Research Article |
|
Corresponding author: Jeong-Hun Song ( jeonghuns@korea.kr ) Academic editor: Christian Jürgen Schwarz
© 2024 Jaeil Shim, Jeong-Hun Song.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Shim J, Song J-H (2024) A taxonomic review of the order Mantodea in Korea based on morphology and DNA barcodes. ZooKeys 1206: 1-43. https://doi.org/10.3897/zookeys.1206.123355
|
Abstract
A taxonomic study of Korean Mantodea using morphological and molecular characters (COI) is presented. Eight species [Amantis nawai (Shiraki, 1908), Acromantis japonica Westwood, 1889, Mantis religiosa sinica Bazyluk, 1960, Statilia maculata (Thunberg, 1784), Tenodera angustipennis Saussure, 1869, T. sinensis Saussure, 1871, Hierodula chinensis Werner, 1929, H. patellifera (Audinet-Serville, 1838)] belonging to six genera in three families are recognized. Interspecific genetic divergence of COI using uncorrected p-distance ranged from 6.7% to 22.4%, while intraspecific divergence ranged from 0% to 2.2% among eight Korean Mantodea species. All eight species were each strongly supported as a single lineage using COI on both neighbor-joining and parsimony trees. An illustrated key, redescriptions, habitus photographs, and illustrations of diagnostic characters of the species of Korean Mantodea are provided to facilitate identification.
Key words
DNA barcodes, Korea, Mantodea, review, taxonomy
Introduction
The order Mantodea comprise approximately 2,400 species in 460 genera, making it a distinctive group of predatory insects (
Prior to 1995, only a few entomologists had recorded four species of Mantodea on the Korean peninsula, Mantis religiosa Linnaeus, 1758, Statilia maculata (Thunberg, 1784), Tenodera angustipennis Saussure, 1869, and T. aridifolia (Stoll, 1813) (
In this paper, we present a taxonomic reassessment of Korean Mantodea, recognizing eight species belonging to six genera in three families, including A. nawai (Shiraki, 1908), Ac. Japonica Westwood, 1889, M. religiosa sinica Bazyluk, 1960, S. maculata (Thunberg, 1784), T. angustipennis Saussure, 1869, T. sinensis Saussure, 1871, H. chinensis Werner, 1929, and H. patellifera (Audinet-Serville, 1838). Re-descriptions incorporate salient morphological features critical for accurate identification of these species, including the male genitalia. We also used molecular criteria including genetic divergence and gene tree monophyly using a COI barcode region as a multiple lines of evidence approach for the species identification. Our taxonomic review of Korean Mantodea provides redescriptions, habitus photographs, an interactive key, and diagnoses.
Materials and methods
Studied specimens were mostly collected from inland and the islands of the Korean peninsula. Specimens were collected by direct sweeping, scanning, shifting leaf litter and light trapping. If nymphs or oothecae were found, samples were reared until the adult insect emerged. The collected specimens were killed by freezing to prevent discoloration and were moved to a drying chamber for dehydration at 60 °C for 10 days until completely hardened. The subsequent sample preparation followed methods by
The specimens were examined with a stereomicroscope (MS5, Leica Microsystem, Wetzlar, Germany). Images were obtained using a Canon DSLR (EOS 5D; Tokyo, Japan) with an attached Canon MP-E 65 mm f/2.8 1–5× lens. Several layers of photographs were combined in Helicon Focus 5.3 software (Helicon Soft Ltd, Kharkov, Ukraine) and edited using Adobe Photoshop CC 2020 (Adobe, San Jose, CA, USA). Measurements were recorded in millimeters using digital Vernier calipers (CD-15APX; Mitutoyo, Sakado, Japan). The terminology of taxonomic characters and measurements of specimens mainly followed
The following abbreviations are used for the foreleg spination formula and male genitalia: spination formula: Avfs = anteroventral femoral spines; Avts = anteroventral tibial spines; Ds = discoidal spines of forefemur; Pvfs = posteroventral femoral spines; Pvts = posteroventral tibial spines. Male genitalia: afa = anterior process of left phallomere (phalloid apophysis); aafa = anterior lobe of phalloid apophysis; fda = posterior lobe of right phallomere; loa = posterior membranous lobe of left phallomere; pafa = posterior lobe of phalloid apophysis; L4B = a sclerite of left phallomere, mostly spoon-shaped; maa = medial arm process of right phallomere; paa = elongated process of left phallomere, titillator; pia = process arising from the midlenth of the ventral wall of right phallomere, located posterolateral area of pva, strongly sclerotized; pva = process arising from midlenth of the ventral wall of right phallomere, located anteromesal area of pia, strongly sclerotized; sdpl = lateral secondary distal process; sdpm = median secondary distal process.
The following abbreviations are used for the provinces of Korean peninsula (Specimens examined): GW: Gangwon-do; GG: Gyeonggi-do; CB: Chungcheongbuk-do; CN: Chungcheongnam-do; GB: Gyeongsangbuk-do; GN: Gyeongsangnam-do; JB: Jeollabuk-do; JN: Jeollanam-do; JJ: Jeju-do (Is.).
For the study of molecular characters, we included a total of 74 specimens for DNA extraction in the dataset and the specimens used are listed in Suppl. material
Taxonomic accounts
Key to species of Mantodea in Korea
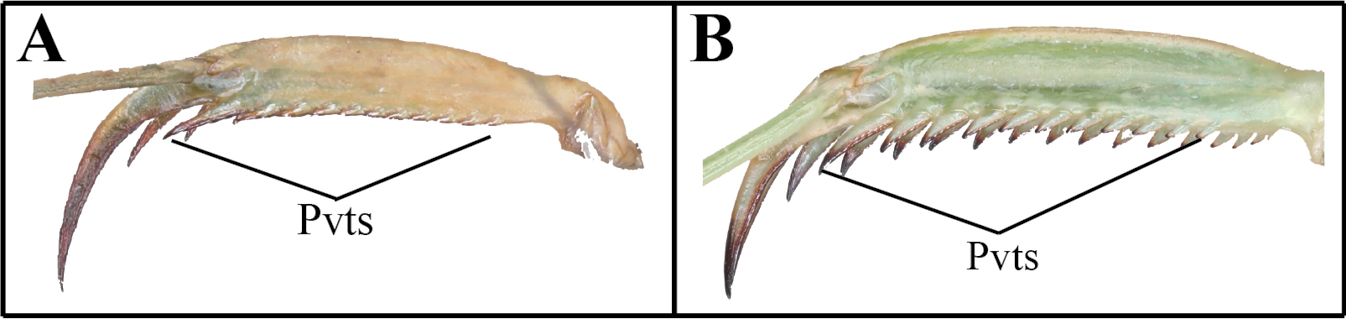
| 1 | (A) Pvts fully decumbent | Acromantis japonica |
| – | (B) Pvts not fully decumbent | 2 |
 |
||
| 2 | (A) Genicular spurs length as long as Pvfs; second Avfs greatly enlarged | Amantis nawai |
| – | (B) Genicular spurs length clearly shorter than Pvfs; second Avfs not enlarged | 3 |
 |
||
| 3 | (A) Forecoxa with a distinct dark or eye spot covering ~ 1/4 to 2/5 of the total length | 4 |
| – | (B) Forecoxa without spot or a faint dark spot covering ~ 1/9 of the total length | 5 |
 |
||
| 4 | (A) Tibial spur groove only with yellow spot or no spot at all | Mantis religiosa sinica |
| – | (B) Tibial spur groove with white spot, and dark patch either side | Statilia maculata |
 |
||
| 5 | (A) Pronotum long and robust; forewing without stigma pattern; abdominal sternites with longitudinally striped pattern | 6 |
| – | (B) Pronotum clavate in shape; forewing with stigma pattern; abdominal sternites without striped pattern | 7 |
 |
||
| 6 | (A) Forecoxa base color yellow (in life); hindwing, area of arculus near cells colored dark brown, subcostal to cubitus area brown to dark brown | Tenodera sinensis |
| – | (B) Forecoxa base color orange (in life); hindwing, area of arculus near cells transparent, subcostal to cubitus area reddish | T. angustipennis |
 |
||
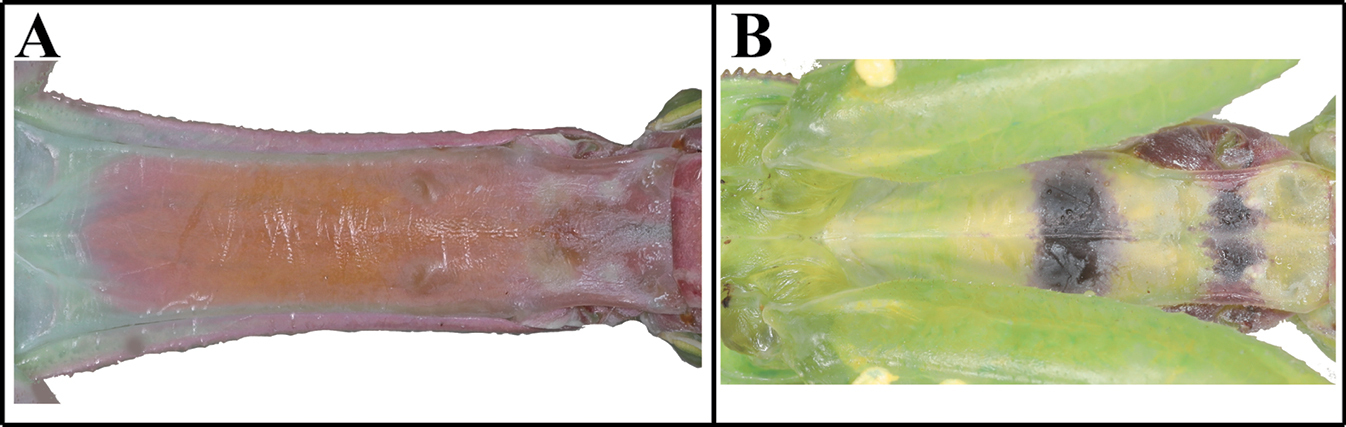
| 7 | (A) Furcasternite without pattern | Hierodula chinensis |
| – | (B) Furcasternite with two-band pattern | H. patellifera |
 |
Species descriptions
Family Gonypetidae Westwood, 1889
Subfamily Iridopteryginae Giglio-Tos, 1915
Amantis
Cimantis
Giglio-Tos, 1915: 154 (synonymized by
Shirakia Beier, 1935: 47.
Type species
Mantis (Oxypilus) reticulata De Haan, 1842.
Diagnosis
Very small sized mantises. Body with mottled dark spot pattern. Pronotum short, kite- shaped, its dorsal surface with longitudinal striped pattern. Genicular spurs length as long as Pvfs length. Second Avfs is as long as second Ds, much longer than the neighboring spines. Wings brachypterous or macropterous.
Amantis nawai
Gonypeta nawai Shiraki, 1908: 47.
Gonypeta maculata Shiraki, 1911: 318.
Amantis nawai
(Shiraki, 1908): ESK and KSAE 1994: 44;
Iridopteryx maculatus
(Shiraki, 1911):
Specimens examined
[NASIC] South Korea: GN: 1♂1♀, Mt. Noja, Dongbu- myeon, Island Geojedo, Geoje-si, 7 VIII 2019, Yeong-Hun Kim; 3♂3♀, Mt. Noja, Dongbu- myeon, Island Geojedo, Geoje-si, 7 VIII 2019, Woojin Jang; JN: 3♂1♀, Island Yeoseodo, Yeoseo-ri, Cheongsan-myeon, Wando-gun, 26 I 2019, Jaeil shim (reared from Ootheca); JJ: 1♀, Andeok Valley, Seogwipo-si, Jeju-do, X 2019, Do-yoon Kim; 2♂1♀, Donnaeko, Seogwipo-si, Jeju-do, 9 IX 2020, Yeong-Hun Kim; 3♂2♀, Seonheul-ri, Jocheon-eup, Jeju-si, Jeju-do, 19 V 2023, Jaeil Shim (reared from ootheca); 2♀, Gamsan-ri, Andeok-myeon, Seogwipo-si, Jeju-do, 23 IX 2023, Jaeil Shim; 1♀, Gamsan-ri, Andeok-myeon, Seogwipo-si, Jeju-do, X 2023, Jisung Kim; Japan: 1♂, Yanbaru, Okinawa, 1–4 I 2020, Wonjun Seong, Forest.
Redescription
Measurements (mm): Body length ♂ 12.1–13.0, ♀ 13.8–15.2; head width ♂ 2.2, ♀ 3.3; head length ♂ 2.9, ♀ 3.9; pronotum width ♂ 2.1, ♀ 2.5; pronotum length ♂ 3.3, ♀ 3.9; forewing (tegmina) length ♂♀ 1.6. Male (Figs


Biological notes
Amantis nawai is distributed throughout the southern islands of the Korean peninsula, as reported by
Distribution
China, Japan, Taiwan, South Korea.
Remarks
Amantis nawai occurs in East Asia (

Family Hymenopodidae Giglio-Tos, 1915
Subfamily Acromantinae Brunner de Wattenwyl, 1893
Acromantis
Type species
Mantis oligoneura De Haan, 1842
Diagnosis
Small-sized mantises. Compound eye with stripe pattern (in live specimens). Anterior area of vertex and lower frons slightly protruding. Pronotum metazone middle area with flat bulge. Genicular spurs clearly shorter than the shortest Pvfs length. Pvts fully decumbent. Meso- and metathoracic femora with weakly expanded postero-ventral femoral lobe. Posterior margins of sternites with a medial lip. Hindwing apex distally truncate.
Acromantis japonica
Acromantis japonica Westwood, 1889: 43.
Acromantis japonica
Westwood, 1889:
Specimens examined
[NASIC] South Korea: GN: 2♀, Mt. Noja, Dongbu-myeon, Island Geojedo, Geoje-si, 10 IX 2016, Jaeil Shim; 1♂, Mt. Wangjo, Nambu-myeon, Tappo-ri, Island Geojedo, Geoje-si, 23 XII 2019, Jun-Ho Lee; 2♀, Mt. Noja, Dongbu-myeon, Island Geojedo, Geoje-si, 29 IX 2016, Byeongmin Jeong; JN: 3♂, Jeongdo-ri, Wando-gun, 10 I 2019, JaeDong Gim (reared from ootheca); 3♂, Jeongdo-ri, Wando-gun, 26 II 2019, Jaeil Shim (reared from ootheca); 8♂6♀, Is. Bogil-do, Bogil-myoen, Wando-gun, 2 III 2023, Jaeil Shim (reared from ootheca); JJ: 1♂, Mt. Sambang, Namjeju-gun, Jeju-si 28 IX 2000, Miae Kim; 1♂, Hwasun-ri, Andeok-myeon, Seogwipo-si, 12–13 IX 2023, Jaeil Shim; 8♂1♀, Sumang-ri, Namwon-eup, Seogwipo-si, 13–14 IX 2023, Jaeil Shim.
Redescription
Measurements (mm): Total length (vertex to tip of abdomen) ♂ 23.3–26.2, ♀ 28.4–32.2; head width ♂ 4.1–4.2, ♀ 5.0–5.1; head length ♂ 2.4, ♀ 3.4; pronotum width ♂ 2.6–2.7, ♀ 3.4; pronotum length ♂ 6.5–7.0, ♀ 8.2–8.4; forewing (tegmina) length ♂ 17.4–20.0, ♀ 17.0–18.5. Male (Figs

Habitus, head, pronotum and foreleg of Acromantis japonica 20 male dorsal aspect 21 female dorsal aspect 22 male face 23 female face 24 female pronotum 25 male furcasternite 26 male foreleg ventral aspect 27 foreleg tibia and tarsus (dorsal aspect) 28 foreleg interior aspect 29 female coxa ventral aspect (live specimens). Red arrows = bulge and protuberance.

Biological notes
Acromantis japonica is found in the southern islands of the Korean Peninsula (
Distribution
China, Japan, Taiwan, North Korea, South Korea.
Remarks
The genus Acromantis has characteristic stripe patterns on their compound eyes when they are alive (see
Family Mantidae Latreille, 1802
Subfamily Mantinae Latreille, 1802
Mantis
Mantes Geoffroy, 1762: 399.
Type species
Gryllus (Mantis) religiosa Linnaeus, 1758
Diagnosis
Medium to large sized mantises. Body color mostly bright green, yellow, brown. Male antenna reddish, conspicuously thicker than the female’s. Vertex swollen. Forecoxal proximal area with dark spot, ventral surface with numerous yellow spots. Tibial spur groove area with yellow spot. Furcasternite with numerous spots. Metathoracical episternum with a dark lateral spot. Male inter-stylar margin notched.
Mantis religiosa sinica
Mantis religiosa sinica Bazyluk, 1960: 255.
Mantis religiosa sinica
Bazyluk, 1960: 255;
Mantis religiosa (Linnaeus, 1758): ESK and KSAE 1994: 44. Korean record.
Specimens examined
[NASIC] South Korea: GW: 3♂, Hotel Ramada, Daegwallyeong Pass, Pyeongchang-gun, 22 VIII 2019, JuHyeong Sohn; GG: 1♀, Jeongok-eup, Yeoncheon-gun, 22 VIII 84, Sunhee Jang; 1♀, Mt. Acha, Gwangjin-gu, Seoul, 8 IX 1977, Sunhee Yoon, Sungshin Univ.; 1 Nymph 1♂4♀, Island Gureopdo, Gureop-ri, Deokjeok-myeon, Incheon, 17 VII 2019, Byeongmin Jeong (reared from nymph); 2♂1♀, Island Gureopdo, Gureop-ri, Deokjeok-myeon, Incheon, 28 VI 2023, Jaeil Shim, Wonjun Sung (reared from nymph); CN: 1♂1♀, Coastal Dune, Sindu-ri, Wonbuk-myeon, Taean-gun, 1 IX 2001, Haechul Park, near the grassland; 4♂6♀, Sonhwang-ri, Woongcheon-eup, Boryeong-si, 12 VII 2023, Jaeil Shim, near the grassland (reared from nymph); 2♂3♀, Coastal Dune, Sindu-ri, Wonbuk-myeon, Taean-gun, 12 VIII 2023, Jaeil Shim, near the grassland; GB: 2♂, Mt. Angi, Songhyeon-dong, Andong-si, VIII 2022, Jaeil Shim; 3 Nymphs, Gyeongjeong Beach, Gyeongjeong-ri, Chuksan-myeon, Yeongdeok-gun, 20 VII 2023, Jaeil Shim; GN: 2♀, Mt. Noja, Dongbu- myeon, Island Geojedo, Geoje-si, 15 IX 2021, Jaeil Shim; JB: 1♂, Byeonsan-myeon, Buan-gun, VIII 1999, Jeonbuk Nat. Univ; 1♂, Mt. Jeoksang, Muju-gun, 8 IX 1999, Taewoo Kim; 1♀, Kunsan Nat. Univ., Gunsan-si, 31 VIII 2019, JuHyeong Sohn; 6♂8♀, Is. Yamido, Okdo-myeon, Gunsan-si, 5 VII 2023, Jaeil Shim (reared from nymph); JJ: 3 Nymphs, Gwangchigi Beach, Goseong-ri, Seongsan-eup, Seogwipo-si, 16 V 2021, Jaeil Shim; Hungary: Mantis religiosa religiosa, 1♂, Mt. Csakyar, Vertes, 23 VIII 2003, J.C. Sohn, Haraszt hegy 250m.
Redescription
Measurements (mm): Total length (vertex to tip of abdomen) ♂ 42.3–55.2, ♀ 50.8–72.4; head width ♂ 5.1–5.4, ♀ 6.2–6.5; head length ♂ 3.8–4.2, ♀ 5.4–5.6; pronotum width ♂ 4.0–4.2, ♀ 5.6–6.0; pronotum length ♂ 13.2–14.0, ♀ 16.6–18.0; forewing (tegmina) length ♂ 38.4–43.2, ♀ 36.1–47.7. Male (Figs

Habitus, head, pronotum and foreleg of Mantis religiosa sinica 34 male dorsal aspect 35 female dorsal aspect 36 male face 37 female face 38 male pronotum 39 female pronotum (inland) 40 female pronotum (Gureopdo island) 41 furcasternite 42 foreleg ventral aspect (live specimens) 43 coxa ventral aspect 44 forleg ventral aspect 45 foreleg interior aspect 46 foreleg dorsal aspect. Abbreviation: Tsg = tibial spur groove. Red arrows = dark spots of furcasternite.
Biological notes
Mantis religiosa sinica is sparsely distributed on the Korean Peninsula. This species prefers broad grasslands and bushy areas, comprised of shrubs and grass in sandy fields as its habitat. It exhibits positive phototaxis, meaning it is attracted to light. When threatened, it makes a hissing sound by rubbing its hindwings against the abdomen. The first instar nymphs hatch from the end of May onwards while the adults emerge in August.
Distribution
China, Japan, South Korea.
Remarks
Mantis religiosa (Linnaeus) is a widely distributed Paleotropical and Holarctic species (

Statilia
Type species
Pseudomantis nemoralis Saussure, 1870
Diagnosis
Medium sized mantises. Vertex nearly flat. Pronotum slender. Ventral surface of foreleg shiny, densely patterned, forcoxa proximal area with dark pattern. Tibial spur groove with whitish yellow to yellow spots. Male inter-stylar margin notched. Male genitalia: sdpl and sdpm well developed, angled at 90°.
Statilia maculata
Mantis maculata Thunberg, 1784: 61.
Pseudomantis haanii Saussure, 1871a: 37, 1871b: 400.
Statilia maculata var. hyalina Giglio-Tos, 1927: 410.
Statilia haani var. major Werner, 1922: 154.
Statilia maculata continentalis Werner, 1935: 495.
Statilia maculata
(Thunberg, 1784): ESK and KSAE 1994: 44;
Statilia nemoralis
(Saussure, 1870):
Specimens examined
[

Habitus, head, pronotum and foreleg of Statilia maculata 52 male dorsal aspect 53 female dorsal aspect 54 female dorsal aspect (green morpho-type) 55 male face 56 female face 57 male pronotum 58 female pronotum 59 furcasternite (brown morpho-type) 60 furcasternite (green morpho-type) 61 femur ventral aspect (green morpho-type) 62 foreleg ventral aspect (live specimens). Abbreviation: Tsg = tibial spur groove. Red arrows = dark spot of furcasternite.
Redescription
Measurements (mm): Total length (vertex to tip of abdomen) ♂ 35.2–43.1, ♀ 43.7–57.7; head width ♂ 4.5–5.0, ♀ 5.4–5.8; head length ♂ 2.9–3.2, ♀ 4.1–4.3; pronotum width ♂ 2.9–3.2, ♀ 4.4–4.6; pronotum length ♂ 12.1–14.0, ♀ 14.0–15.3; forewing (tegmina) length ♂ 26.4–31.2, ♀ 33.1–36.1. Male (Figs

Foreleg, wings, male genitalia and ootheca of Statilia maculata 63 foreleg ventral aspect 64 foreleg interior aspect 65 foreleg dorsal aspect 66 wings 67 subgenital plate 68 male genitalia 69 ootheca (dorsal aspect) 70 ootheca (lateral asepct). Abbreviation: Ts = tibial spur Tsg = tibial spur groove. Red arrow = stigma.
Biological notes
Statilia maculata is found throughout the Korean Peninsula. This species has shown a remarkable adaptability to urban and suburban environments, and is often observed on building walls and streetlights, exhibiting positive phototaxis. They are known to deposit their oothecae under stones and in cracks of tree bark. Notably, S. maculata is capable of producing a hissing sound by rubbing its hindwings and abdomen together. This species typically hatches in early June, with adults emerging in the middle of August.
Distribution
China, Japan, Nepal, South Korea, Taiwan. Invasive in Eastern USA and Russia.
Remarks
Statilia maculata is predominantly found in the eastern Palearctic regions (S. maculata has also been introduced to Eastern USA and Russia), whereas S. nemoralis is described from the Philippines and, erroneously, from various Southeast Asian countries (
Subfamily Tenoderinae Brunner de Wattenwyl, 1893
Tenodera
Mantis (Tenodera) Burmeister, 1838: 534.
Paratenodera Rehn, 1903: 705.
Type species
Mantis fasciata Manuel, 1797
Diagnosis
Large sized mantises. Male body slender, female robust. Ventral surface of forefemur patterned with minute spots. Area between forecoxae attachment point of yellow to orange in color. Hindwing with dark mottled pattern. Abdominal sternites with yellowish longitudinal stripes at middle. Male genitalia: aafa and pafa well developed, pafa spoon- or blade-shaped; loa elongate (
Tenodera angustipennis
Tenodera angustipennis Saussure, 1869: 69.
Tenodera angustipennis
Saussure, 1869: ESK and KSAE 1994: 44;
Specimens examined
[

Habitus, head, pronotum and foreleg of Tenodera angustipennis 71 male dorsal aspect 72 female dorsal aspect 73 male face 74 female face 75 male pronotum 76 female pronotum (small sized) 77 female pronotum (large sized) 78 foreleg ventral aspect 79 foreleg interior aspect 80 foreleg dorsal aspect 81 ventral aspect of foreleg and furcasternite (live specimens). Abbreviation: Tsg = tibial spur groove. Red arrows = dark spot near the tibial spur groove.
Redescription
Measurements (mm): Total length (vertex to tip of abdomen) ♂ 44.2–63.3, ♀ 51.2–86.6; head width ♂ 5.9–6.5, ♀ 6.9–7.8; head length ♂ 4.5–4.9, ♀ 5.6–6.7; pronotum width ♂ 4.3–4.9, ♀ 4.9–6.4; pronotum length ♂ 18.0–22.3, ♀ 19.2–25.0; forewing (tegmina) length ♂ 38.4–48.2, ♀ 38.1–55.2. Male (Figs

Biological notes
Tenodera angustipennis occurs throughout the Korean peninsula and both adults and oothecae can be commonly found on trees and shrubs. First instar nymphs hatch from late May to mid-June, and adults emerge in mid-August.
Distribution
China, India, Japan, Java, South Korea. Invasive in NE USA and Hawaii.
Remarks
Tenodera angustipennis is morphologically similar to T. sinensis in Korea but can be distinguished by their more slender bodies, the pronotum length/width ratio, the orange coloration between the forecoxa base (
Tenodera sinensis
Mantis mandarinea Saussure, 1871a: 289.
Tenodera aridifolia var. sinensis Saussure, 1871b: 417.
Tenodera aridifolia (Stoll, 1813): ESK and KSAE 1994: 44 (misidentification). Korean record.
Tenodera sinensis
Saussure, 1871:
Specimens examined
[

Habitus, head, pronotum and foreleg of Tenodera sinensis 88 male dorsal aspect 89 female dorsal aspect 90 male face 91 female face 92 male pronotum 93 female pronotum 94 ventral aspect of foreleg and furcasternite (live specimens) 95 foreleg ventral aspect 96 foreleg interior aspect 97 foreleg tibia and tarsus (dorsal aspect). Abbreviation: Tsg = tibial spur groove. Red arrows = dark spot near the tibial spur groove.
Redescription
Measurements (mm): Total length (vertex to tip of abdomen) ♂ 54.2–89.1, ♀ 58.2–100.8; head width ♂ 5.9–6.5, ♀ 6.9–7.8; head length ♂ 4.5–4.9, ♀ 5.6–6.7; pronotum width ♂ 4.3–5.6, ♀ 4.9–7.4; pronotum length ♂ 18.0–24.3, ♀ 19.2–27.0; forewing (tegmina) length ♂ 38.4–55.2, ♀ 38.1–63.2. Male (Figs

Wings, abdomen, male genitalia and ootheca of Tenodera sinensis 98 wings 99 male abdomenal sternites (above: green morpho-type below: brown morpho-type) 100 subgenital plate 101 male genitalia (small box = lateral aspect of aafa) 102 ootheca (left: dorsal aspect right above: lateral aspect right below: distal aspect). Red arrow = arculus area.
Biological notes
Tenodera sinensis occurs throughout the Korean peninsula and has adapted well to urban, suburban, and riverside environments. It spawns ootheca in various locations, such as on stones, tree trunks, and branches. First instar nymphs hatch from mid-April to mid-May, while adult mantises typically emerge in mid-August.
Distribution
China, Nepal, Japan, Russia, Thailand, South Korea. Invasive in Canada and the USA.
Remarks
Tenodera sinensis was originally described as a variation of the widely distributed species Tenodera aridifolia (Stoll, 1813), and treated as a subspecies for quite a long time (
Subfamily Hierodulinae Brunner de Wattenwyl, 1893
Hierodula
Parhierodula Giglio-Tos, 1912: 108.
Type species
Hierodula membranacea Burmeister, 1838
Diagnosis
Pronotum clavate. Furcasternite in Korean specimens with spotted pattern or reddish coloration. Forewing stigma triangular, whitish to yellow. Hindwing transparent (Fig.
Hierodula chinensis
Hierodula chinensis Werner, 1929: 75.
Hierodula chinensis
Werner, 1929:
Specimens examined
[NASIC] South Korea: CB: 1 Nymph, Sannam-dong, Seowon-gu, Cheongju-si, 18 VII 2023, NASIC; GB: 1♀, Namtong-dong, Gumi-si 10 X 2023, Jaeil Shim; 1♀, Mangudang park, Hyomok-dong, Daegu-si 29 X 2023, Jaeil Shim; GN: 2♀, Yonggang-ri, Hwagae-myeon, Hadong-gun, 15 IX 2023, Jaeil Shim; JB: 3♂1♀, Jeonbuk Art Museum, Gui-myeon, Wanju-gun, 9 IX 2018, Jaeil Shim; 1♂, Mt. Moak, Gui-myeon, Wanju-gun, 13 IX 2018, Jaeil Shim; 5♂3♀ Mt. Moak, Gui-myeon, Wanju-gun, 7 IX 2019, Jaeil Shim; 6♀, Jeonbuk Art Museum, Gui-myeon, Wanju-gun, 5 IX 2020, Jaeil Shim; 3♂, Jeonbuk Art Museum, Gui-myeon, Wanju-gun, 19 IX 2020, Jaeil Shim; 5♂2♀, National Institute of Agricultural Sciences, Iseo-myeon, Wanju-gun, 20 IX 2023, Jaeil Shim; JN: 2 Nymphs 1♂, Cheongso-ri, Seo-myeon, Suncheon-si, 17 VIII 2019, Jaeil Shim; 2 Nymphs, Haesan-dong, Yeosu-si, 20 VII 2022, Jaeil Shim.

Habitus, head, pronotum, foreleg and hindleg of Hierodula chinensis 103 male dorsal aspect 104 female dorsal aspect 105 male face 106 female face 107 male pronotum 108 female pronotum (live specimens) 109 furcasternite (live specimens) 110 foreleg coxa ventral aspect (right: male left: live female) 111 foreleg ventral aspect 112 foreleg interior aspect 113 foreleg dorsal aspect 114 hindleg femur (interior aspect). Red arrows = dark spot of foreleg trochanter. White arrow = dark spot of joint.

Description
See
Biological notes
Hierodula chinensis is sparsely distributed in the Korean peninsula, primarily inhibiting wooded areas with shrubs and tall trees. It typically lives under tree leaves throughout its lifespan and deposits its oothecae on tree branches. Nymphs have the ability to camouflage themselves by folding their abdomens backwards. The first instar nymphs of H. chinensis hatch in early July, with adult mantises typically emerging at the end of August.
Distribution
China. Invasive in Japan and South Korea.
Remarks
Hierodula chinensis was recently reported in the Korean peninsula by
Hierodula patellifera
Mantis patellifera Audinet-Serville, 1838: 185.
Mantis bipapilla Audinet-Serville, 1838: 188.
Hierodula assamensis
Hierodula manillensis Saussure, 1870: 233.
Hierodula raptoria Stål, 1877: 38.
Hierodula dispar Kirby, 1900: 146.
Hierodula saussurei Kirby, 1904: 245.
Hierodula manillana Giglio-Tos, 1912: 96.
Hierodula (Hierodula) manillana Giglio-Tos, 1927: 448.
Hierodula yunnanensis Wang, 1993: 137.
Hierodula xishaensis Wang, 1993: 140.
Hierodula multispina Wang, 1993: 141.
Hierodula daqinshanensis Wang, 1993: 143.
Hierodula patellifera
(Audinet-Serville, 1838):
Specimens examined
[NASIC] South Korea: GG: 1♀, Seodun-dong, Suwon-si, 2 IX 1999, Taewoo Kim; 1♂, Seodun-dong, Suwon-si, 13 IV–17 VIII 2001, Taewoo Kim (reared); 4♂1♀, SETEC, Daechi-dong, Gangnam-gu, Seoul, 8 VI 2023, Jaeil Shim (reared from nymph); CB: 1♂5♀, Parking area of Cheongju Airport, Ipsang-ri, Naesu-eup, Cheongwon-gu, Cheongju-si, 15 IX 2023, Jaeil Shim; CN: 1♂, Rest area, Geumsan-gun, 10 IX 2013, Haechul Park; 1♂, Mt. Gubong, Gwanjeo-dong, Seo-gu, Daejeon, 1 IX 2014, Taekyu Kim; 1♀, Chungnam Nat. Univ., Yuseong-gu, Daejeon, 27 IX 2015, Taeman Han; 4♂2♀, Samsong-ri, Haemi-myeon, Seosan-si, 24 IV 2023, Jaeil Shim (reared from oothecae); 4♂9♀, Chungnam Nat. Univ., Yuseong-gu, Daejeon-si, 19 VIII 2023, Jaeil Shim; GB: 2♀, Street near Gomo station (Gomo-ro), Suseong-gu, Daegu, 14 IX 2019, Jaeil Shim; 8♀, Hyomok Elementary Scholl, Hyomok-dong, Daegu-si 29 X 2023, Jaeil Shim; GN: 1♀, Mt. Mang, Island Geojedo, Geoje-si, 23 VII 2019, Jun-Gi Lee, Jun-Ho Lee; JB: 2♂2♀, Jeonbuk Nat. Univ., Jeonju-si, 21 VIII 2017, Jaeil Shim; 3♀, National Institute of Agricultural Sciences, Iseo-myeon, Wanju-gun, 4 VII 2018, Jaeil Shim (reared from nymph); 2♂, Jangsu-eup, Jangsu-gun, 5 IX 2019, Jaeil Shim; 2♂3♀, Mt. Moak, Gui-myeon, Wanju-gun, 7 IX 2019, Jaeil Shim; 2♂, Jeonbuk Art Museum, Gui-myeon, Wanju-gun, 21 IX 2019, Jaeil Shim; 4♂6♀, Jeonbuk Art Museum, Gui-myeon, Wanju-gun, 22 IX 2019, Jaeil Shim; 2♂10♀, Iseo-myeon, Wanju-gun, 10 IX 2021, Jaeil Shim; : 5♂3♀, Eunpa Lake, Gunsan-si, VI 2022, Jaeil Shim, JuHyeong Sohn (reared from nymph); 2♀, Byeonsan-myeon, Buan-gun, VIII 2022, Jeonbuk Jaeil Shim; 1♂1♀, Hyangga-ro, Pungsan-myeon, Sunchang-gun, 26 VII 2023, Jaeil Shim (reared from nymph); JN: 1♂1♀, Near the Korea Coast Guard Academy (KCGA), Yeosu-si, 1 IX 2019, Byeongmin Jeong; 1♂1♀, Dal-dong, Mokpo-si, VIII 2020, Jaeil Shim; JJ: 1♀, Jeju Airport, Jeju-si, Jeju-do, 22 IX 2023, Jaeil Shim; 1♂3♀, Donnaeko, Seogwipo-si, Jeju-do, 22 IX 2023, Jaeil Shim; Guam: 1♀, Guam, USA, 19 VII 2017, Yeong-Hun Kim; Japan: 1♂1♀, Yanabaru, Okinawa, Japan, 1–4 I 2020, Wonjun Sung; Java: 1♂1♀, Mt. Argopuro, East Java, Indonesia, V 2019, Jaeil Shim (purchase) Vietnam: 1♂, Quang Trung, Bao Loc, Lam Dong, 20 II 2012, Lam Dong Agro-Foresty Research and Experiment Center.

Habitus, head, pronotum and foreleg of Hierodula patellifera 121 male dorsal aspect 122 female dorsal aspect 123 male face 124 female face 125 male pronotum (small sized) 126 male pronotum (large sized) 127 female pronotum 128 ventral aspect of foreleg and furcasternite (live specimens) 129 foreleg coxa ventral aspect (right: female with 4 forecoxal spines left: male with 2 forecoxal spines) 130 foreleg ventral aspect 131 foreleg interior aspect 132 foreleg dorsal aspect.
Redescription
Measurements (mm): Total length (vertex to tip of abdomen) ♂ 44.3–57.8, ♀ 54.3–74.2; head width ♂ 6.6–7.5, ♀ 8.4–9.7; head length ♂ 5.1–5.7, ♀ 7.3–8.1; pronotum width ♂ 5.1–6.0, ♀ 6.7–8.6; pronotum length ♂ 12.9–15.1, ♀ 16.1–20.2; forewing (tegmina) length ♂ 33.8–40.9, ♀ 36.8–47.5. Male (Figs

Biological notes
Hierodula patellifera occurs throughout the Korean peninsula. This species is well-adapted to urban and suburban environments, and can be easily found near mountains, expressway rest areas, and parks. It typically lives under tree leaves throughout its life cycle and lays its oothecae on tree branches, trunks, and building walls near trees. Nymphs fold their abdomens back to camouflage themselves. First instar nymphs hatch in early June and adult mantises emerge in mid-August.
Distribution
China, Guam, India, Japan, Java, New Guinea, Philippines, Sumba, Taiwan, Thailand, Vietnam, Korea. Invasive in France, Italy, and Hawaii.
Remarks
Hierodula patellifera is a widely distributed species (
DNA barcoding of Korean Mantodea
In total, 56 new sequences from seven species in six genera were generated (657 bp of COI). All new sequences were deposited in GenBank under the accession numbers OQ826709–OQ826764 (Suppl. material

Inter- and intraspecific genetic differences in Korean Mantodea species at each taxonomic level for COI (657 bp) calculated using p-distance.
| Taxonomic level | Mean | SD | Max | Min |
| Intraspecific distances | 0.007 | 0.006 | 0.022 | 0.000 |
| Interspecific distances | 0.150 | 0.025 | 0.224 | 0.067 |
| Interspecific distances of congeners | 0.101 | 0.033 | 0.146 | 0.067 |
| Interspecific distances in family | 0.146 | 0.023 | 0.188 | 0.067 |
Discussion
This study presents the first comprehensive taxonomic review of the Mantodea species in Korea, recognizing eight species based on morphology and DNA barcodes. In contrast to previous studies that primarily focused on the documentation of unrecorded species, our study meticulously examined 494 specimens, encompassing all eight species that have been recorded in Korea. Notably, while the majority of species exhibit a broad distribution across the Korean peninsula (Figs

The morphology of male genitalia is a crucial diagnostic feature for species identification and delineation in mantids, supporting the monophyly of higher taxa (

Inter- and intraspecific genetic differences among Korean Mantodea species for COI (657 bp) calculated using p-distance.
| Amantis nawai | Acromantis japonica | Mantis religiosa | Statilia mculata | Tenodera angustipennis | T. sinensis | Hierodula chinensis | H. patellifera | |
| Amantis nawai | 0–0.003 | |||||||
| Acromantis japonica | 0.165–0.170 | 0–0.003 | ||||||
| Mantis religiosa | 0.159–0.193 | 0.146–0.178 | 0–0.022 | |||||
| Statilia mculata | 0.155–0.182 | 0.153–0.160 | 0.140–0.164 | 0–0.011 | ||||
| Tenodera angustipennis | 0.179–0.188 | 0.152–0.161 | 0.139–0.168 | 0.168–0.188 | 0–0.009 | |||
| T. sinensis | 0.165–0.177 | 0.131–0.143 | 0.123–0.150 | 0.142–0.164 | 0.067–0.083 | 0–0.017 | ||
| Hierodula chinensis | 0.196–0.204 | 0.169–0.176 | 0.146–0.159 | 0.165–0.182 | 0.172–0.181 | 0.143–0.154 | 0–0.011 | |
| H. patellifera | 0.210–0.224 | 0.142–0.153 | 0.150–0.173 | 0.163–0.179 | 0.138–0.154 | 0.122–0.145 | 0.134–0.146 | 0–0.020 |
Although the diversity of Mantodea in Korea is relatively modest when compared to the high species diversity and endemism observed in neighboring China and Japan (
Acknowledgements
We are grateful to A.L. Suzumura (Graduate School of Agriculture Hokkaido University) for her English editing and T. Kim (National Institute of Biological Resources) for providing us with valuable specimens. Comments and suggestions by reviewer E. Shcherbakov (Lomonosov Moscow State University, Russia) and subject editor C.J. Schwarz (Ruhr University Bochum, Germany) significantly improved the manuscript.
Additional information
Conflict of interest
The authors have declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Funding
This work was carried out with the support of the ‘Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01727703)’ and the 2024 Collaborative Research Program between University and Rural Development Administration, Republic of Korea.
Author contributions
Conceptualization: JHS. Data curation: JS. Formal analysis: JHS, JS. Supervision: JHS. Visualization: JS. Writing - original draft: JS. Writing - review and editing: JHS.
Author ORCIDs
Jeong-Hun Song https://orcid.org/0000-0003-0317-1457
Data availability
All of the data that support the findings of this study are available in the main text or Supplementary Information.
References
- An SL (2011) Current Status of Research about Insect Fauna of Natural Reserves in Korea. Journal of Korean Nature 4(4): 273–285. https://doi.org/10.7229/jkn.2011.4.4.273
- Audinet-Serville JG (1838) Histoire naturelle des insectes: Orthoptères. Librairie Encyclopédique de Roret, Paris XVIII, 776 pp. https://doi.org/10.5962/bhl.title.16081
- Battiston R, Leandri F, di Pietro W, Andria S (2019) Mantis, Hierodula e Sphodromantis: aggiornamento su conoscenze e identificazione delle mantidi (Mantodea: Mantinae) native ed aliene presenti in Italia. Scienze e storia dell’ ambiente padano 38: 89–96.
- Bazyluk W (1960) Die geographische Verbreitung und Variabilitat von Mantis religiosa (L.) (Mantodea, Mantidae), sowie Beschreibungen neuer Unterarten. Annales Zoologici (Wars. ) 18: 231–272.
- Bazyluk W (1977) Fauna Polski, Blattodea et Mantodea (Insecta). Polska Akademia Nauk, Warszawa 6: 1–175. [Mantodea: 109–167]
- Befu T (1992) Notes on some ecological traits of Acromantis japonica and A. satsumensis. Battarigisu 93: 23. [in Japanese]
- Beier M (1932) Beiträge zur Fauna sinica. Herausgegeben von R. Mell, Berlin. XIII. Die Mantodeen Chinas. Mitteilungen aus dem Museum für Naturkunde in Berlin. Zoologisches Museum und Institut für Spezielle Zoologie (Berlin) 18: 322–337. https://doi.org/10.1002/mmnz.19320180304
- Beier M (1935) Mantodea. Subfamilie: Mantinae. In: Wytsman P (Ed.) Genera Insectorum, Fascicule 203, 1–146.
- Bey-Bienko G (1930) XLIX. Further studies on the Dermaptera and Orthoptera of Manchuria. Annals & Magazine of Natural History 10(29): 493–500. https://doi.org/10.1080/00222933008673159
- Brannoch SK, Wieland F, Rivera J, Klass KD, Béthoux O, Svenson GJ (2017) Manual of praying mantis morphology, nomenclature, and practices (Insecta, Mantodea). ZooKeys 696: 1–100. https://doi.org/10.3897/zookeys.696.12542
- Bruins E (1999) Terrarien Enzyklopädie. Karl Müller Verlag, Erlangen, 320 pp.
- Brunner de Wattenwyl K (1893) Révision du système des Orthoptères et description des espèces rapportées par M. Leonardo Fea de Birmanie. Annali, Museo Civico de Storia Naturale Giacomo Doria, Genova, Serie 2 13: 230. https://doi.org/10.5962/bhl.title.5121
- Burmeister H (1838) Frangschrecken. Mantodea. In: Burmeister H (Ed.) Handbuch der Entomologie. Zweiter Band. Besondere Entomologie. Zweite Abtheilung. Kaukerfe, Gymnognatha (Erste Hälfte; vulgo Orthoptera). Enslin, Berlin, 517–552.
- Cho PS (1959) A manual of the Orthoptera of Korea. Journal of Colleague Liveral Arts Natrual Science of Korea Univsity 4: 131–198.
- Chou HY (2006) Wing dimorphism of Amantis nawai (Shiraki, 1908) (Mantodea: Mantidae): male A. nawai in Taiwan possesses two wing morphs. (Doctoral dissertation). Unpublished D. Thesis. National Taiwan University’s Institute of Entomology.
- De Haan W (1842) Bijdragen tot de Kennis der Orthoptera. In: Temmink CJ (Ed.) Verhandelingen over de Natuurlijke Geschiedenis der Nederlansche Overzeesche Bezittingen. Natuurkuundige Commissie in Indie, Leiden, 248 pp. [23 pls]
- Doi H (1932) Miscellaneous of Insects. Journal of Chosen Natural History Society 13: 33–39.
- Ehrmann R (2002) Mantodea, gottesanbeterinnen der Welt. Natur und Tier, Münster, 519 pp.
- Ehrmann R, Borer M (2015) Mantodea (Insecta) of Nepal: An annotated checklist. Biodiversität und Naturausstattung im Himalaya 5: 227–274.
- ESK and KSAE (The Entomological Society of Korea and Korean Society of Applied Entomology) (1994) Check List of Insects from Korea. Kon-Kuk University Press, Seoul.
- Geoffroy M (1762) Histoire abrégée des insectes, dans laquelle ces animaux sont rangés suivant un ordre méthodique. Delalain 1, Paris, 28+523 pp. [Mantodea: pp. 399–400] https://doi.org/10.5962/bhl.title.154767
- Giglio-Tos E (1912) Mantidi esotici. V. Mantes, Tenoderae, Hierodulae et Rhomboderae. Bollettino della Società Entomologica Italiana 43: 3–167.
- Giglio-Tos E (1915) Mantidi esotici. Generi e specie nuove. Bollettino della Società Entomologica Italiana 46: 134–200.
- Giglio-Tos E (1927) Das Tierreich. Orthoptera-Mantidae. Walter de Gruyter & Co., Berlin, 707 pp. https://doi.org/10.1515/9783111430669
- Hashimoto K, Suzuki K, Hayashi F (2016) Unique set of copulatory organs in mantises; Concealed female genital opening and extremely asymmetric male gentalia. Entomological Science 19(4): 383–390. https://doi.org/10.1111/ens.12219
- Iwasaki T (1996) Comparative Studies on the life histories of two praying mantises, Tenodera aridifolia (Stoll) and Tenodera angustipennis Saussure (Mantodea: Mantidae): I. temporal pattern of egg hatch and nymphal development. Applied Entomology and Zoology 31(3): 345–356. https://doi.org/10.1303/aez.31.345
- Jensen D, Svenson GJ, Song H, Whiting MF (2010) Phylogeny and evolution of male genitalia within the praying mantis genus Tenodera (Mantodea: Mantidae). Invertebrate Systematics 23(5): 409–421. https://doi.org/10.1071/IS09004
- Jeon JB, Lee SH, Lee SM (1999) Notes on the Praying Mantids (Mantodea) in Korea. Insecta Koreana 16: 225–229.
- Ju DR (1969) Check list of insect classification. Gwahakwon Publish, Pyeongyang.
- Kim TW (2010) Dictyoptera. In: Paek MK (Ed.) Checklist of Korean Insects. Nature and Ecology, Seoul, 31 pp.
- Kim TW (2021) Mantodea. In: Park JK, Lee JE et al. (Eds) Checklist of Insects from Korea. Korean Society of Applied Entomology & The Entomological Society of Korea. Paper and Pencil, Daegu, 65 pp.
- Kimura M (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution 16(2): 111–120. https://doi.org/10.1007/BF01731581
- Kirby WF (1900) Mantodea, In: Andrews CW (Ed.) A monograph of Christmas Island (Indian Ocean): physical Features and geology by Charles W Andrews. With descriptions of the fauna and flora by numerous contributors. Lonemans, London, 146–147.
- Kirby WF (1904) A synonymic catalogue of Orthoptera. Euplexoptera, Cursoria (Forficulidae, Hemimeridae, Blattidae, Mantidae, Phasmidae). Print of the Trust, British Museum, London 1: I–X+501 pp. [Mantodea: pp. 207–316]
- Klass KD (1997) External male genitalia and phylogeny of Blattaria and Mantodea. Zoologisches Forschungsinstitut 42: 1–341.
- Klass KD (2001) Morphological evidence on blattarian phylogeny: “phylogenetic histories and stories” (Insecta, Dictyoptera). Deutsche Entomologische Zeitschrift 48: 223–265. https://doi.org/10.1002/mmnd.4800480203
- Klass KD, Meier R (2006) A phylogenetic analysis of Dictyoptera (Insecta) based on morphological characters. Entomologische Abhandlungen 63: 3–50.
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Molecular Biology and Evolution 35(6): 1547–1549. https://doi.org/10.1093/molbev/msy096
- Kwon YJ, Suh SJ, Huh EY (1996) Insect diversity of Pogil island in Korea. Report on the survey of Natural Environment in Korea 11: 213–261.
- Latreille PA (1802) Histoire, générale et particulière des Crustacés et des Insectes. Sonnini CS Vol. 3. Dufart, Paris, XII + 467 pp. https://doi.org/10.5962/bhl.title.15764
- Lia A (2007) Distribution of Mantis religiosa (L.) and its changes in Poland. Fragmenta Faunistica 50(2): 91–125. https://doi.org/10.3161/00159301FF2007.50.2.091
- Linnaeus C (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus 1, editio decima, reformata. Holmiae, Laurentii Salvii, Stockholm, 847 pp. https://doi.org/10.5962/bhl.title.542
- Liu QP, Liu ZJ, Chen ZT, Yuan ZL, Shi Y (2020) A new species and two new species records of Hierodulinae from China, with a revision of Hierodula chinensis (Mantodea: Mantidae). Oriental Insects 55(1): 99–118. https://doi.org/10.1080/00305316.2020.1754954
- Liu QP, Liu ZJ, Wang G, Yin Z (2021) Taxonomic revision of the praying mantis subfamily Hierodulinae of China (Mantodea: Mantidae). Zootaxa 4951(3): 401–433. https://doi.org/10.11646/zootaxa.4951.3.1
- Manuel B (1797) Histoire Naturelle, Insectes– Mantes. Encyclopédie Méthodique. 7: 619–642.
- Moulin N (2020) When Citizen Science highlights alien invasive species in France: The case of Indochina mantis, Hierodula patellifera (Insecta, Mantodea, Mantidae). Biodiversity Data Journal 8: e46989. https://doi.org/10.3897/BDJ.8.e46989
- Mukherjee TK, Ghosh AK, Hazra AK (1995) The mantid fauna of India (Insecta: Mantodea). Oriental Insects 29(1): 185–358. https://doi.org/10.1080/00305316.1995.10433744
- Mukherjee TK, Iyer G, Chatterjee P (2017) Twenty-three new records of mantodea (Insecta) from some states of India. Journal of Threatened Taxa 9(2): 9829–9839. https://doi.org/10.11609/jott.1936.9.2.9829-9839
- Nakamine H (2016) Mantodea. In: Orthopterological Society of Japan (Ed.) The standard of Polyneoptera in Japan. Gakken Plus, Tokyo, 198–205.
- Nakamine H, Yamasaki K, Naka H (2017) The number of stripes on the compound eyes reflects each instar in Acromantis satsumensis (Mantodea: Hymenopodidae). Applied Entomology and Zoology 52(1): 135–138. https://doi.org/10.1007/s13355-016-0445-0
- Okamoto H (1924) The insect Fauna of Quelpatr island (Saishiu-to). Bulletin of the Agricultural Experiment Station. Government General of Chosen 1, iv+47–223.
- Oshima K (2017) New records of Statilia nemoralis and Tenodera fasciata from Kume Island, the Ryukyus, Japan. Journal of Entomology, New Series 20: 129–131. [in Japanese] https://doi.org/10.20848/kontyu.20.3_129
- Oshima K (2018) New records of six species of Mantodea from some islands adjacent to Okinawa-jima Island, the Ryukyus, Japan. Journal of Entomology, New Series 21: 151–160. (in Japanese). https://doi.org/10.20848/kontyu.21.2_151
- Oshima K (2020) Confirmation of the presence of Statilia maculata (Thunberg) (Mantodea, Mantidae) in Taiwan, with notes on its morphology. Nodai Entomology 2: 1–4.
- Otte D, Spearman L, Stiewe MBD (2024) Mantodea Species File Online. Version 5.0/5.0. http://mantodea.speciesfile.org
- Patel S, Singh R (2016) Updated checklist and distribution of Mantidae (Mantodea: Insecta) of the World. International Journal of Research Studies in Zoology 2(4): 17–54. https://doi.org/10.20431/2454-941X.0204003
- Patel S, Singh G, Singh R (2016) Checklist of global distribution of Hymenopodidae (Mantodea: Dictyoptera: Insecta). International Journal of Current Research 8: 42047–42054.
- Rehn JAG (1903) Studies in Old World Mantidae (Orthoptera). Proceedings. Academy of Natural Sciences of Philadelphia 55: 701–718.
- Roy R (1968) Compléments à la connaissance du genre Mantis: L’identité de M. nobilis et M. octospilota. Bulletin de la Société Entomologique de France 73(7): 174–176. https://doi.org/10.3406/bsef.1968.21013
- Saussure HD (1869) Essai d’un Système des Mantides. Mitteilungen der Schweizerische Entomologische Gesellschaft 3: 49–73.
- Saussure HD (1870) Additions au système des Mantides. Mitteilungen der Schweizerische Entomologische Gesellschaft 3: 221–244.
- Saussure HD (1871a) Mélanges Orthoptérologiques, IIIme Fascicule. IV. Mantides. Mémoires de la Société de Physique et d’Histoire naturelle de Genève 21: 1–214. [pl. 4–6]
- Saussure HD (1871b) Mélanges Orthoptérologiques, Supplément au IIIme Fascicule. H. Georg, Genève & Bâle, 363–460. [pl. 7]
- Schwarz C, Roy R (2019) The systematics of Mantodea revisited: an updated classification incorporating multiple data sources (Insecta: Dictyoptera). Annales de la Société entomologique de France (N.S. ) 55: 101–196. https://doi.org/10.1080/00379271.2018.1556567
- Schwarz C, Ehrmann R, Borer M, Monnerat C (2018) Mantodea (Insecta) of Nepal: Corrections and annotations to the checklist. Biodiversität und Naturausstattung im Himalaya 6: 201–247.
- Shcherbakov E, Anisyutkin L (2018) Update on the praying mantises (Insecta: Mantodea) of South-East Vietnam. Annales de la Société entomologique de France (N.S. ) 54: 119–140. https://doi.org/10.1080/00379271.2018.1447394
- Shcherbakov E, Govorov V (2020) Statilia maculata (Thunberg, 1784) the first invasive praying mantis (Mantodea, Mantidae) in the fauna of Russia. Annales de la Société entomologique de France (N.S. ) 56: 189–202. https://doi.org/10.1080/00379271.2020.1785941
- Shim J, Park H, Ju HJ, Song JH (2021a) The giant Asian mantis Hierodula chinensis Werner (Mantodea: Mantidae) new to Korea. Journal of Asia-Pacific Biodiversity 14(1): 121–126. https://doi.org/10.1016/j.japb.2020.11.001
- Shim J, Park H, Kim S, Ju HJ, Song JH (2021b) Species delimitation of the praying mantis Hierodula patellifera (Audinet-Serville) based on morphological and molecular characters (Mantodea: Mantidae). Zootaxa 4951(1): 147–158. https://doi.org/10.11646/zootaxa.4951.1.7
- Shiraki T (1908) Gonypeta nawai. Shiraki. Insect World 12: 47–50. [in Japanese]
- Shiraki T (1911) Phasmiden und Mantiden Japans. The Zoological Society of Japan. 7: 291–331.
- Shiraki T (1932) Orthoptera of the Japanese Empire Part III. Insecta Matsumurana 22: 113–123.
- Srivathsan A, Meier R (2012) On the inappropriate use of Kimura-2-parameter (K2P) divergences in the DNA-barcoding literature. Cladistics 28(2): 190–194. https://doi.org/10.1111/j.1096-0031.2011.00370.x
- Stål C (1877) Systema Mantodeorum. Essai d’une systematisation nouvelle des Mantodées. Bihang till köngliche Svenska Vetenskaps Akademien Handlingar, Stockholm 4: 1–91.
- Svenson GJ, Roy R (2011) Taxonomic treatment of the endemic Malagasy praying mantis genus Hyalomantis Giglio-Tos, 1915, with a new synonymy and the description of three new species (Mantodea, Iridopterygidae, Tropidomantinae). Zootaxa 2777(1): 1–24. https://doi.org/10.11646/zootaxa.2777.1.1
- Svenson GJ, Vollmer W (2014) A case of the higher-level classification of praying mantises (Mantodea) obscuring the synonymy of Majangella Giglio-Tos, 1915 (Liturgusidae, Liturgusinae) and Ephippiomantis Werner, 1922 (Hymenopodidae, Acromantinae). Zootaxa 3797(1): 103–119. https://doi.org/10.11646/zootaxa.3797.1.10
- Svenson GJ, Hardy NB, Cahill Wightman HM, Wieland F (2015) Of flowers and twigs: phylogenetic revision of the plant‐mimicking praying mantises (Mantodea: Empusidae and Hymenopodidae) with a new suprageneric classification. Systematic Entomology 40(4): 789–834. https://doi.org/10.1111/syen.12134
- Taniguchi I (1987) Notes on breeding Acromantis japonica. Insectarium 24: 24–25. [in Japanese]
- Thunberg CP (1784) Dissertatio entomologica novas insectorum specis sistens, III. Mantodea. Edmann, Jahan. Novae Insectorum Species, Uppsala 3: 53–68.
- Tinkham ER (1937) Studies in Chinese Mantidae (Orthoptera). Lingnan Science Journal 16: 481–499.
- Vyjayandi MC, Rajeesh RS, John PS, Dhanasree MM (2010) On a collection of praying mantids (Insecta: Mantodea) from Goa, India, with new distribution records. Journal of Threatened taxa 1325–1329. https://doi.org/10.11609/JoTT.o2188.1325-9
- Wang TQ (1993) Synopsis on the classification of Mantodea from China. Shanghai Scientific and Technological, Literature Publishing House, Shanghai.
- Wang Y, Zhou S, Zhang Y (2020) Revision of the genus Hierodula Burmeister (Mantodea: Mantidae) in China. Entomotxonomia 42: 1–21.
- Werner F (1922) Philippine mantids, or praying insects. Philippine Journal of Science 21: 147–157.
- Werner F (1929) Über einige Mantiden aus China (Expedition Stötzner) und andere neue oder seltene Mantiden des Museum Dresden. Entomologische Zeitung 90: 74–78. [in German]
- Werner F (1935) Farther communication on Indian mantids or praying insects. Proceedings of the Zoological Society of London 105(3): 495–498. https://doi.org/10.1111/j.1096-3642.1935.tb01676.x
- Westwood JO (1889) Revisio Insectorum Familiae Mantidarum speciebus novis aut minus cognitis descriptis et delineatis. Gurney and Jackson, London.
- Wieland F (2013) The phylogenetic system of Mantodea (Insecta: Dictyoptera). Species. Phylogeny and Evolution 3: 3–222. https://doi.org/10.17875/gup2013-711
- Yager DD, Svenson GJ (2008) Patterns of praying mantis auditory system evolution based on morphological, molecular, neurophysiological, and behavioural data. Biological J ournal of the Linnean Society 94: 541–568. https://doi.org/10.1111/j.1095-8312.2008.00996.x
- Yamasaki T (1981) The taxonomic status of “Iridopteryx maculata” (Mantodea, Mantidae) with notes on its distribution. Memoirs of the National Museum of Nature and Science, Tokyo 14: 95–102.
- Yamasaki K, Schütte K, Nawa T (2022) New record of Chinese reddish mantis, Hierodula chinensis Werner, 1929 (Mantodea, Mantidae) from Japan. Check List 18(1): 147–150. https://doi.org/10.15560/18.1.147
- Zhu X, Wu C, Yuan Q (2012) Mantodea in China. Xiyuan Publishing House, Beijing.
Supplementary material
List of species with their locality data, voucher numbers, and GenBank accession numbers
Data type: xlsx
Explanation note: Asterisks indicate new addition.