





(C) 2011 Manpreet Singh Pandher. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The genus Kisaura Ross, 1956 (Trichoptera, Philopotamidae, Philopotaminae) is reported from India the first time. Six new species are described and illustrated: Kisaura elongata sp. n., Kisaura eloct sp. n., Kisaura clavata sp. n., Kisaura gangtokensis sp. n., Kisaura truncata sp. n., all from Gangtok (Sikkim) and Kisaura himachalica sp. n. from Barot (Himachal Pradesh). Male genitalia of this genus are distinguishable from those of other genera of the family by the pair of long lateral processes of tergum X, the well-developed mesoventral plates between two segments of the inferior appendages, and by the brush-like row of dark setae on the inner surfaces of the terminal segments (harpago) of the inferior appendages.
Trichoptera, Philopotamidae, Philopotaminae, systematic, Kisaura, Sortosa, new species, Oriental Region, India, male genitalia
Kisaura was established by
This genus is currently represented by 42 species, mostly confined to the Oriental and Palaearctic Regions (
Adults were collected by light traps (mercury vapour bulb and UV) placed near the edge of high altitude streams of the Himalayan belt of India. The species described here were mainly collected from Gangtok (Sikkim) in very dense humid forests at altitudes ranging from 1800 m to 2100 m. One species was collected from Himachal Pradesh. The specimens were preserved in 70% ethyl alcohol with a drop of glycerol added. Pertinent collection and locality data were recorded.
For species level identification it is essential to observe the lateral processes of the Xth tergite which are hidden below the preanal appendages in lateral view and are also not clearly visible even in dorsal view. For accomplishing this, the male genitalia were removed from the specimens and put in 10% KOH solution overnight. After this treatment the genitalia were put in 80% ethyl alcohol with a drop of glycerol and observed for morphological characters. The drawings of various aspects were done with the aid of zoom stereoscopic binocular microscope (with maximum magnification of 120×) fitted with an ocular grid in one eye piece. The final drawings were rendered in black ink. The illustrations were scanned at 600 dpi grayscale, and mounted onto plates in Adobe© Photoshop© 7.0. The genitalic terminology corresponds to
Sortosa obrussa
Spurs: 2, 4, 4; wings with primitive venation except fork I variable: it may be near or considerably beyond sectorial cross vein s, or R2 may be atrophied and 2A of forewing incomplete (
Oriental and Palaearctic Regions.
The genus Kisaura can be easily separated from Dolophilodes Ulmer by a pair of elongate and sclerotized lateral processes of segment X and by the brush-like row of dark setae on the inner surface of apical segment of the inferior appendages which are lacking in Dolophilodes.
urn:lsid:zoobank.org:act:CA276C7B-013B-4C7C-A428-961110C17518
http://species-id.net/wiki/Kisaura_clavata
Figs 1–5In superficial comparison, this species seems somewhat allied to Kisaura moselyi Kimmins, 1955 from North East Myanmar, but the combination of characters, including segment VIII with a deep median V-shaped indentation, clavate preanal appendages, and basal part of the apical segment of the inferior appendage with a stout spine like setae, sets Kisaura clavata sp. n. apart from Kisaura moselyi.
Adult. Color in alcohol entirely fulvous except mesoscutellum flavid; maxillary palp with pale annulations at the joints; head with golden and fulvous pubescence; antenna moderately long, scape:pedicel ratio = 1.66: 1; maxillary palp segments ratios 1: 2: 3: 4: 5 = 1: 2.4: 2.6 : 2.6: 5.3; labial palp segments ratios 1: 2: 3 = 1: 1.75: 3.1. Length of forewing 7 mm, sprinkled with white patches along the posterior margin and covered with moderate, sparse and brownish setae, pterostigma not prominent in both wings; discoidal cell very small in forewing; fork I absent in both wing.
Male Genitalia (Figs 1–5). Sternum VIII without ventral process; tergum VIII with deep median V-shaped zone of spines. Segment IX rather short, sclerotized, quadrate with median prominence on anterolateral side, posterolaterally with shallow, rounded excision, apically setose. Inferior appendages two-segmented; basal segment (coxopodite) little longer and stouter than apical segment (harpago), narrow at its base, broad and rounded towards apex, in lateral view, with two lobes, inferior lobe with tuft of long setae; apical segment with curved row of dark brush-like setae and stout spine-like setae at base on mesal surface in dorsal view. Tergum X membranous, extending beyond apex of segment IX, at base on each side arises blade-like, lateral spiniform process, with articulated spinelet at apex, recurved cephalad and then caudoventrad, reaching up to apex of segment IX. Preanal appendage as long as spiniform process of tergum X and clavate at apex. Phallus membranous, intimately surrounded by tergum X.
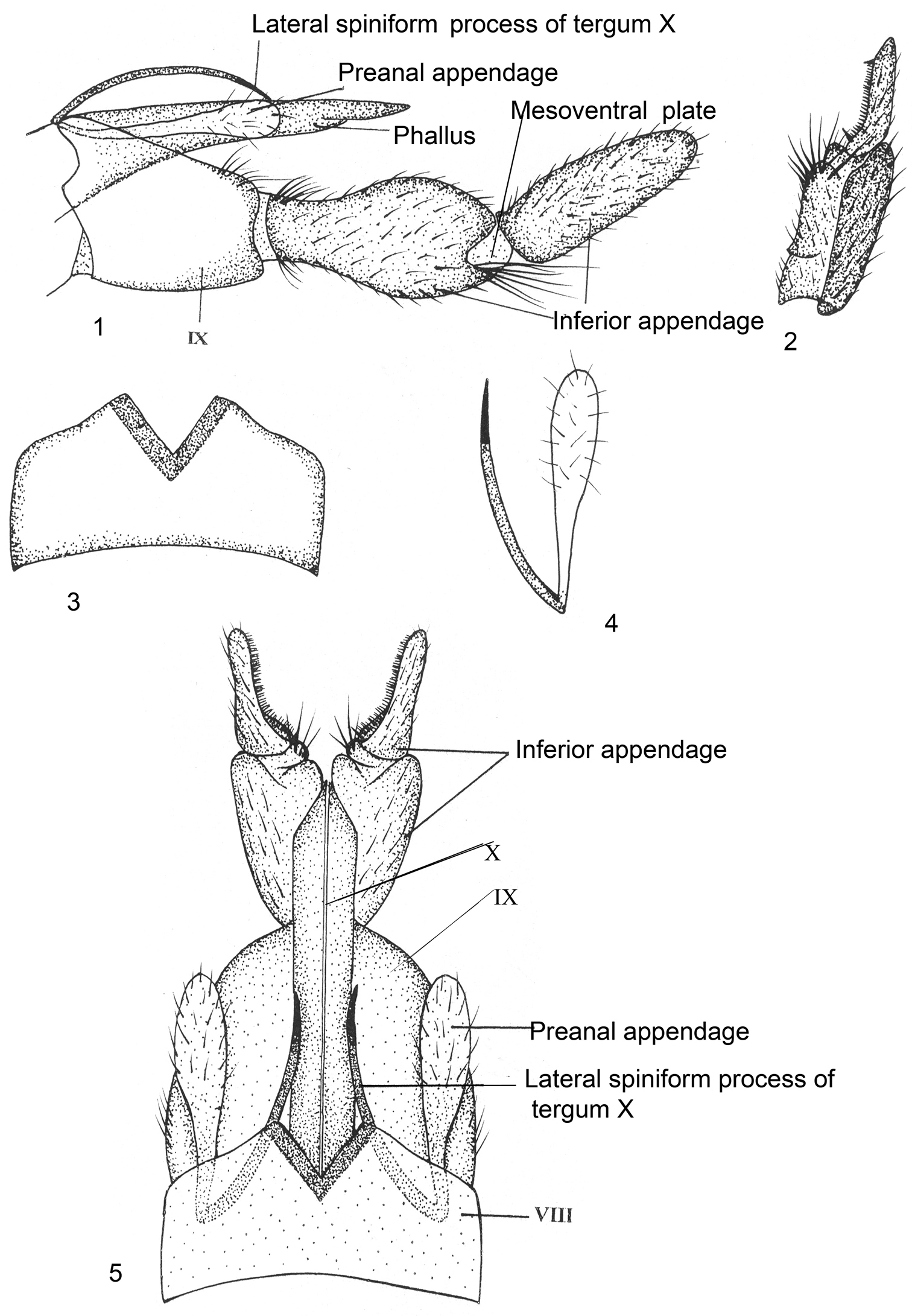
Kisaura clavata sp. n. 1 Male genitalia, lateral 2 Inferior appendage, dorsal 3 Tergite VIII, dorsal 4 Preanal appendage, dorsal 5 Male genitalia, dorsal. Abbreviations: VIII, IX, X = abdominal segments VIII, IX, and X, respectively
Holotype male: INDIA: Sikkim: Gangtok: 27°36'0"N, 88°37'0"E, 1700 m, 25.v.1999, M. Saini, (PUPM).
same data as holotype, 2 males (PUPM).
INDIA: Sikkim: Gangtok: 27°36'0"N, 88°37'0"E, 1, 700m, 15.v.2011, Parey, 1 male (PUPM).
India: Sikkim.
Because of the club-like, (i.e., clavate) apex of preanal appendage, this species has been named clavata.
urn:lsid:zoobank.org:act:B6A4576F-56E1-4811-A60F-031FF8F3535C
http://species-id.net/wiki/Kisaura_elongata
Figs 6–10This species is allied to Kisaura sura Malicky & Chantaramongkol, 1993 and Kisaura consagia Malicky & Chantaramongkol, 1993, both reported from Thailand, because of the presence of bilobed preanal appendages. However, Kisaura elongata is sufficiently distinct from these two species as the two lobes are unequal, i.e., one lobe of the preanal appendage is long while the second is very small, and also in having elongate and inwardly recurved lateral spiniform process of tergum X. In Kisaura sura and Kisaura consagia the two lobes of preanal appendage are almost equal and the lateral spiniform process of tergum X is not inwardly recurved.
Adult. Color in alcohol entirely fulvous excepting antenna with dark brown annulation at joints; body covered with moderate, sparse and fuscous setae excepting head with nigrescent pubescence. Antenna almost equal to length of forewing; scape:pedicel ratio = 1: 0.58; maxillary palp segments ratios 1: 2: 3: 4: 5 = 1: 2: 2.6: 1.6: 4.3 and labial palps segments ratios 1: 2: 3 = 1: 0.8: 1.8. Length of forewing 6 mm, sprinkled with yellow patches, in both wings pterostigma not prominent; discoidal cell almost triangular; apical fork I present in both wings, in forewing positioned considerably beyond sectorial cross-vein s.
Male Genitalia (Figs 6–10). Sternum VIII without ventral process, tergum VIII not indented, posterior margin widely excised with V-shaped mark dorsally. Segment IX sclerotized, short, anterolaterally much produced, posterodorsal corner rounded. Inferior appendages long and stout, basal segment (coxopodite) as long as apical segment (harpago), basal segment with small lobe on proximal end of superior side in lateral view, and distally bilobed with inferior lobe bearing long tuft of setae; apical segment curved upright and pointed apically in lateral view, with curved row of black comb-like setae on inner surface in dorsal view. Tergum X membranous, with long inwardly recurved or bent backward, broad ribbon-like lateral spiniform processes which are pointed apically. Preanal appendages long, with two lobes of unequal size, larger lobe thumb-like while smaller wart-like in lateral view. Phallus membranous, surrounded by tergum X.
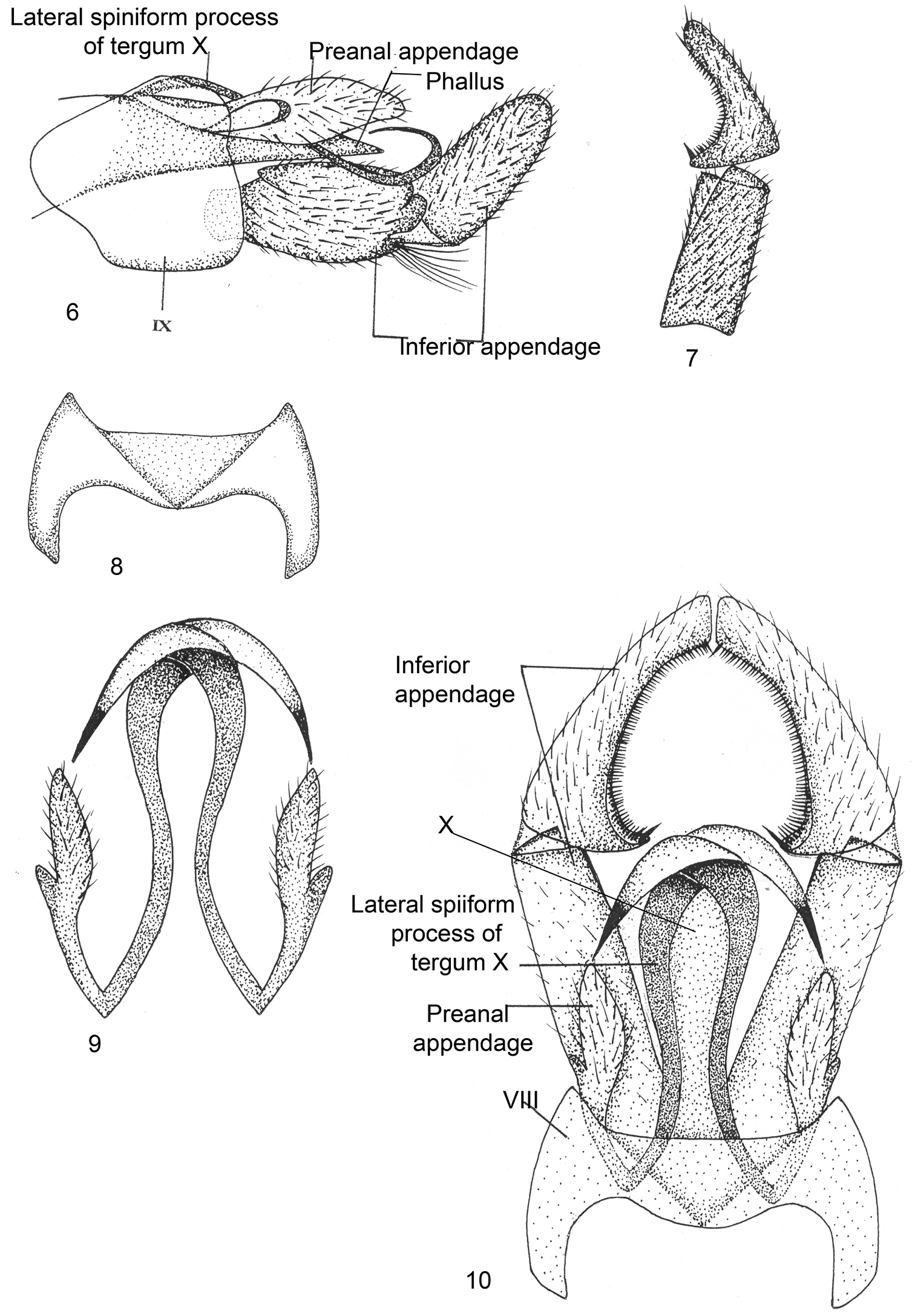
Kisaura elongata sp. n. 6 Male genitalia, lateral 7 Inferior appendage, dorsal 8 Tergite VIII, dorsal 9 Preanal appendages, dorsal 10 Male genitalia, dorsal
Holotype male: INDIA: Sikkim: Gangtok: 27°36'0"N, 88°37'0"E, 1700 m, 25.v.1999, M. Saini (PUPM).
India: Sikkim: Rongli: 27°13'0"N, 88°42'0"E, 900 m, 1.v.2009, Pandher and Parey, 2 males (PUPM).
India: Sikkim.
This species name pertains to excessively long lateral spiniform process of segment X of male genitalia.
urn:lsid:zoobank.org:act:BE5FE988-64E4-4098-836F-34A49EAB5550
http://species-id.net/wiki/Kisaura_eloct
Figs 11–15This species is close to Kisaura longispina
Adult. Color in alcohol entirely fulvous, except mesoscutellum flavid; maxillary palps with pale annulations at their joints. Body covered with dense and fuscous setae except head with fulvous pubescence. Antenna long, scape:pedicel ratio= 1: 0.6; maxillary palp segments ratios 1: 2: 3: 4: 5 = 1: 1.5: 2: 1.2: 2.5 and labial palp segments ratios 1: 2: 3 = 1: 0.75: 1.75. Length of forewing 7 mm; forewing with long discoidal cell; apical fork I absent in both fore- and hind wings.
Male Genitalia (Figs 11–15).Ventral process absent on sternum VIII, tergum VIII with diamond-shaped lobe medially on dorsal surface, with two small incurved depressions on posterior margin. Segment IX sclerotized, long, trapezoidal, stout in lateral view, with posterodorsal margin slightly setose and postventrally produced apically; anterior margin covered partially by sternum VIII. Inferior appendage with basal segment (coxopodite) stout, about as long as apical, narrow at base, broad and bilobed apically in lateral view, inferior lobe with long tuft of setae; apical segment (harpago) curved upwards, with curved row of strong black brush-like setae on inner surface in dorsal view, pointed apically. Tergum X short, narrow, membranous, with pair of long lateral spiniform processes placed medially, extending almost to middle of apical segment of inferior appendage in dorsal view. Preanal appendages narrow, finger-like, scarcely dilated apically, slightly longer than segment IX in lateral view. Phallus tubular, membranous, apparently fused dorsally with membranous tergum X.
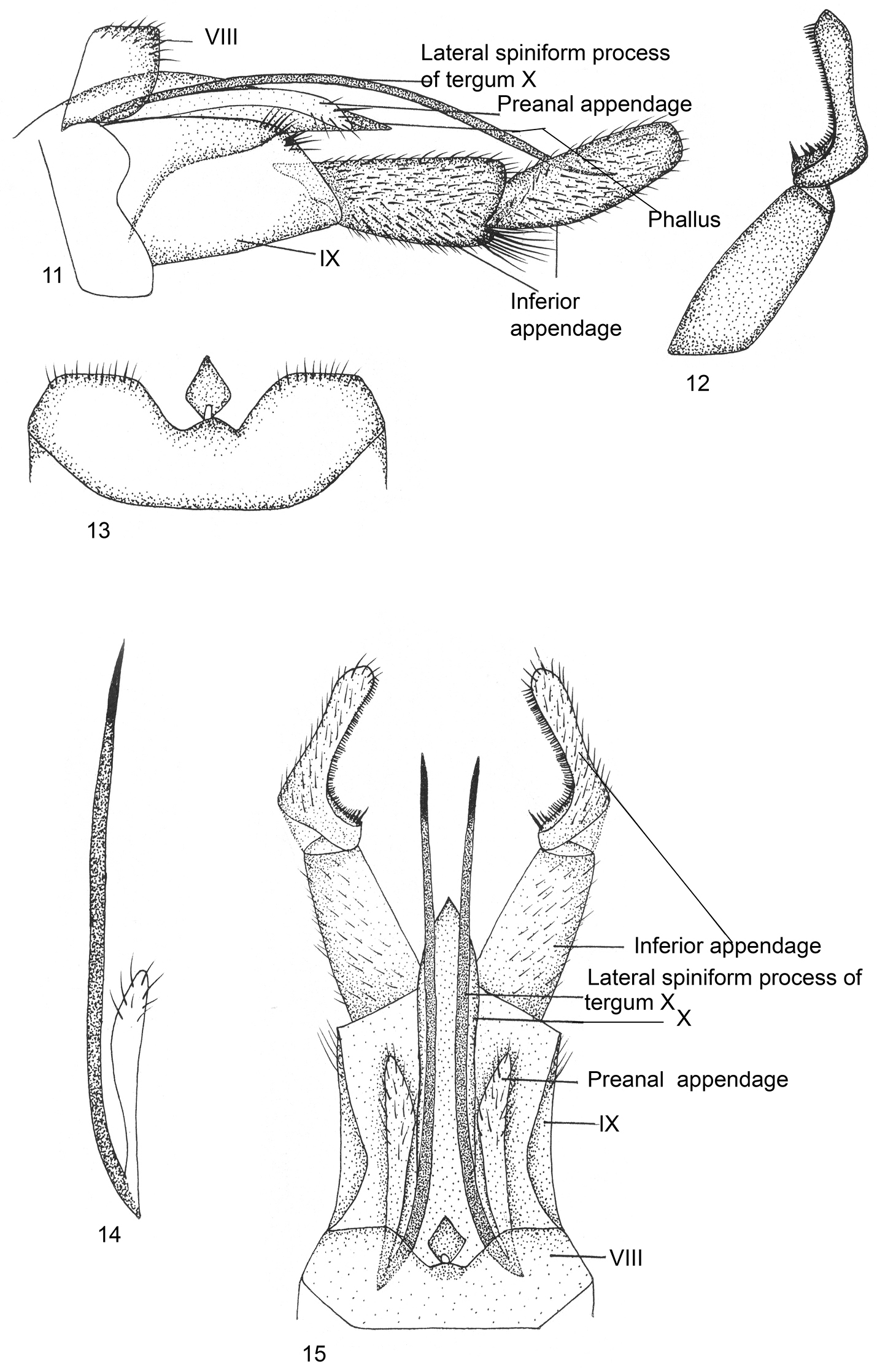
Kisaura eloct sp. n. 11 Male genitalia, lateral 12 Inferior appendage, dorsal 13 Tergite VIII, dorsal 14 Preanal appendage, dorsal 15 Male genitalia, dorsal
Holotype male: INDIA: Sikkim: Gangtok: 27°36'0"N, 88°37'0"E, 1700 m, 25.v.1999, M. Saini, (PUPM).
same data as holotype, 2 males (PUPM).
India: Sikkim.
This species name is based on an arbitrary combination of letters (i.e., Entomology Laboratory of Caddisfly Taxonomy - “eloct").
urn:lsid:zoobank.org:act:38468B63-43EF-4FB9-B32C-072DF504A657
http://species-id.net/wiki/Kisaura_gangtokensis
Figs 16–9This species is close to Kisaura rossi Kimmins, 1955 from North East Myanmar, based on similarities in the shape of the preanal appendages in dorsal view. In Kisaura rossi segment IX is very short, quadrate in lateral view and the preanal appendages are not twisted at mid- length, but preapically twisted. In Kisaura gangtokensis sp. n. segment IX is long, trapezoidal in lateral view, and the preanal appendages are twisted at mid length.
Adult.Color in alcohol entirely fulvous and covered with moderate, fuscous setae excepting head with golden and fulvous pubescence. Antenna long, scape: pedicel ratio = 1: 0.5; maxillary palp segments ratios 1: 2: 3: 4: 5 = 1: 1.3: 2.6: 2.1: 5; labial palp segments ratios 1: 2: 3= 1: 1.5: 2. Length of forewing 7.5 mm; discoidal cell small, apical fork I lacking in both wings.
Male genitalia (Figs 16–19).Sternum VIII without ventral process; tergum VIII roundly produced dorsomesally. Segment IXsclerotized, long, trapezoidal in lateral view, anterodorsally pointed at apex, posteroventrally much produced. Inferior appendages short, stout and two-segmented, both segments subequal in length, basal segment narrow basally, with convex superior and inferior margins, apically bilobed, inferior lobe with long tuft of setae; apical segment vertically upright, its superior margin almost straight, inferior margin convex, rounded apically, mesally with black brush-like setae. Tergum X membranous, intimately surrounding phallus, lateral spiniform process blade-like, long, almost reaching posterodorsal margin of IX in lateral view, with spinelet at apex. Preanal appendagesas long as lateral spiniform process in dorsal view twisted at mid-length, broad and rounded apically. Phallus membranous, fused dorsally with tergum X.
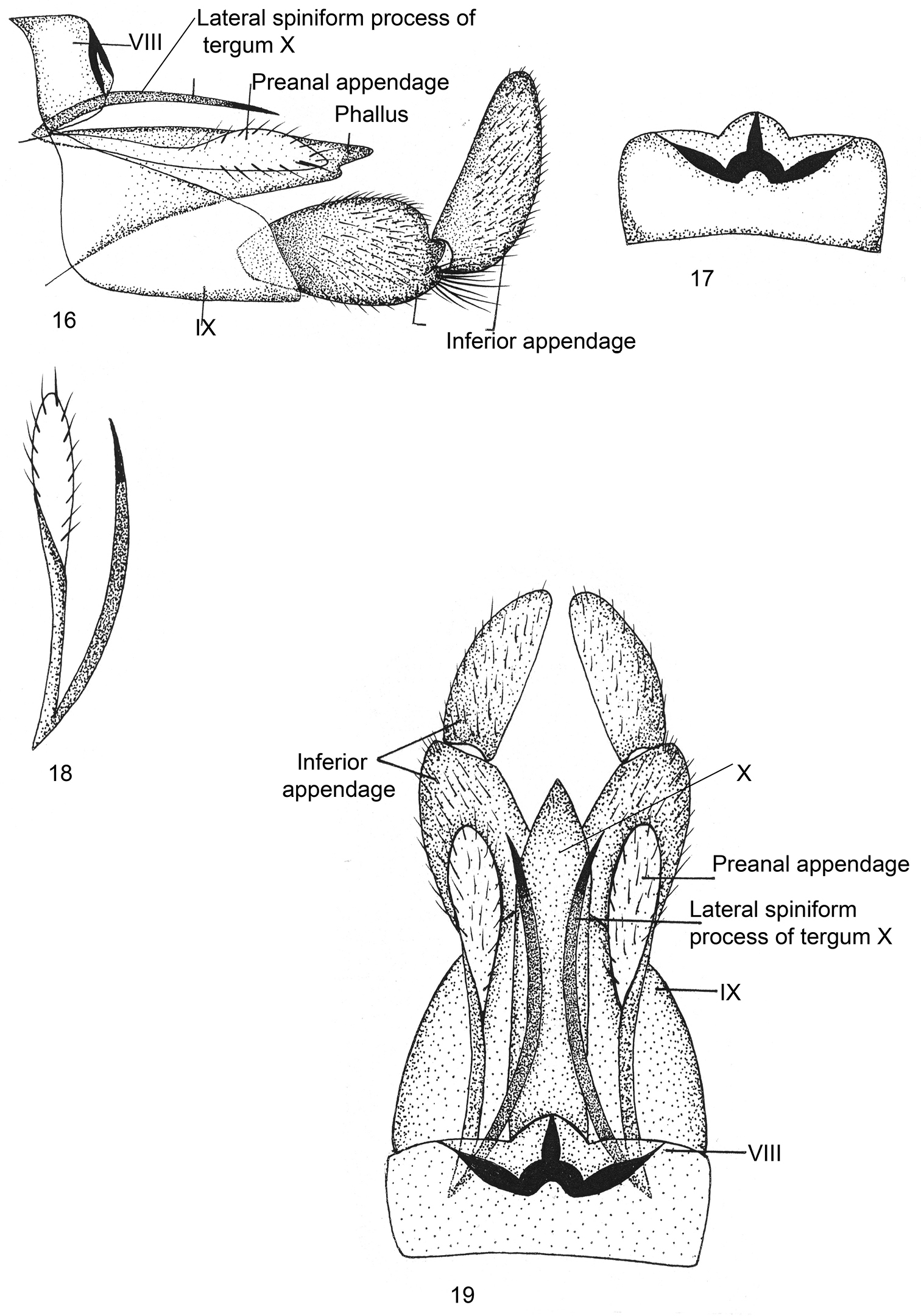
Kisaura gangtokensis sp. n. 16 Male genitalia, lateral 17 Tergite VIII, dorsal 18 Preanal appendage, dorsal 19 Male genitalia, dorsal.
Holotype male: INDIA: Sikkim: Gangtok: 27°36'0"N, 88°37'0"E, 1700 m, 25.v.1999, M. Saini, (PUPM).
same data as holotype, 1 male (PUPM).
India: Sikkim.
This species is named after the type locality, Gangtok.
urn:lsid:zoobank.org:act:995FA64D-0F65-43D9-BF2F-A679AF9649D7
http://species-id.net/wiki/Kisaura_himachalica
Figs 20–24In superficial comparison Kisaura himachalica sp. n. is allied to Kisaura rossi Kimmins and Kisaura cina Malicky & Chantaramongkol, 1993. However Kisaura himachalica is distinct as segment IX is long, the basal segment of the inferior appendage is almost oval in shape, and the preanal appendage is knob-like in dorsal view. In Kisaura rossi segment IX is small, the basal segment of the inferior appendage is quadrate, and thr preanal appendage is twisted preapically and not knob-shaped. The new species also differs in the shape of segment IX, which is broader, and the preanal appendages, which are bilobed and clavate in dorsal view in Kisaura cina.
Adult. Color in alcohol uniformly fulvous and body covered with inconspicuous, sparse and fulvous setae excepting mesoscutellum where is golden. Antenna long, scape:pedicel ratio = 1: 0.7; maxillary palp segments ratios 1: 2: 3: 4: 5 = 1: 1.5: 2.5: 2: 5; labial palp segments ratios 1: 2: 3= 1:1.2: 1.75. Forewing with golden irrorations; length of forewing 7 mm, dicoidal cell small, apical fork I lacking in both wings.
Male genitalia (Figs 20–24).Ventral process absent on sternum VIII; tergum VIII with apical margin produced into two small triangular extensions, separated by shallow excision, area between the two lobes slightly elevated. Segment IX roughly quadrate, anterodorsally pointed, anteromedially with excision, posterodorsally rounded. Inferior appendage with basal segment equal in length to apical segment but stouter than the later, roughly oval in shape, superior and inferior sides convex, divided into two lobes, inferior one with path of long setae, apical segment slender, incurved, with curved row of stout black comb-like setae on inner surface in dorsal view. Tergum X membranous with pair of sword-like lateral spiniform process, about as long as segment IX. Preanal appendage as long as segment IX, clavate in lateral view, apically bulbous and setose in dorsal view. Phallus tubular, membranous, apparently surrounded by Tergum X.
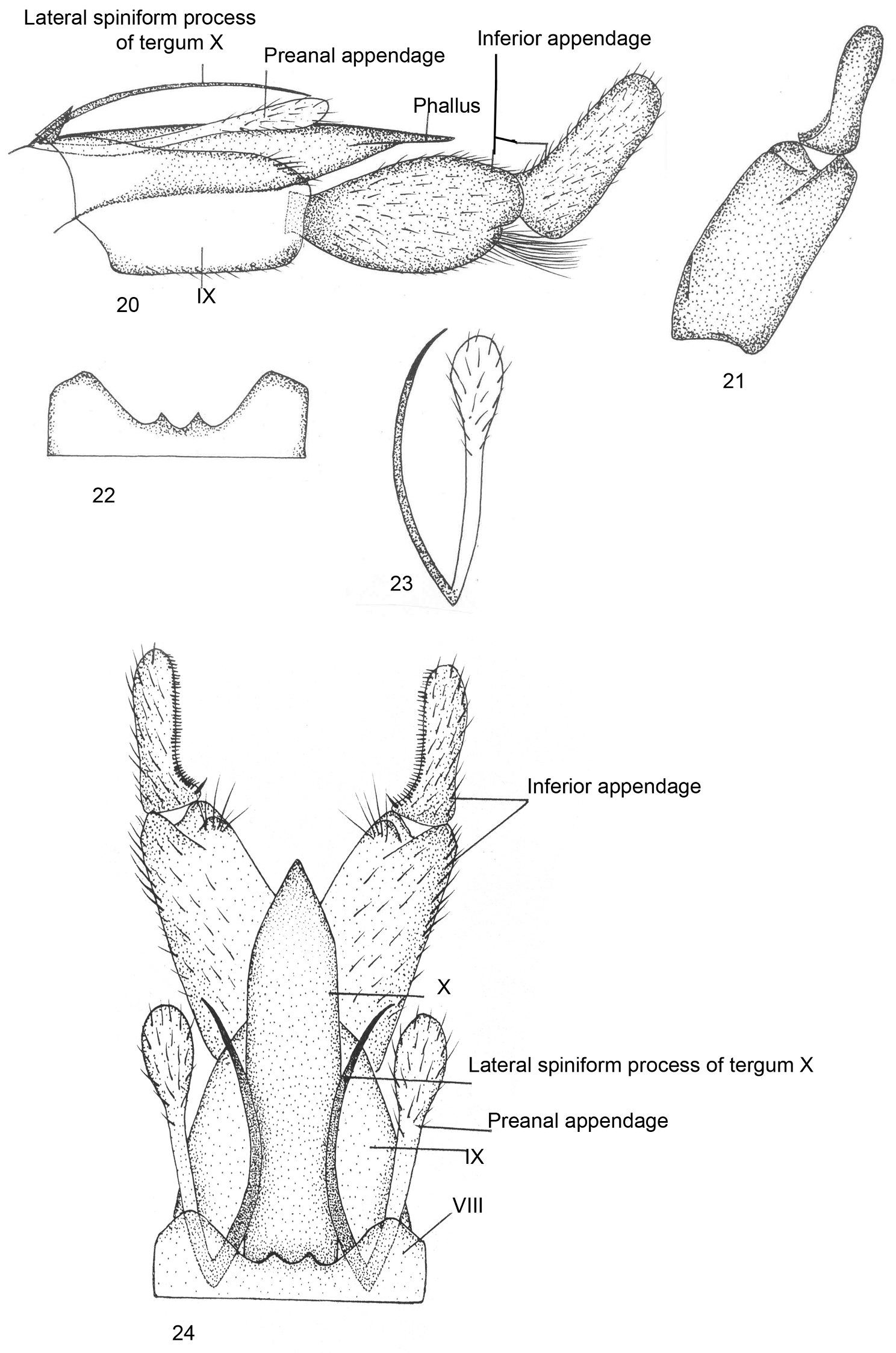
Kisaura himachalica sp. n. 20 Male genitalia, lateral 21 Inferior appendage, dorsal 22 Tergite VIII, dorsal 23 Preanal appendage, dorsal 24 Male genitalia, dorsal
Holotype male: Himachal Pradesh: Barot: 32°17'0"N, 75°77'0"E, 2300 m, 22.v.1998, M. Saini, (PUPM).
India, Himachal Pradesh.
This species is named after the state in which the holotype was found, Himachal Pradesh.
urn:lsid:zoobank.org:act:35D84A5D-ADFC-4BD4-AE82-865821D1DC42
http://species-id.net/wiki/Kisaura_truncata
Figs 25–29This species resembles Kisaura gangtokensis sp. n., based on the ratio of the lengths of the lateral spiniform process and preanal appendages, but differs in several aspects: the preanal appendages are long and slender, segment IX is broad anterolaterally and without rounded lobes at the posterolateral end, and the basal segment of the inferior appendage is long and the apical segment is directed upright and truncate apically.
Adult. Color in alcohol entirely fulvous excepting fuscous mesoscutellum. Antenna almost 1.2 × longer than forewing, scape:pedicel ratio = 1: 0.46; maxillary palp segments ratios 1: 2: 3: 4: 5= 1: 2.6: 3.3: 2.6: 5.6; labial palp segments ratios 1: 2: 3= 1: 1: 2. Forewing with white spots and covered with moderate and short setae; length of forewing 9 mm; discoidal cell small, triangular; apical fork I absent in both wings.
Male genitalia (Figs 25–29). Sternite VIII without ventral process, tergite VIII indented dorsomesally, with rounded median lobe. Segment IX long and broad, roughly pentagonal, anterodorsally with pointed median prominence in lateral view. Inferior appendage two-segmented, basal segment convex on lateral margin, broader, bilobed at apex with short inferior lobe and bearing tuft of long setae; apical segment nearly as long as basal segment, narrower, with curved row of spines, rounded at apex in dorsal view. Tergite X membranous, almost reaching apex of basal segment of inferior appendage, with lateral spiniform processes extending beyond apical margin of segment IX, with spinelet at apex. Preanal appendage almost equal to lateral spiniform process, rounded apically in lateral view. Phallus membranous and fused with tergite X.
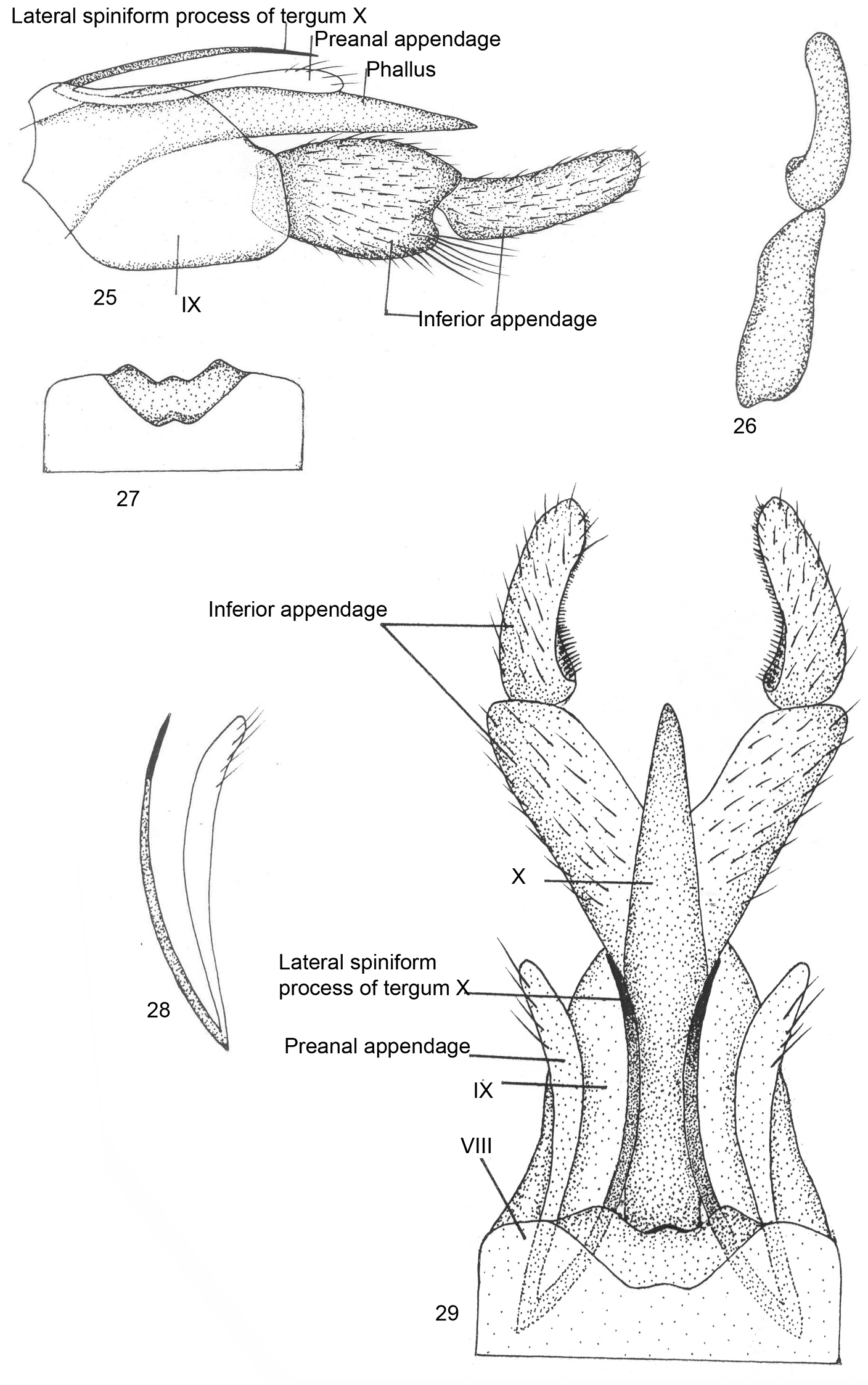
Kisaura truncata sp. n. 25 Male genitalia, lateral 26 Inferior appendage, dorsal 27 Tergite VIII, dorsal 28 Preanal appendage, dorsal 29 Male genitalia, dorsal.
Holotype male: INDIA: Sikkim: Gangtok: 27°36'0"N, 88°37'0"E, 1700 m, 25.v.1999, M.Saini, (PUPM).
INDIA: Sikkim: Singhik: 27°31'0"N, 88°34'0"E, 1900 m, 14.ix.2009, Pandher, 2 males (PUPM).
India: Sikkim.
This species is named for the truncate posterolateral margin of segment IX.
The authors are indebted to Dr John Morse (Clemson University, USA) for providing important literature, valuable suggestions and guidance during the research. We greatly appreciate the support provided in the form of the relevant literature for the Oriental as well as Palaearctic Region by Dr Hans Malicky, Austria. Sincere thanks are due to Dr VV Ramamurthy, National Coordinator, Network Project on Insect Biosystematics (NPIB-21-17), Indian Agricultural Research Institute, New Delhi, Government of India, for providing financial support for this research. Thanks are also due to the parent institute, Punjabi University, Patiala, for providing necessary facilities for the present research. Last, but not least, authors are thankful to the PCCF's, CCF's DFO, and other forest authorities of Himachal Pradesh and Sikkim (especially Mrs Usha Lachungpa, Senior Research Officer, Government of Sikkim) for their help and cooperation during the expedition to these states. Timely support and many-fold assistance provided by laboratory colleagues is also acknowledged.
The authors also appreciate the effort of two anonymous reviewers and the subject editor for Trichoptera in Zookeys for reviewing the manuscript and providing their valuable suggestion to bring the manuscript in its present form.