Research Article |
|
Corresponding author: James W. Hody ( jwhody@ncsu.edu ) Academic editor: Eduardo Eizirik
© 2018 James W. Hody, Roland Kays.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Hody JW, Kays R (2018) Mapping the expansion of coyotes (Canis latrans) across North and Central America. ZooKeys 759: 81-97. https://doi.org/10.3897/zookeys.759.15149
|
Abstract
The geographic distribution of coyotes (Canis latrans) has dramatically expanded since 1900, spreading across much of North America in a period when most other mammal species have been declining. Although this considerable expansion has been well documented at the state/provincial scale, continent-wide descriptions of coyote spread have portrayed conflicting distributions for coyotes prior to the 1900s, with popularly referenced anecdotal accounts showing them restricted to the great plains, and more obscure, but data-rich accounts suggesting they ranged across the arid west. To provide a scientifically credible map of the coyote’s historical range (10,000–300 BP) and describe their range expansion from 1900 to 2016, we synthesized archaeological and fossil records, museum specimens, peer-reviewed reports, and records from wildlife management agencies. Museum specimens confirm that coyotes have been present in the arid west and California throughout the Holocene, well before European colonization. Their range in the late 1800s was undistinguishable from earlier periods, and matched the distribution of non-forest habitat in the region. Coyote expansion began around 1900 as they moved north into taiga forests, east into deciduous forests, west into costal temperate rain forests, and south into tropical rainforests. Forest fragmentation and the extirpation of larger predators probably enabled these expansions. In addition, hybridization with wolves (C. lupus, C. lycaon, and/or C. rufus) and/or domestic dogs has been documented in the east, and suspected in the south. Our detailed account of the original range of coyotes and their subsequent expansion provides the core description of a large scale ecological experiment that can help us better understand the predator-prey interactions, as well as evolution through hybridization.
Keywords
coyote, Canis latrans , range expansion, museum records, FAUNMAP, VertNet, historical ecology, Holocene
Introduction
During the past century, coyotes have undergone a dramatic range expansion across much of North and Central America. Previously restricted to the western two-thirds of North America, the species now occurs across most of the continent, from the Atlantic to the Pacific seaboard and from Alaska to Panama (
Various interacting factors are thought to have contributed to coyotes’ rapid expansion in North America. First, extirpation of apex predators likely helped coyotes expand by reducing predation risk and allowing coyotes to expand their niche to larger prey. Specifically, the extirpation of wolves (C. lupus, C. rufus, and/or C. lycaon) and cougar (Puma concolor) across most of eastern North America, and the decline of cougar and jaguar (Panthera onca) in Central America probably set the stage for coyote colonization (
This ongoing range expansion poses an excellent case study in community ecology and acclimation or adaptation in the Anthropocene, and also presents a new challenge for conservation, as the ecological implications of spreading coyotes are still largely unknown. Coyotes may represent a new top predator in eastern North America and other parts of the continent, with cascading effects on predator communities and disease dynamics (
First, the historic distribution of coyotes prior to the westward expansion of European settlers in the 1800s has recently been confused in the literature. This confusion is largely due to misinterpretation of a figure from
In this popular narrative, coyotes were restricted to true prairie ecosystems prior to European settlement (Figure

However, range maps based on physical evidence (
Additionally, the original northern and southern range limits of coyotes remain uncertain in both narratives (
A second problem with existing large-scale accounts of coyote range is that the recent expansion of coyotes has been coarsely described, without clear spatiotemporal detail. Maps are typically offered without citing reference material, and with few, widely scattered time intervals. Consolidating and improving continent-wide descriptions of coyote range expansion would facilitate testing hypotheses about their effects on newly colonized ecosystems.
Fortunately, coyotes are well represented in museum collections, having been hunted extensively due to their abundance and widespread reputation as a nuisance species. Furthermore, coyotes are also well represented in the fossil and zooarchaeological record, allowing inferences about their distribution several thousand years ago. We compiled museum records from recent biological surveys, fossil and zooarchaeological collections, peer-reviewed literature, and management agency reports to characterize the historical distribution of coyotes prior to European settlement and catalogue their expansion decade-by-decade from 1900 to 2016.
Materials and methods
We compiled coyote occurrences from two data repositories: VertNet (
For our query in FAUNMAP, we searched for excavated remains of coyotes (Canis latrans) from the Holocene epoch (10,000-0 years before present, BP). Taxonomically modern coyotes (C. latrans) also occurred in the late Pleistocene, but biomes and faunal assemblages present in North America at the time drastically differed from those of the Holocene (
Our query in VertNet considered preserved specimens of coyotes (Canis latrans), coydogs (C. latrans × familiaris), and coywolves (C. latrans × lycaon/lupus/rufus) that were collected during 1850–2016. We restricted our query to records that included information about the year and location where the specimen was collected. For quality control reasons, we only considered specimens that included georeferenced point coordinates or enough locality information to reference the data to a specific county. Coyote records from Mexico collected between 1850–1899 were retained as an exception to this rule, because more precise data were not available. In these cases, we allowed records that were referenced to at least a state-level.
In addition to these specimen records, we also compiled first-occurrence records and fossil records of coyotes from peer-reviewed literature and reports by state wildlife management agencies (references listed in Suppl. material
We used these datasets to create two maps. First, we sought to clarify the Holocene distribution of coyotes before large-scale settlement by Europeans using FAUNMAP and a subset of the VertNet data (collected 1850–1899). We also identify which FAUNMAP records had a known minimum age >300 BP to permit stronger inference. Second, we used data from VertNet, peer-reviewed literature, and state management agencies to develop a highly detailed map of 20th century coyote range expansion at 10-year intervals. In both cases, we approximated range boundaries for each historical period (Holocene, 1900, 1910, etc.) by manually constructing polygons around occurrence records from the corresponding time interval.
During the 20th century, coyotes were occasionally brought into areas by hunters and trappers prior to natural expansion into the area (
In the Holocene figure, we also displayed coarse approximations of potential forest cover based on
Results
Our query in FAUNMAP yielded 347 records from the United States and Canada with specific data on the minimum and maximum age of the coyote remains. These were distributed between the Pacific Ocean and the Mississippi River, with the exception of two spatial outliers occurring in New Brunswick, Canada and Florida, USA (Figure

Our query in VertNet yielded 12,319 records of coyotes and coyote hybrids from North and Central America, providing specimen-vouchered coyote occurrences from 1850-2016. Among these records, 4,949 were already georeferenced, and an additional 3,523 records had sufficient locality information to reference the data to individual counties or corresponding political units. An additional 3,747 records could only be referenced to the state- or province-level. We retained such occurrence records for Mexico to address the dearth of available data prior to 1900, but omitted these records elsewhere due to the availability of higher-quality county-level data. Only 100 records had no useable locality information.
Holocene distribution (10,000 BP–1899)
The spatial distribution of coyote specimens from the late 1800s was similar to the distribution of coyote remains older than 300 BP. Specifically, coyotes extended east to Mississippi and Ohio Rivers and west through California and the arid west (Figure
Contemporary expansion (1900–2016)
Combining museum records and regional coyote literature, we created a detailed continent-wide description of coyote range expansion at 10 year intervals (Figure
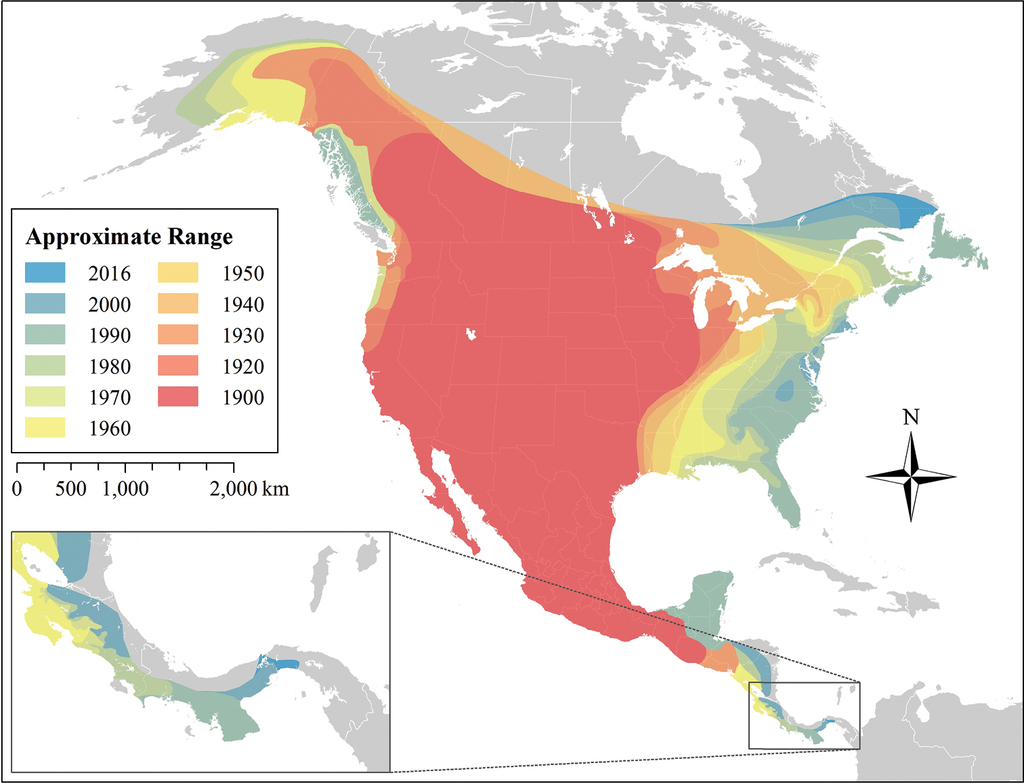
Coyote range expansion by decade, 1900–2016. Ranges are based on occurrence of museum specimens, peer-reviewed literature with associated specimens or photographs, and reports from state wildlife management agencies. The distribution of coyotes between the Yucatán Peninsula and Nicaragua is coarsely depicted due to the paucity of available data, representing the earliest confirmed occurrence. All referenced materials are listed in Suppl. material
Discussion
We compiled coyote occurrences from past biological surveys, fossils, zooarchaeological records, and existing literature to document the historical distribution of coyotes throughout the Holocene and reconstruct decade-by-decade range expansion during 1900–2016. Our findings indicate that coyotes historically occupied a larger area of North America than generally suggested in recent literature, more closely matching the historical range presented by
The distribution of excavated coyote remains 10,000–300 BP matches the distribution of preserved coyote specimens collected between 1850 and 1899 almost identically, suggesting that the geographic range of coyotes in the late 1800s had already been established prior to the 1700s. This same spatial pattern emerged when FAUNMAP data were subdivided in other ways, suggesting that this was not an artifact of how we defined our time intervals. Importantly, Holocene coyote remains ≥4,000 BP showed the same general pattern presented in Figure
Excavated coyote remains and 19th century museum records occurred throughout most non-forested habitats in North America. These specimen records show that coyotes occurred in the Rocky Mountains and Arid West throughout the Holocene, contradicting the proposed western expansion of coyotes during the late-1800s (
The distribution of excavated remains includes four notable outliers, warranting further discussion: one in southern Florida, one in New Brunswick, and two on the Yucatán Peninsula. Although we consider the New Brunswick sample questionable, the Florida and Yucatán specimens might reflect historical range dynamics of coyotes. The Florida record is dated to the early Holocene, but its estimated range age overlaps with the late Pleistocene as well. Coyote fossils from this geological epoch have been documented across the Florida peninsula (
The two Yucatán specimens, both noted by
We cannot definitively assess the Holocene southern limit of coyotes due to paucity of data in Central America. However, we generally agree with
Our map of coyote records from 1900-2016 shows how and when coyotes expanded their range into forested biomes. Agriculture was widespread in these previously forested regions by 1900, so this more open, fragmented landscape presumably aided their expansion, although
Although coyote range expansion into eastern Canada has been well studied (
Likewise, coyote expansion southward across Central America is also not well studied. Coyotes rapidly expanded into deforested habitats in eastern Panama (
Conclusion
The expansion of coyotes across the American continent offers a natural experimental system for assessing ecological questions related to their roles as predators, and evolutionary questions related to their hybridization with dogs and wolves. By collecting and mapping all historical and fossil records of coyotes we were able to correct old misconceptions of their original range, and more precisely map and date their recent expansions. We hope these maps will provide useful context for future research into the ecology and evolution of this incredibly adaptive carnivore.
References
- Adams JR, Leonard JA, Waits LP (2003) Widespread occurrence of a domestic dog mitochondrial DNA haplotype in southeastern US coyotes. Molecular Ecology 12(2): 541–546. https://doi.org/10.1046/j.1365-294X.2003.01708.x
- Andelt WF (1987) Coyote predation. In: Novak M (Ed.) Wild furbearer management and conservation in North America.Ontario Ministry of Natural Resources, Toronto, 128–140.
- Arroyo-Cabrales J, Alvarez T (2003) A preliminary report of the late Quaternary mammal fauna from Loltún Cave, Yucatán, México. In: Schubert BW, Mead J., Graham RW (Eds) Ice Age cave faunas of North America. Indiana University Press, Bloomington, 262–272.
- Bekoff M, Gese EM (2003) Coyote (Canis latrans). In: Feldhamer B, Thompson C, Chapman JA (Eds) Wild mammals of North America: Biology, management, and conservation.Johns Hopkins University Press, Baltimore, 467–481.
- Berger KM, Gese EM (2007) Does interference competition with wolves limit the distribution and abundance of coyotes? Journal of Animal Ecology 76: 1075–1085. https://doi.org/10.1111/j.1365-2656.2007.01287.x
- Bermúdez SEC (2013) Ticks (Acari: Ixodidae, Argasidae) of coyotes in Panama. Systematic and Applied Acarology 18(2): 112–115. https://doi.org/10.11158/saa.18.2.2
- Chubbs TE, Phillips FR (2005) Evidence of range expansion of eastern coyotes, Canis latrans, in Labrador. Canadian Field-Naturalist 119: 381–384. https://doi.org/10.22621/cfn.v119i3.149
- Cluff HD (2006) Extension of Coyote, Canis latrans, breeding range in the Northwest Territories, Canada. Canadian Field-Naturalist 120: 67–70. https://doi.org/10.22621/cfn.v120i1.248
- Constable H, Guralnick R, Wieczorek J, Spencer C, Peterson AT, Bart H, Bates J, Cotter G, Hanken J, Moritz C, Simmons N, Trueb L (2010) VertNet: A new model for biodiversity data sharing. PLoS Biology 8: e1000309. https://doi.org/10.1371/journal.pbio.1000309
- Cook BI, Anchukaitis KJ, Kaplan JO, Puma MJ, Kelley M, Gueyffier D (2012) Pre-Columbian deforestation as an amplifier of drought in Mesoamerica. Geophysical Research Letters 39: 2–6. https://doi.org/10.1029/2012GL052565
- Cove MV, Pardo VLE, Spínola RM, Jackson VL, Sáenz JC (2012) Coyote Canis latrans (Carnivora: Canidae) Range extension in northeastern Costa Rica: Possible explanations and consequences. Latin American Journal of Conservation 3: 82–86.
- Crête M, Desrosiers A (1995) Range expansion of coyotes, Canis latrans, threatens a remnant herd of caribou, Rangifer tarandus, in southeastern Québec. Canadian Field-Naturalist 109: 227–235.
- Crête M, Ouellet J-P, Tremblay J-P, Arsenault R (2001) Suitability of the forest landscape for coyotes in norteastern North America and its implications for coexistence with other carnivores. Ecoscience 8: 311–319. https://doi.org/10.1080/11956860.2001.11682658
- Denevan WM (1992) The pristine myth: The landscape of the Americas in 1492. Annals of the Association of American Geographers 82: 369–385. https://doi.org/10.1111/j.1467-8306.1992.tb01965.x
- Emery KF (1999) Continuity and variability in Postclassic and colonial animal use at Lamanai and Tipu, Belize. In: White CD (Ed.) Reconstructing ancient Maya diet.University of Utah Press, Salt Lake City, 61–82.
- Fener HM, Ginsberg JR, Sanderson EW, Gompper ME (2005) Chronology of range expansion of the Coyote, Canis latrans, in New York. Canadian Field-Naturalist 119: 1–5. https://doi.org/10.22621/cfn.v119i1.74
- Feranec RS (2004) Geographic variation in the diet of hypsodont herbivores from the Rancholabrean of Florida. Palaeogeography, Palaeoclimatology, Palaeoecology 207: 359–369. https://doi.org/10.1016/S0031-0182(04)00047-1
- Feranec RS, MacFadden BJ (2000) Evolution of the grazing niche in Pleistocene mammals from Florida: Evidence from stable isotopes. Palaeogeography, Palaeoclimatology, Palaeoecology 162: 155–169. https://doi.org/10.1016/S0031-0182(00)00110-3
- Georges S (1976) A range extension of the coyote in Quebec. Canadian Field-Naturalist 90: 78–79.
- Gier HT (1975) Ecology and social behavior of the coyote. In: Fox MW (Ed.) The wild canids.Von Nostrand Reinhold, New York, NY, USA, 247–262.
- Gompper ME (2002) Top carnivores in the suburbs? Ecological by colonization issues raised by colonization of north-eastern North America by coyotes. BioScience 52: 185–190. https://doi.org/10.1641/0006-3568(2002)052[0185:TCITSE]2.0.CO;2
- Graham RW, Lundelius EL (2010) FAUNMAP II: New data for North America with a temporal extension for the Blancan, Irvingtonian and early Rancholabrean. FAUNMAP II Database, version 1.0. http://www.ucmp.berkeley.edu/neomap/use.html [July 29, 2016]
- Greene DU, Gore JA (2013) Coyote (Canis latrans) in the Florida Keys. Florida Field Naturalist 41: 126–129.
- Grinnell J, Dixon JS, Linsdale JM (1937) Fur-bearing mammals of California: Their natural history, systematic status and relations to man. University of California Press, Berkeley, 777 pp.
- Hidalgo-Mihart MG, Cantú-Salazar L, González-Romero A, López-González CA (2004) Historical and present distribution of coyote (Canis latrans) in Mexico and Central America. Journal of Biogeography 31: 2025–2038. https://doi.org/10.1111/j.1365-2699.2004.01163.x
- Hidalgo-Mihart MG, Contreras-Moreno FM, Pérez-Solano LA, Hernández-Lara C (2013) Primeros registros de coyote (Canis latrans) en Campeche, México. Revista Mexicana de Biodiversidad 84: 1012–1017. https://doi.org/10.7550/rmb.33108
- Hill EP, Sumner PW, Wooding JB (1987) Human influences on range expansion of coyotes in the southeast. Wildlife Society Bulletin 15(4): 521–524.
- Hody JW (2016) Canid collision - Range expansion by coyotes (Canis latrans) and crab-eating foxes (Cerdocyon thous) in Panama and interpretation of camera trap data. MS thesis, Raleigh, USA: North Carolina State University. https://repository.lib.ncsu.edu/handle/1840.20/33317
- Ingraham RC (2011) Specialized taphonomies in an eastern Maine shell midden: Faunal analysis of Site 62-8, Holmes Point West, Machias, Maine. MS thesis, Orono, USA: University of Maine. http://digitalcommons.library.umaine.edu/etd/1615
- Kays R, Curtis A, Kirchman JJ (2010) Rapid adaptive evolution of northeastern coyotes via hybridization with wolves. Biology Letters 6: 89–93. https://doi.org/10.1098/rsbl.2009.0575
- Kays RW, Gompper ME, Ray JC (2008) Landscape ecology of eastern coyotes based on large-scale estimates of abundance. Ecological Applications 18: 1014–1027. https://doi.org/10.1890/07-0298.1
- Kimmerer RW, Lake FK (2001) The role of indigenous burning in land management. Journal of Forestry 99(11): 36–41.
- Knowlton FF, Gese EM, Jaeger MM (1999) Coyote depredation control: An interface between biology and management. Journal of Range Management 52: 398–412. https://doi.org/10.2307/4003773
- Kuiper K (2010) Pre-Columbian America: Empires of the New World. Britannica Educational Publishing, New York, 194 pp.
- Laliberte AS, Ripple WJ (2004) Range contractions of North American carnivores and ungulates. BioScience 54: 123–138. https://doi.org/10.1641/0006-3568(2004)054[0123:RCONAC]2.0.CO;2
- Leigh EG, O’Dea A, Vermeij GJ (2014) Historical biogeography of the Isthmus of Panama. Biological Reviews 89: 148–172. https://doi.org/10.1111/brv.12048
- Levi T, Kilpatrick AM, Mangel M, Wilmers CC (2012) Deer, predators, and the emergence of Lyme disease. Proceedings of the National Academy of Sciences 109: 10942–10947. https://doi.org/10.1073/pnas.1204536109/-/DCSupplemental.www.pnas.org/cgi/doi/10.1073/pnas.1204536109
- Levy S (2012) Rise of the coyote: The new top dog. Nature 485: 296–297. https://doi.org/10.1038/485296a
- Lucas SG, Alvarado GE, Vega E (1997) The Pleistocene mammals of Costa Rica. Journal of Vertebrate Paleontology 17: 413–427. https://doi.org/10.1080/02724634.1997.10010985
- Macdonald DW, Sillero-Zubiri C (2004) The biology and conservation of wild canids. Oxford University Press Inc., New York, 450 pp. https://doi.org/10.1093/acprof:oso/9780198515562.001.0001
- MacDonald SO, Cook JA (2009) Recent mammals of Alaska. University of Alaska Press, Fairbanks, 387 pp.
- Marshall LG, Webb SD, Sepkoski JJ, Raup DM (1982) Mammalian evolution and the Great American Interchange. Science 215: 1351–1357. https://doi.org/10.1126/science.215.4538.1351
- Meachen JA, Janowicz AC, Avery JE, Sadleir RW (2014) Ecological changes in coyotes (Canis latrans) in response to the Ice Age megafaunal extinctions. PLoS ONE 9: e116041. https://doi.org/10.1371/journal.pone.0116041
- Meachen JA, Samuels JX (2012) Evolution in coyotes (Canis latrans) in response to the megafaunal extinctions. Proceedings of the National Academy of Sciences 109: 4191–4196. https://doi.org/10.1073/pnas.1113788109
- Méndez-Carvajal P, Moreno R (2014) Mammalia, Carnivora, Canidae, Canis latrans (Say, 1823): Actual distribution in Panama. Check List 10: 376–379. https://doi.org/10.15560/10.2.376
- Monge-Nájera J, Morera Brenes B (1987) Why is the coyote (Canis latrans) expanding its range? A critique of the deforestation hypothesis. Revista de Biología Tropical 35: 169–171.
- Moore GC, Parker GR (1992) Colonization by the eastern coyote (Canis latrans). In: Boer AH (Ed.) Ecology and management of the eastern coyote.Wildlife Research Unit, University of New Brunswick, Fredericton, 23–38.
- Nowak RM (1978) Evolution and taxonomy of coyotes and related Canis. In: Bekoff M (Ed.) Coyotes: Biology, behavior, and management.Academic Press, Inc., New York, 3–16.
- Nowak RM (1979) North American Quaternary Canis. Museum of Natural History, University of Kansas, Lawrence, 154 pp.
- Nowak RM (2002) The original status of wolves in eastern North America. Southeastern Naturalist 1(2): 95–130. https://doi.org/10.1656/1528-7092(2002)001[0095:TOSOWI]2.0.CO;2
- Pardi MI, Smith FA (2016) Biotic responses of canids to the terminal Pleistocene megafauna extinction. Ecography 39: 141–151. https://doi.org/10.1111/ecog.01596
- Parker G (1995) Colonization. In: Eastern coyote: the story of its success. Nimbus Publishing, Halifax, 16–35.
- Patterson BR, Messier F (2003) Age and condition of deer killed by coyotes in Nova Scotia. Canadian Journal of Zoology 81: 1894–1898. https://doi.org/10.1139/z03-189
- Peña-Mondragón JL, Castillo Álvarez A, Benítez-Malvido J (2014) Primer registro de coyote (Canis latrans) en la región de La Selva Lacandona, Chiapas, México. Acta Zoológica Mexicana 30(3): 696–700.
- Platt SG, Miller BW, Miller CM (1998) First record of the coyote (Canis latrans) in Belize. Vida Silvestre Neotropical 7: 139–140.
- Ramankutty N, Foley JA (2010) ISLSCP II Potential Natural Vegetation Cover. In: Hall FG, Collatz G, Meeson B, Los S, Brown de Colstoun E, Landis D (Eds) ISLSCP Initiative II Collection. Oak Ridge National Laboratory Distributed Active Archive Center, Oak Ridge, Tennessee, USA. https://doi.org/10.3334/ORNLDAAC/961
- Sacks BN, Brown SK, Ernest HB (2004) Population structure of California coyotes corresponds to habitat-specific breaks and illuminates species history. Molecular Ecology 13: 1265–1275. https://doi.org/10.1111/j.1365-294X.2004.02110.x
- Simpson GG (1980) Splendid isolation: The curious history of South American mammals. Yale University Press, New Haven, 266 pp.
- Sosa-Escalante J, Hernández S, Segovia A, Sánchez-Cordero V (1997) First record of the coyote, Canis latrans, in the Yucatan Peninsula, Mexico. Southwestern Naturalist 42(4): 494–495.
- Stewart FL (1976) Coyote in New Brunswick during prehistoric times. Nature Canada 5: 27. https://doi.org/10.1017/CBO9781107415324.004
- Thornton DH, Murray DL (2014) Influence of hybridization on niche shifts in expanding coyote populations. Diversity and Distributions 20: 1355–1364. https://doi.org/10.1111/ddi.12253
- Van Valkenburgh B, Hertel F (1993) Tough times at La Brea: Tooth breakage in large carnivores of the late Pleistocene. Science 261: 456–459. https://doi.org/10.1126/science.261.5120.456
- Vaughan C (1983) Coyote range expansion in Costa Rica and Panama. Brenesia 21: 27–32.
- vonHoldt BM, Kays R, Pollinger JP, Wayne RK (2016) Admixture mapping identifies introgressed genomic regions in North American canids. Molecular Ecology 25: 2443–2453. https://doi.org/10.1111/mec.13667
- vonHoldt BM, Pollinger JP, Earl DA, Knowles JC, Boyko AR, Parker H, Geffen E, Pilot M, Jedrzejewski W, Jedrzejewska B, Sidorovich V, Greco C, Randi E, Musiani M, Kays R, Bustamante CD, Ostrander EA, Novembre J, Wayne RK (2011) A genome-wide perspective on the evolutionary history of enigmatic wolf-like canids. Genome Research 21: 1294–1305. https://doi.org/10.1101/gr.116301.110
- Wallace AR (1876) The geographical distribution of animals: With a study of the relations of living and extinct faunas as elucidating the past changes of the earth’s surface.
- Weckel M, Bogan DA, Burke RL, Nagy C, Siemer WF, Green T (2015) Coyotes go “bridge and tunnel”: A narrow opportunity to study the socio-ecological impacts of coyote range expansion on Long Island, NY pre- and post-arrival. Cities and the Environment 8: 1–28.
- Weeks JL, Tori GM, Shieldcastle MC (1990) Coyotes (Canis latrans) in Ohio. Ohio Journal of Science 90(5): 142–145.
- Williams JW, Shuman BN, Webb III T, Bartlein PJ, Leduc PL (2004) Late-Quaternary vegetation dynamics in North America: Scaling from taxa to biomes. Ecological Monographs 74: 309–334. https://doi.org/10.1890/02-4045
- Young SP, Jackson HHT (1951) The clever coyote. University of Nebraska Press, Lincoln, 411 pp.
Supplementary materials
Detailed list of references and data sources
Coyote range expansion, 1900–2016