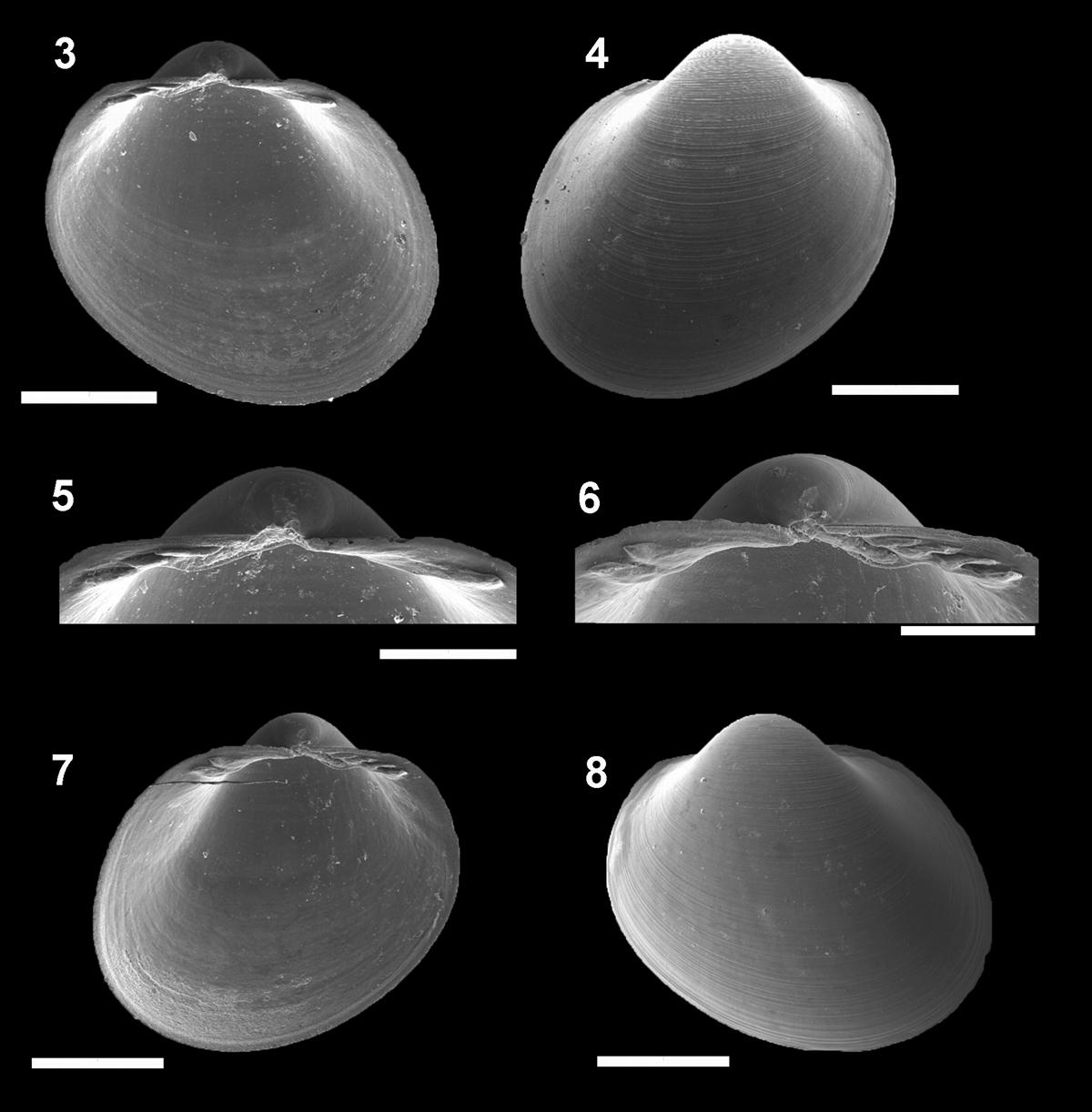
Pristigloma alba Sanders & Allen, 1973 IBUFRJ 16161. Left valve: internal view 3 external view 4 detail of the hinge plate 5 Right valve: detail of the hinge plate 6 internal view 7 external view 8 Scale bars 3–4, 7–8 = 500 μm, 5–6= 250 μm
Angola, 3739–4597 m; Canaries, 6709–6711 m; North America, 2178–4892 m; Brazil, 3459 m; Argentina, 4382–4405 m (all from
MNRJ 19114 (# 68, 2003), 4 valves; MZUSP 99977 (# 68, 2003), 4 valves; IBUFRJ 16161 (# 68, 2002), 3 valves; IBUFRJ 19001 (# 10, 2002), 5 valves; IBUFRJ 19002 (# 42, 2002), 3 valves; IBUFRJ 19003 (# 47, 2002), 1 valves; IBUFRJ 19004 (# 48, 2002), 7 valves; IBUFRJ 19005 (# 52, 2002), 1 valve; IBUFRJ 19006 (# 58, 2002), 6 valves; IBUFRJ 19007 (# 62, 2002), 3 valves; IBUFRJ 19008 (# 63, 2002), 6 valves; IBUFRJ 19009 (# 73, 2002), 14 valves; IBUFRJ 19010 (# 77, 2002), 2 valves; IBUFRJ 19011 (# 78, 2002), 4 valves; IBUFRJ 19012 (# 83, 2002), 2 valves; IBUFRJ 19013 (# 87, 2002), 6 valves; IBUFRJ 19014 (# 46, 2003), 1 valve; IBUFRJ 19015 (# 48, 2003), 15 valves; IBUFRJ 19016 (# 52, 2003), 1 valve; IBUFRJ 19017 (# 53, 2003), 16 valves; IBUFRJ 19018 (# 58, 2003), 3 valves; IBUFRJ 19019 (# 61, 2003), 2 valves; IBUFRJ 19020 (# 63, 2003), 16 valves; IBUFRJ 19021 (# 68, 2003), 14 valves; IBUFRJ 19022 (# 72, 2003), 1 valve; IBUFRJ 19023 (# 73, 2003), 8 valves; IBUFRJ 19024 (# 78, 2003), 14 valves; IBUFRJ 19025 (# 82, 2003), 1 valve; IBUFRJ 19026 (# 86, 2003), 4 valves.
Shell H/L ratio about 1.04. W/H ratio approximately 0.86. Muscle scars rarely visible; anterior adductor scar 2/3 of posterior scar. Anterior and posterior parts of the hinge plate usually of same length. Width of posterior row of teeth occupies about 65% of width of hinge plate, which is thin for its size (whp/H approximately 0.07). Posterior part of the hinge plate longer than anterior one. Prodissoconch smooth, with length approximately 120 µm.
This species was recently well described and for this reason we add only new information on the proportions of the shell (H/L and width) and hinge plate characteristics.
http://species-id.net/wiki/Microgloma_mirmidina
Figs 9–13Southeast of Flores, Azores, 1846 m (
MNRJ 19115 (#68, 2003), 6 valves; MZSP 99978 (#71, 2003), 7 valves; IBUFRJ 15889 (#67, 2003), 5 valves; IBUFRJ 17501 (#87, 2003), 4 valves; IBUFRJ 19084 (# 10, 2001), 4 valves; IBUFRJ 19085 (# 28, 2001), 1 valve; IBUFRJ 19086 (# 42, 2002), 1 valve; IBUFRJ 19087 (# 48, 2002), 6 valves and 2 specimens; IBUFRJ 19088 (# 51, 2002), 1 valve; IBUFRJ 19089 (# 53, 2002), 6 valves and 1 specimen; IBUFRJ 19090 (# 57, 2002), 1 valve IBUFRJ 19091 (# 62, 2002), 4 valves; IBUFRJ 19092 (# 63, 2002), 10 valves; IBUFRJ 19093 (# 68, 2002), 1 valve; IBUFRJ 19094 (# 73, 2002), 6 valves and 1 specimen; IBUFRJ 19095 (# 75, 2002), 1 valve; IBUFRJ 19096 (# 77, 2002), 12 valves and 1 specimen; IBUFRJ 19097 (# 78, 2002), 4 valves; IBUFRJ 19098 (# 81, 2002), 2 valves; IBUFRJ 19098 (# 81, 2002), 2 valves; IBUFRJ 19099 (# 83, 2002), 4 valves and 1 specimen; IBUFRJ 19100 (# 85, 2002), 1 valve; IBUFRJ 19101 (# 86, 2002), 6 valves; IBUFRJ 19102 (# 87, 2002), 5 valves; IBUFRJ 19103 (# 50A, 2003), 1 valve; IBUFRJ 19105 (# 61, 2003), 3 valves; IBUFRJ 19106 (# 63, 2003), 2 valves; IBUFRJ 19107 (# 72, 2003), 2 valves and 1 specimen; IBUFRJ 19108 (# 73, 2003), 4 valves; IBUFRJ 19109 (# 77, 2003), 5 valves and 3 specimens; IBUFRJ 19110 (# 78, 2003), 4 valves; IBUFRJ 19111 (# 82, 2003), 2 valves; IBUFRJ 19112 (# 84, 2003), 2 valves; IBUFRJ 19113 (# 86, 2003), 2 valves; IBUFRJ 19114 (# C10, 2008), 1 valve.
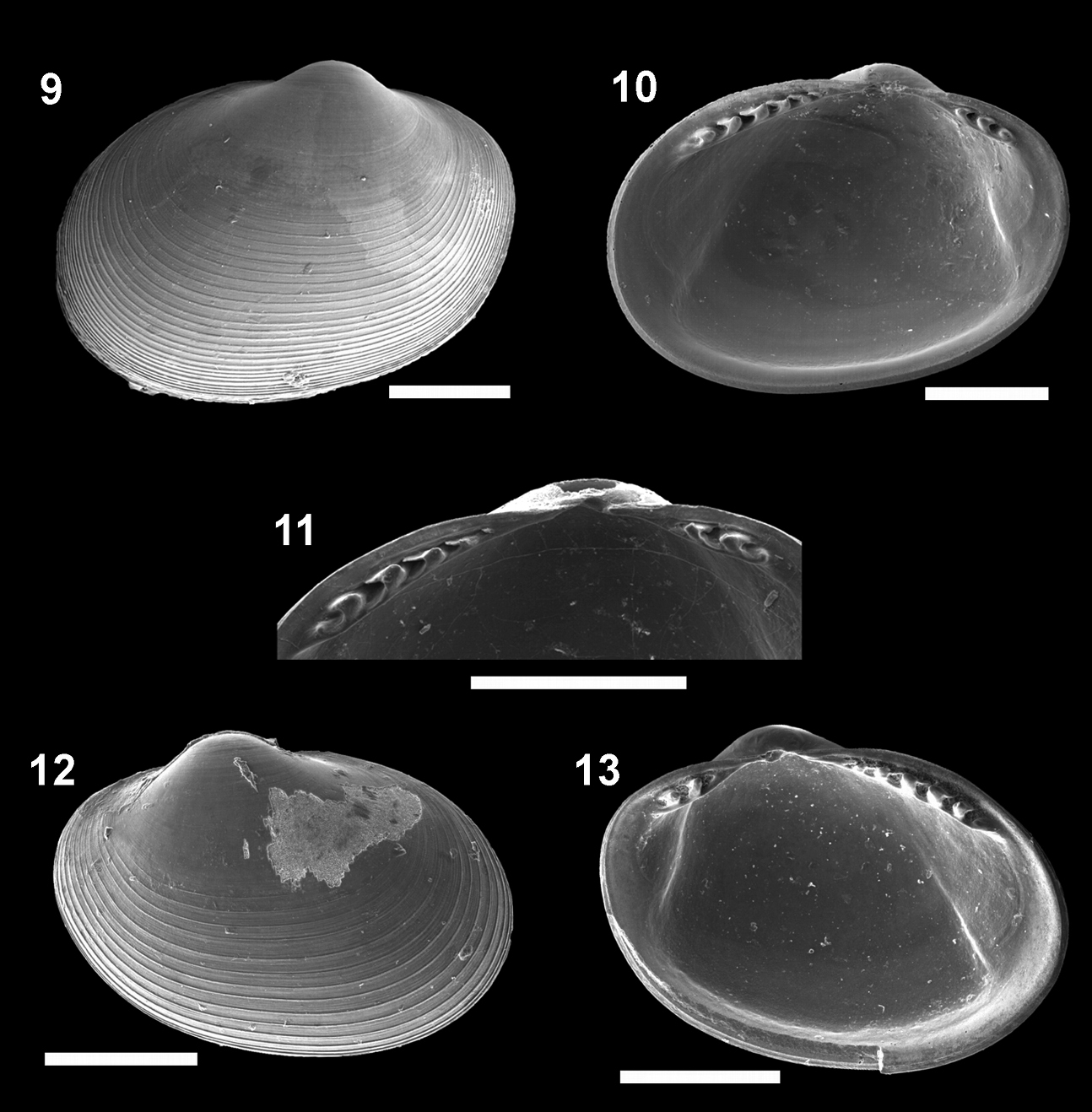
Figures 9–13. Microgloma mirmidina (Dautzenberg & Fischer, 1897). External view, left valve 9 internal view, right valve 10 Detail of the hinge plate 11 External view, right valve 12 internal view, left valve 13 All from the lot IBUFRJ 15889. Scale bars A= 200 μm; B, E = 300 μm; C = 100 μm; D = 250μm.
Shell H/L approximately 0.79 (n 10, min 0.75, max 0.83), W/H approx. 0.59(n 10, min 0.53, max 0.63). Posterior part of the hinge plate shorter than anterior one. Width of both the anterior and posterior rows of teeth occupies nearly half of the width of the hinge plate, which is moderately thick for its size (whp/H about 0.11). Prodissoconch smooth, length approximately 176 µm.
The radial sculpture on the prodissoconch surface is absent in Microgloma mirmidina, but admittedly it is not always clearly developed in the other species of Microgloma (
urn:lsid:zoobank.org:act:31275EB9-546D-414B-82B0-C59CA55F8C93
http://species-id.net/wiki/Microgloma_macaron
Figs 14– 22MNRJ 19.112 (Figs 14, 16, 18).
Campos Basin, #54, 12/12/2002, 750m, 21°57'17, 5"S, 39°56' 01, 1"W.
Paratypes. IBUFRJ 15297, 8 valves and 2 specimens; MZUSP 99979 , 4 valves; USNM 1156943 , 6 valves; MNHN 24596, 6 valves; all from the type locality.
The species epithet refers to the French macaroon cookie (“macaron" in the French language), which has a similar appearance to the articulated valves. The species epithet is proposed as a noun in apposition.
IBUFRJ 19145 (# 32, 2002), 3 valves; IBUFRJ 19146 (# 33, 2002), 1 valve; IBUFRJ 19147 (# 34, 2002), 1 valve; IBUFRJ 19148 (# 36, 2002), 1 valve; IBUFRJ 15482 (# 54, 2002), 6 valves; IBUFRJ 17033 (# 59, 2002), 1 specimen; IBUFRJ 19150 (# 61, 2002), 3 specimens; IBUFRJ 16074 (# 64, 2002), 14 valves and 1 specimen; IBUFRJ 15141 (# 69, 2002), 7 valves; IBUFRJ 15285 (# 74, 2002), 2 valves; IBUFRJ 15635 (# 54, 2003), 32 valves and 5 specimens; IBUFRJ 19152 (# 59, 2003), 6 valves and 1 specimen; IBUFRJ 19153 (# 61, 2003), 1 valve; IBUFRJ 19153 (# 61, 2003), 1 valve; IBUFRJ 19154 (# 64, 2003), 18 valves and 4 specimens; IBUFRJ 19155 (# 69, 2003), 4 valves; IBUFRJ 19156 (# D11, 2008), 1 specimen; IBUFRJ 19157 (# G12, 2008), 1 specimen; IBUFRJ 19158 (# A7, 2008), 10 specimens; IBUFRJ 19159 (# A7, 2008), 1 specimen; IBUFRJ 19160 (# A7, 2008), 2 valves and 5 specimens; IBUFRJ 19161 (# D7, 2008), 1 specimen; IBUFRJ 19162 (# D7, 2008), 6 specimens; IBUFRJ 19163 (# CANAC7, 2008), 1 specimen; IBUFRJ 19164 (# H7, 2008), 1 specimen; IBUFRJ 19165 (# H7, 2008), 1 specimen; IBUFRJ 19166 (# H7, 2008), 5 specimens; IBUFRJ 19167 (# I7, 2008), 2 specimens; IBUFRJ 19168 (# A7, 2009), 3 specimens; IBUFRJ 19169 (# A7, 2009), 5 specimens; IBUFRJ 19170 (# D6, 2009), 1 specimen; IBUFRJ 19171 (# D7, 2009), 1 specimen; IBUFRJ 19172 (# H7, 2009), 12 specimens; IBUFRJ 19173 (# CANAC7, 2009), 1 specimen; IBUFRJ 19174 (# CANAC7, 2009), 1 specimen; IBUFRJ 19175 (# CANG7, 2009), 3 specimens; IBUFRJ 19177 (# 64, 2003), 4 specimens.
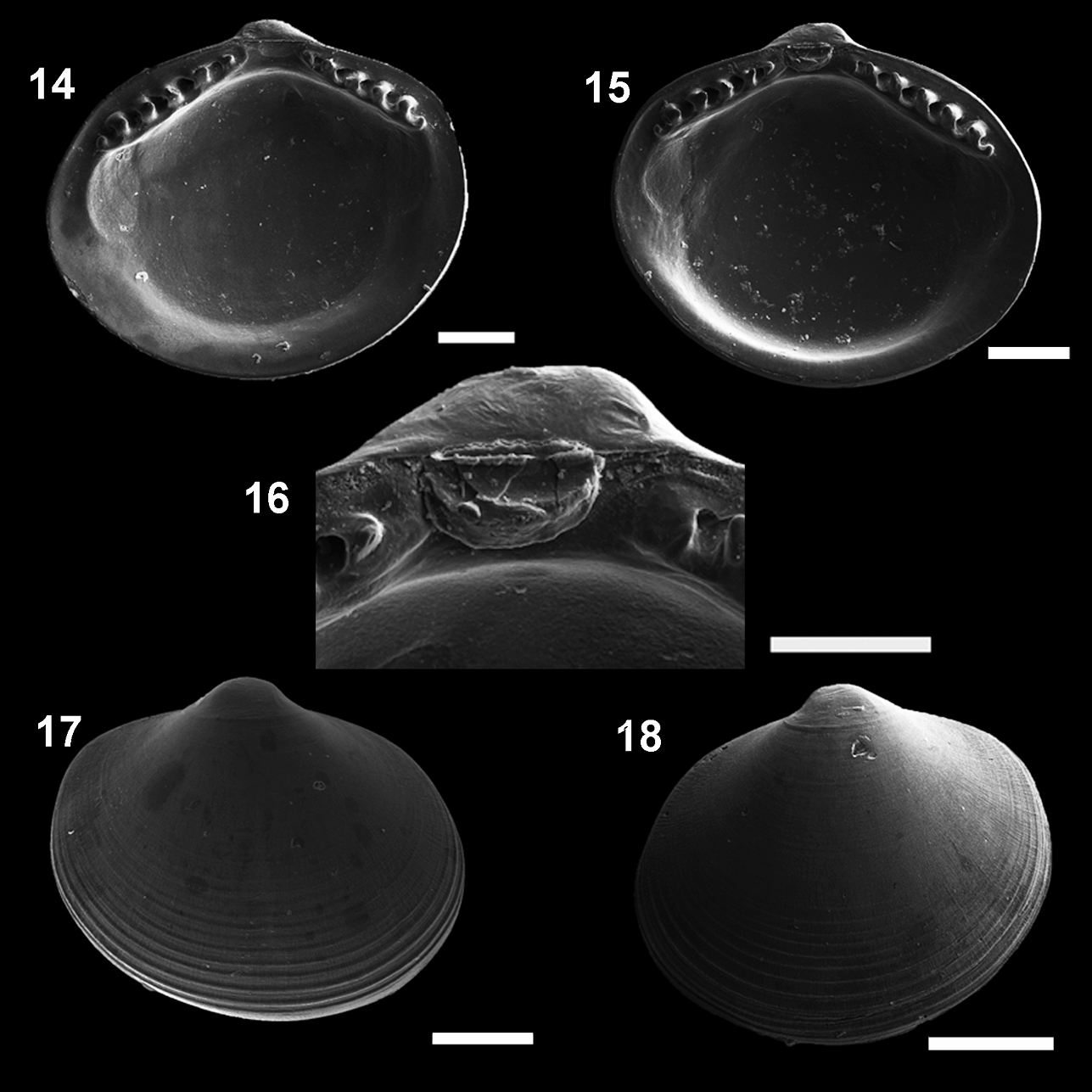
Microgloma macaron sp. n. Internal view, right valve 14 left valve 15 detail of the hinge plate and ligament 16 External view, right valve 17 left valve 18 Holotype MNRJ 19112 (14, 16, 18). Paratype IBUFRJ 15297 15, 17 Scale bars: 14, 16= 200 µm; 15, 18 = 300 µm; 17 = 250 µm.
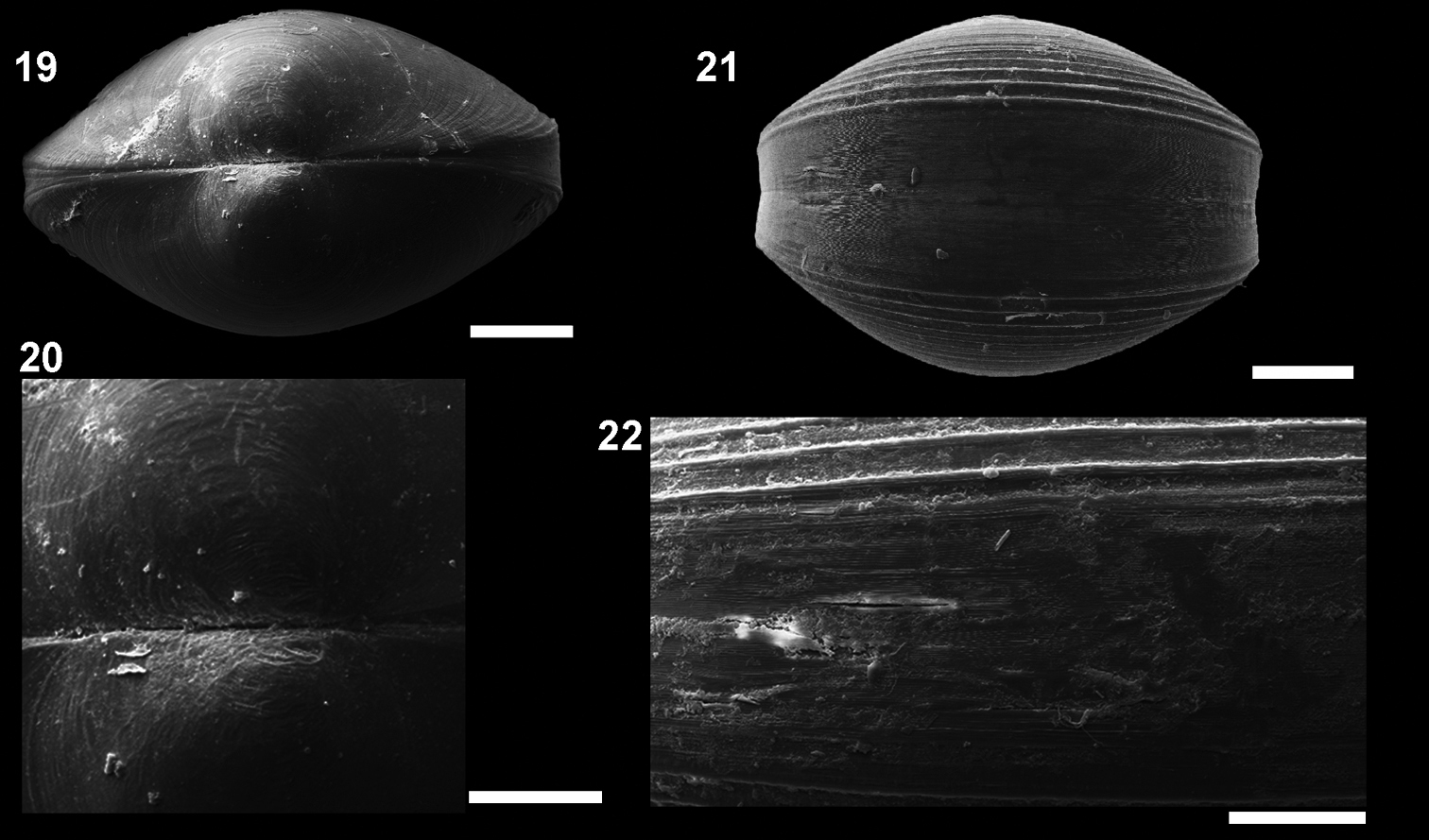
Microgloma macaron sp. n. Dorsal view 19 prodissoconch 20 (IBUFRJ 15297). Ventral margin view, extended margin 21 periostracum of the ventral margin 22 (IBUFRJ 19177). Scale bars: 19, 21 = 200 µm; 20, 22 = 100 µm.
Shell minute, ovate. Entire shell margin thickened and covered by an extension of the periostracum. Prodissoconch surface with one subtle radial striae.
Shell minute, ovate, H/L ratio about 0.81 (n 38, min 0.75, max 0.92), W/H ratio about 0.84 (n 14, min 0.77, max 0.87), glossy, translucent, robust for its size, equilateral; umbones prominent, large, posterior to midline, orthogyrous. Antero–dorsal margin straight, oblique; anterior margin rounded, extended. Antero–ventral margin, postero–ventral margin shorter and rising up to the short posterior end; posterior margin slightly truncated, forming a small shoulder. Entire shell margin thickened and covered by an extension of the periostracum, resembling a macaroon in ventral view (Figs 21–22). Surface with commarginal growth lines. Hinge plate with 5–7 anterior and 4–5 posterior teeth, interrupted by a large, rectangular and shallow resilifer. Width of both anterior and posterior row of teeth occupies about 70% of width of hinge plate, which is thick for its size (whp/H approximately Campos Basin, #54, 12/12/2002 0.14). Posterior part of the hinge plate shorter than anterior one. Prodissoconch surface nacreous, with one subtle radial striae (Fig 20), and length approximately 244 µm. Maximum adult shell length 1.20 mm.
Measurements of the type material. * Width = width of a single valve.
| Microgloma macaron sp. n. | |||
|---|---|---|---|
| Length | Height | Width* | |
| MNRJ 19.112 | 1.20 | 1.00 | 0.36 |
| IBUFRJ 15.297 | 1.11 | 0.87 | 0, 31 |
| IBUFRJ 15.297 | 1.11 | 0.89 | 0.36 |
| IBUFRJ 15.297 | 1.11 | 0.91 | 0.36 |
| IBUFRJ 15.297 | 1.11 | 0.84 | 0.33 |
| IBUFRJ 15.297 | 1.11 | 0.89 | 0.36 |
| IBUFRJ 15.297 | 1.11 | 0.89 | 0.36 |
| IBUFRJ 15.297 | 1.16 | 0.93 | 0.38 |
| IBUFRJ 15.297 | 1.11 | 0.91 | 0.33 |
| IBUFRJ 15.297 | 1.13 | 0.82 | 0.31 |
| IBUFRJ 15.297 | 1.13 | 0.89 | 0.33 |
| MZSP 99.979 | 1.11 | 0.87 | 0.33 |
| MZSP 99.979 | 1.13 | 1.04 | 0.36 |
| MZSP 99.979 | 1.16 | 0.93 | 0.38 |
| MZSP 99.979 | 1.11 | 0.91 | 0.31 |
| USNM 1156943 | 1.11 | 0.89 | 0.36 |
| USNM 1156943 | 1.11 | 0.89 | 0.36 |
| USNM 1156943 | 1.13 | 0.91 | 0.36 |
| USNM 1156943 | 1.13 | 0.89 | 0.38 |
| USNM 1156943 | 1.13 | 0.91 | 0.33 |
| USNM 1156943 | 1.11 | 0.89 | 0.36 |
| MNHN 24596 | 1.11 | 0.91 | 0.33 |
| MNHN 24596 | 1.18 | 0.93 | 0.38 |
| MNHN 24596 | 1.11 | 0.91 | 0.36 |
| MNHN 24596 | 1.11 | 0.89 | 0.38 |
| MNHN 24596 | 1.11 | 0.91 | 0.36 |
| MNHN 24596 | 1.04 | 0.87 | 0.38 |
In some other species of Microgloma, the shell is expanded ventrally, around the valve, which enlarges the internal volume of the animal and counterbalances the effects of miniaturization (
Compared to Microgloma pusilla and Microgloma mirmidina, Microgloma macaron is distinguished by the ovate outline, umbones at midline and much more projecting, and a thicker hinge plate. Microgloma macaron is similar to Microgloma yongei in outline, but compared with the paratypes figured by
Except for two, probably worn, valves found at twostations at approx. 3000 m depth, and one at 1970 m, this species is concentrated at depths between 400–750 m. Untill now this species is reccorded solely in Campos Basin.
urn:lsid:zoobank.org:act:511F7840-3490-4EA1-A48F-CB56C60329FC
http://species-id.net/wiki/Microgloma_nhanduti
Figs 23–31MNRJ 19.113 (Figs 26, 28).
Campos Basin, #54, 12/12/2002, 750m, 21°57'17.5"S, 39°56'01.1"W.
IBUFRJ 14991 (# 54, 2002), 2 valves and 1 specimen; IBUFRJ 19176 (# 64, 2003), 1 specimen; MZSP 99980 (# 54, 2002), 5 valves; USNM 1156944 (# 54, 2002), 5 valves; MNHN 24597 (# 54, 2002), 2 valves 1 specimen.
Nhanduti is a Tupi–Guarani term (the language spoken by the largest groups of native people living in Brazil prior to the European colonization) for a spider web–like structure, similar to those present on the prodissoconch of this species. The species epithet is proposed as a noun in apposition.
IBUFRJ 15140 (# 69, 2002), 17 valves and 2 specimens; IBUFRJ 15283 (# 69, 2002), 6 valves; IBUFRJ 19115 (# 33, 2002), 1 valve; IBUFRJ 19116 (# 64, 2002), 16 valves and 1 specimen; IBUFRJ 19117, (# 49, 2003), 7 valves; IBUFRJ 19118, (# 54, 2003), 1 valve; IBUFRJ 19119, (# 59, 2003), 10 valves and 2 specimens; IBUFRJ 19121, (# 34, 2002), 1 valve; IBUFRJ 19122, (# 37, 2002), 1 valve; IBUFRJ 19123, (# 38, 2002), 1 valve; IBUFRJ 19124, (# 64, 2003), 12 valves and 1 specimen; IBUFRJ 19125, (# 75, 2003), 2 valves; IBUFRJ 19126, (# A7, 2008), 4 specimens; IBUFRJ 19127, (# A7, 2008), 1 valve; IBUFRJ 19128, (# A7, 2008), 1 valve and 3 specimens; IBUFRJ 19129, (# D7, 2008), 1 specimen; IBUFRJ 19130 (# H7, 2008), 2 specimens; IBUFRJ 19131 (# D6, 2008), 3 specimens; IBUFRJ 19132 (# H7, 2008), 1 specimen; IBUFRJ 19133 (# H7, 2008), 4 valves; IBUFRJ 19134 (# D6, 2008), 1 specimen; IBUFRJ 19135 (# I7, 2008), 1 specimen; IBUFRJ 19136 (# A7, 2009), 2 specimens; IBUFRJ 19137 (# A7, 2009), 6 specimens; IBUFRJ 19138 (# D6, 2009), 4 specimens; IBUFRJ 19139 (# H7, 2009), 10 specimens; IBUFRJ 19140 (# CANAC7, 2009), 5 specimens and 2 valves; IBUFRJ 19141(# CANAC7, 2008), 1 specimen; IBUFRJ 19142 (# I7, 2008), 2 specimens; IBUFRJ 19143 (# CANAC7, 2008), 6 specimens; IBUFRJ 19144 (# D7, 2008), 1 specimen.
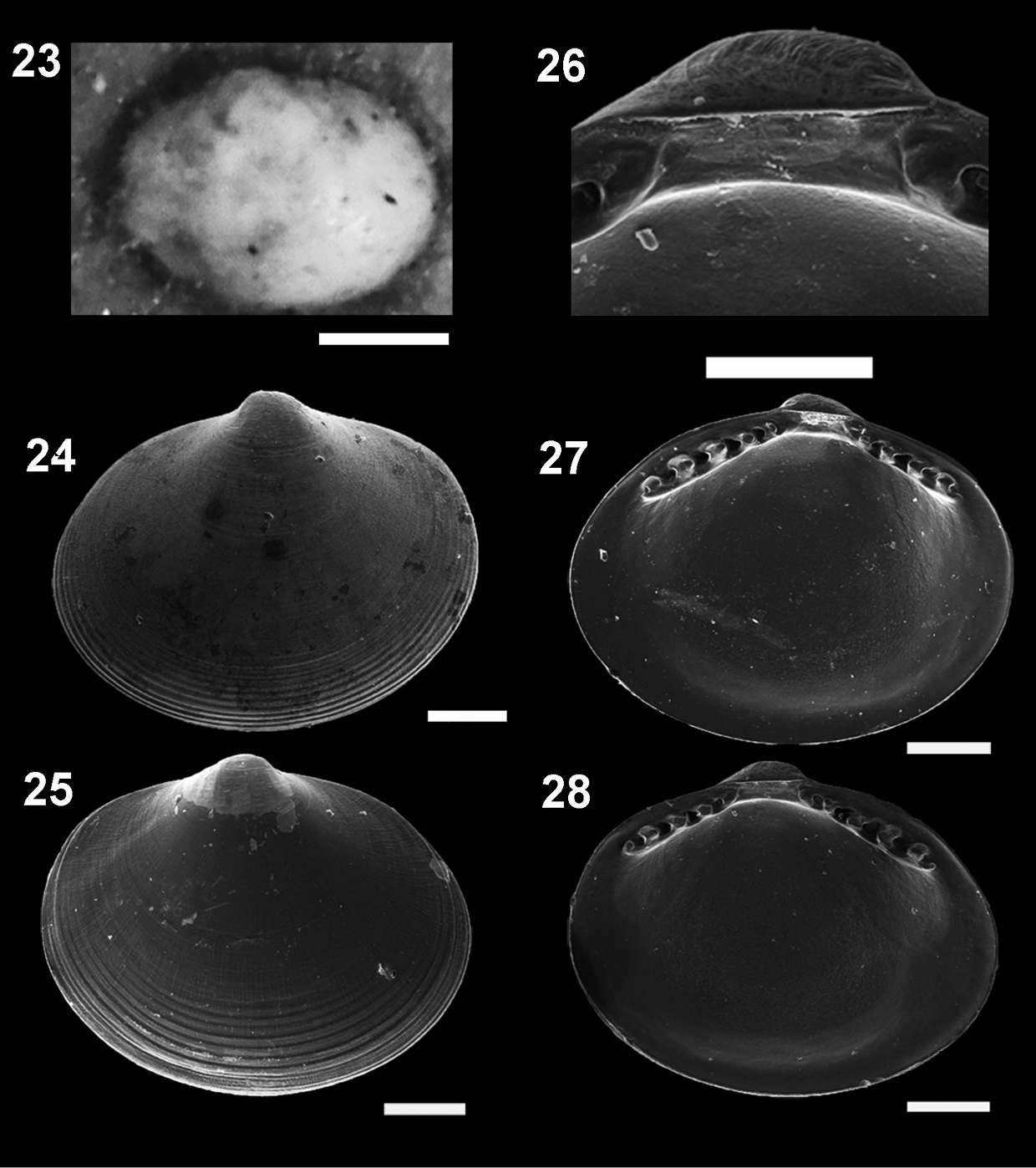
Figures 23–28. Microgloma pusilla (Jeffreys, 1879) USNM 199712 23 Microgloma nhanduti sp. n. External view, leftt valve 24 IBUFRJ 15283, right valve 25 IBUFRJ 14991. Internal view holotype MNRJ 19.113, detail of the reslifer and umbo 26 right valve 27 left valve 28 Scale bars: 23 = 500 µm; 26 = 100 µm; 24–25, 27–28 = 200 µm.
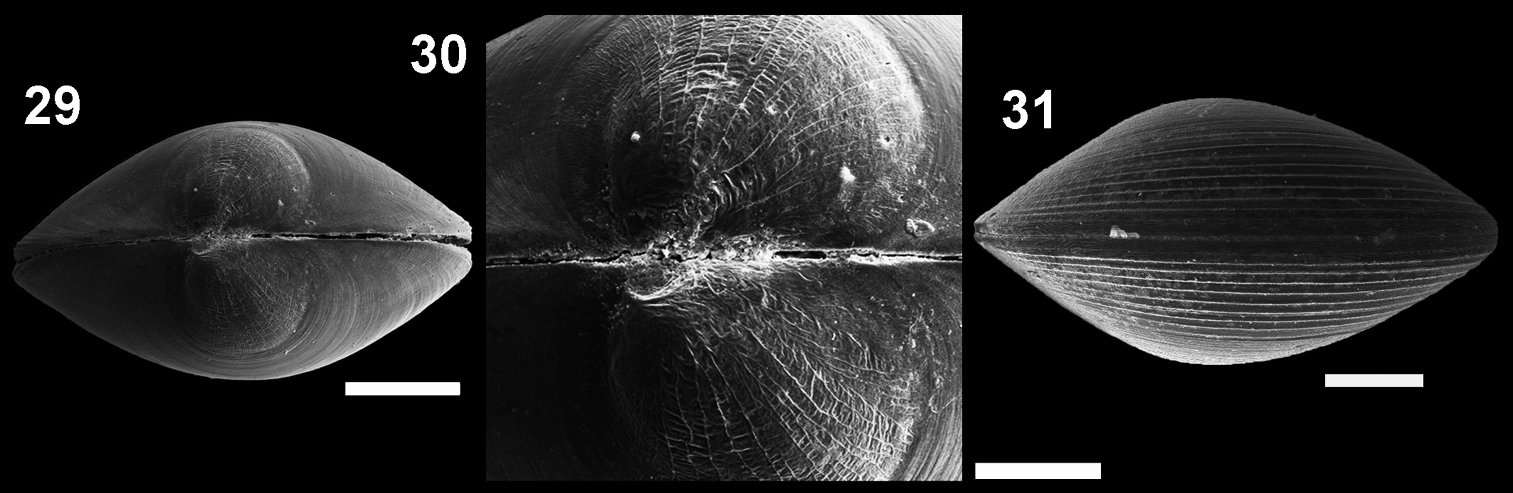
Microgloma nhanduti sp. n. Dorsal view IBUFRJ 19131 29 prodissoconch surface 30 ventral margin IBUFRJ 19176 31 Scale bars: 29, 30 = 200 µm, 31 = 80 µm.
Shell minute, ovate. Entire margin slightly thickened. Hinge plate moderately thick. Prodissoconch with web–like striae.
Shell minute, ovate, H/L ratio about 0.78 (n 29, min 0.74, max 0.81), Width of both valves/H ratio about 0.7 (n 11, min 0.67, max 0.77), inequilateral, translucent, glossy; umbones moderately prominent, large, posterior to midline, orthogyrous. Antero–dorsal margin convex and oblique; anterior margin rounded and projected. Antero–ventral margin convex, postero–ventral margin shorter and rising up to short and rounded posterior end; postero–dorsal margin convex, but forming a small shoulder. Entire shell margin slightly thickened. Surface with subtle commarginal growth lines. Hinge plate with 5–7 anterior and 4–5 posterior teeth, interrupted by a shallow rectangular hinge plate. Width of both anterior and posterior row of teeth occupies about 70% of width of hinge plate, which is thick for its size (whp/H approximately 0.14). Posterior part of the hinge plate shorter than anterior one. Prodissoconch surface nacreous, with several radial striae, resembling a spider’s web. Prodissoconch length approximately 246 µm.Maximum adult shell length 1.16 mm.
Measurements of the type material. * Width = width of a single valve.
| Microgloma nhanduti sp. n. | |||
|---|---|---|---|
| Length | Height | Width* | |
| MNRJ 19.113 | 1.12 | 0.90 | 0.29 |
| IBUFRJ 14.991 | 1.11 | 0.84 | 0.24 |
| IBUFRJ 14.991 | 1.04 | 0.82 | 0.29 |
| IBUFRJ 14.991 | 1.09 | 0.89 | 0.29 |
| IBUFRJ 19.176 | 1.13 | 0.89 | 0.29 |
| MZSP 99.980 | 1.09 | 0.82 | 0.29 |
| MZSP 99.980 | 1.09 | 0.82 | 0.27 |
| MZSP 99.980 | 0.98 | 0.76 | 0.24 |
| MZSP 99.980 | 1.09 | 0.82 | 0.27 |
| MZSP 99.980 | 1.11 | 0.82 | 0.27 |
| MZSP 99.980 | 1.11 | 0.84 | 0.22 |
| USNM 1156944 | 1.11 | 0.87 | 0.27 |
| USNM 1156944 | 1.11 | 0.87 | 0.29 |
| USNM 1156944 | 0.96 | 0.78 | 0.27 |
| USNM 1156944 | 1.07 | 0.80 | 0.24 |
| USNM 1156944 | 1.11 | 0.89 | 0.27 |
| MNHN 24596 | 1.07 | 0.87 | 0.22 |
| MNHN 24596 | 1.07 | 0.87 | 0.22 |
| MNHN 24596 | 1.09 | 0.80 | 0.22 |
Microgloma nhanduti sp. n. is similar to Microgloma pusilla and Microgloma tumidula in the oval outline. but has more prominent umbones (Figs 26–28). Compared to Microgloma macaron sp. n. and Microgloma yongei, Microgloma nhanduti sp. n. has a more elongated outline, a longer anterior area, and a more evident prodissoconch sculpture, with a web–like pattern (Figs 29–30). In Microgloma nhanduti sp. n. the umbo is not as prominent and the shell margin not as thick as in Microgloma macaron sp. n.Untill now this species is reccorded solely in Campos Basin.
The genus Microgloma is still in need of review since, as stated by
The sculpture pattern on the prodissoconch surface is not a character commonly used in taxonomy of the protobranchs and, considering the confused taxonomy of the protobranchs (as seen by the genus Microgloma), we believe it might be useful to better determine the genera. This character has been recorded recently in the literature, and some species, from different families, show particular patterns. The reticulated sculpture on the prodissoconch surface seems to be a common character for the Nuculanidae, and has been recorded for the genera Nuculana, Sacella Woodring 1925 (
The apparent absence of species of the genus Microgloma along the Brazilian coast wais an artifact, reflecting the logistical difficulties in obtaining material from these depths. Once this material became available, additions to the fauna was brought to light. The description of two new species of Microgloma and the new information on the conchology, and bathymetric and geographical distributions of Microgloma mirmidina and Pristigloma alba contribute to knowledge of the biodiversity of deep–sea mollusks of the Campos Basin and Brazil.
We are indebted to Dr. John Allen for sending Pristigloma alba material, to Ellen Strong, Smithsonian Institution, for sending the image of Microgloma pusilla holotype, to Cleo Dilnei de Castro Oliveira for searching out material at the Museum of Comparative Zoology (Harvard University, USA), to the profitable suggestions made by the reviewers of this manuscript, and to Petrobras S.A for making this material available and for SEM support.