Research Article |
|
Corresponding author: Cornelis van Achterberg ( kees@vanachterberg.org ) Academic editor: Jose Fernandez-Triana
© 2023 Cornelis van Achterberg.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
van Achterberg C (2023) Illustrated key to the European genera of Opiinae (Hymenoptera, Braconidae), with the description of two new Palaearctic genera and two new species. ZooKeys 1176: 79-115. https://doi.org/10.3897/zookeys.1176.104850
|
Abstract
An illustrated key to the European genera of the subfamily Opiinae (Hymenoptera, Braconidae) is presented and two new genera are described and illustrated: Cavopius gen. nov. (type species: Opius (Agnopius) daghoides Zaykov & Fischer, 1983) from West and East Palaearctic regions and Pseudosteres gen. nov. (type species: Biosteres adanaensis Fischer & Beyarslan, 2005) from West Palaearctic region. Two new species are described and illustrated: Cephaloplites gijswijti sp. nov. from Greece and Cavopius depressorius sp. nov. from S. Korea. Opius (Hypocynodus) kilisanus Fischer & Beyarslan, 2005 is a new synonym of Cephaloplites mocsaryi Szépligeti, 1897. The following new combinations are proposed: Cavopius daghestanicus (Telenga, 1950), comb. nov., C. daghoides (Zaykov & Fischer, 1983), comb. nov., Pseudosteres adanaensis (Fischer & Beyarslan, 2005), comb. nov., P. arenaceus (Jakimavičius, 1986), comb. nov., P. christenseni (Papp, 1982), comb. nov., P. pseudarenaceus (Fischer & Beyarslan, 2005), comb. nov., and P. riphaeus (Tobias, 1986), comb. nov. Keys to species are provided for Cavopius gen. nov., Cephaloplites Szépligeti, 1897, and Pseudosteres gen. nov.
Key words
Bulgaria, Cavopius, Greece, Korea, new combination, new genus, new synonym, Pseudosteres, Turkey
Introduction
Opiinae is a large subfamily of the family Braconidae with ca 2,000 valid species and 39 genera according to
Among the large collection of Opiinae in Naturalis Biodiversity Center (Leiden) two new genera were discovered and a new species of the rare genus Cephaloplites Szépligeti. The new taxa are described, keyed, and illustrated below and an illustrated key to the genera is provided. This paper is part of the revision of the European species of the subfamily Opiinae.
In this paper the criterium for recognition as a separate (new) genus is the possession of a set of presumably derived characters. The results of molecular research published in
Materials and methods
The specimens were either collected in a Malaise trap or collected by using a sweep net. The Malaise trap specimens were chemically treated with a mixture of xylene + alcohol 96% and amylacetate, respectively (AXA-method;
Morphological terminology follows
Observations and descriptions were made either under an Olympus SZX11 stereomicroscope. Photographic images were taken with a Canon 5Ds 50.6-megapixel camera combined with a Canon MP-E 65 mm f/2.8 1–5× Macro lens, Laowa Macro Twin flash KX-800 and an electronic WeMacro Z-stepper rail. The photos were stacked with Helicon Focus 7 software. The type specimens are deposited in the
Naturalis collection (
Key to European genera of Opiinae
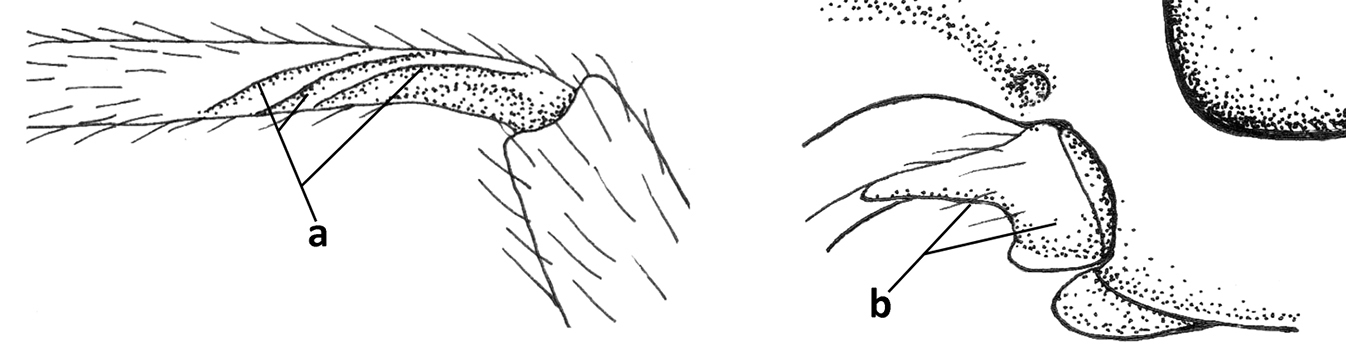
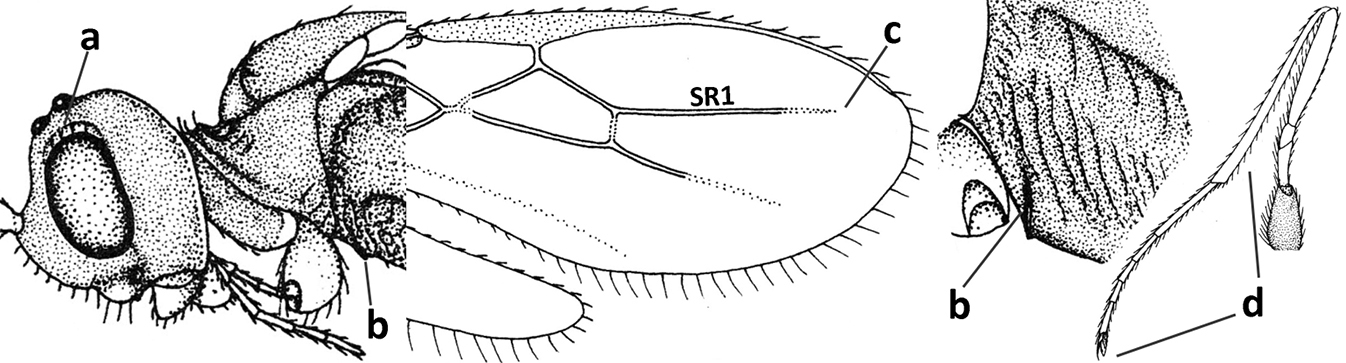
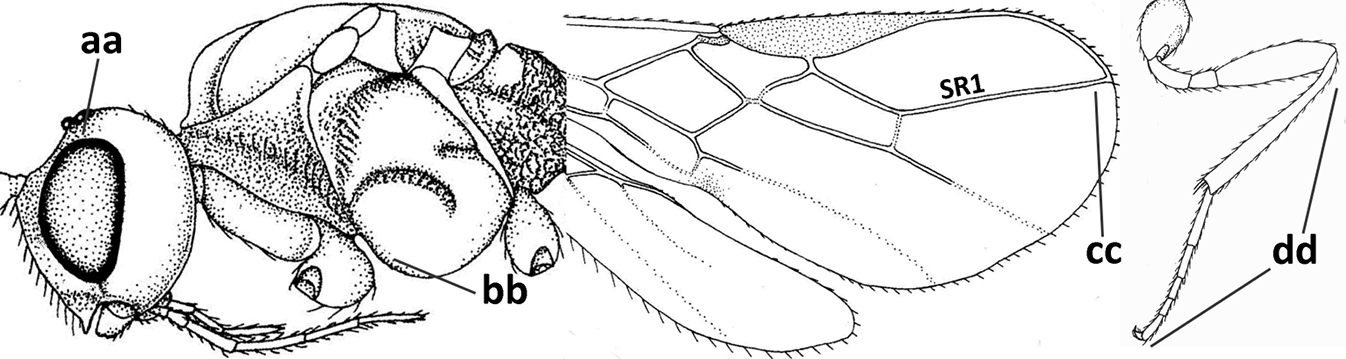
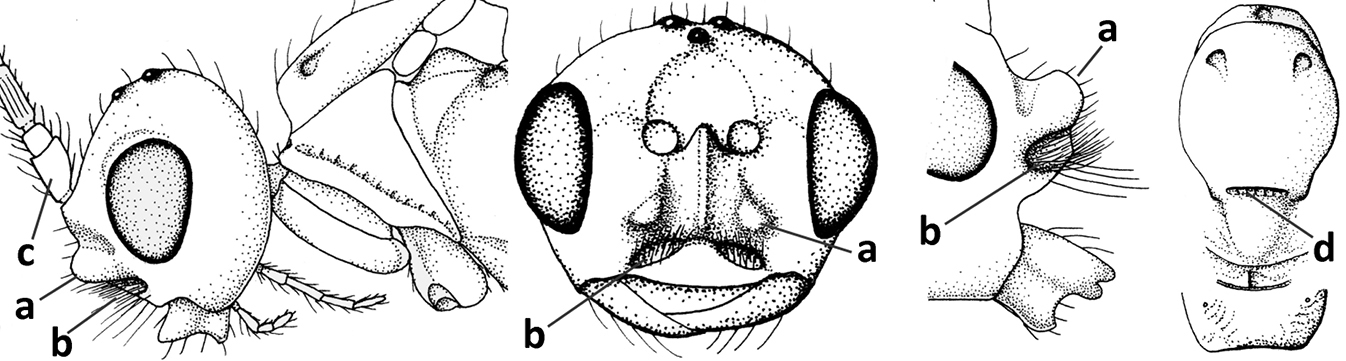
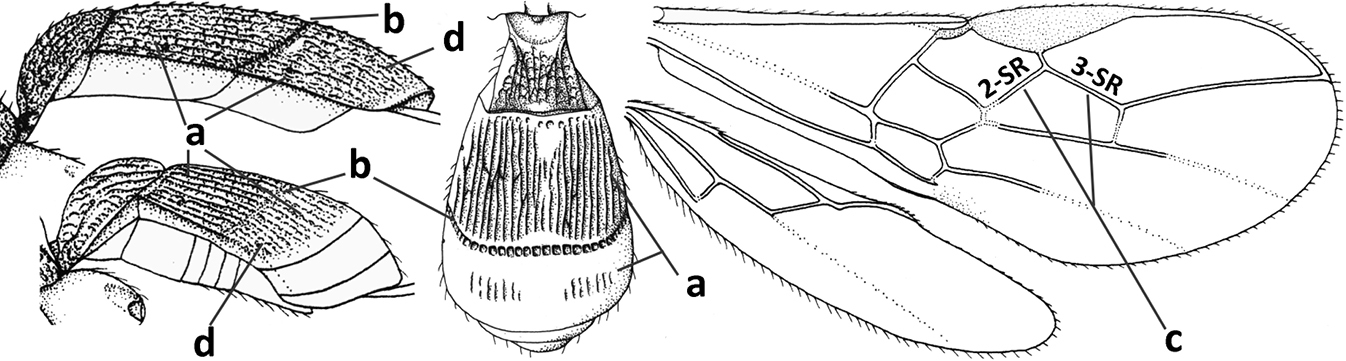
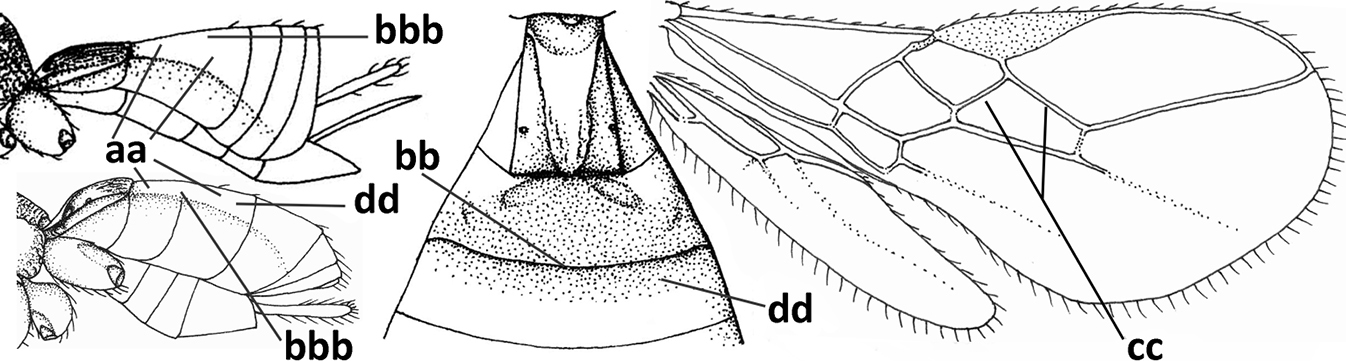
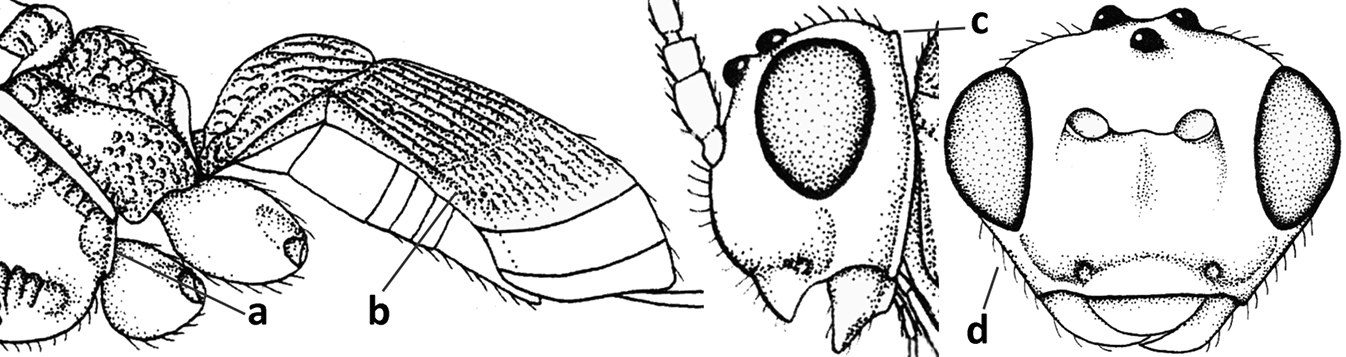
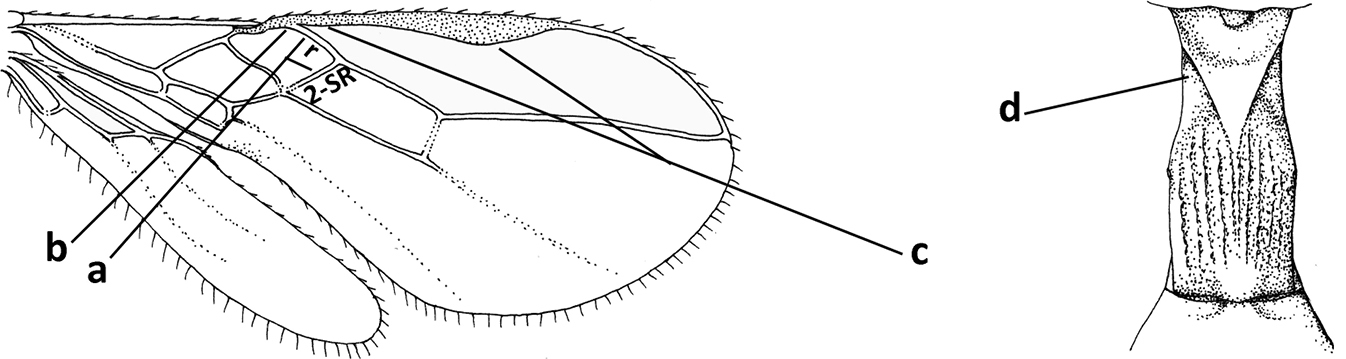
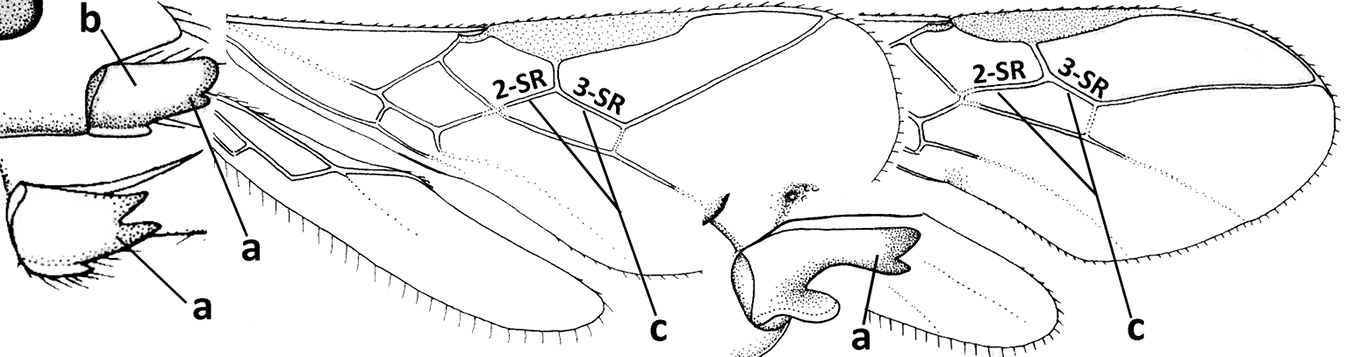
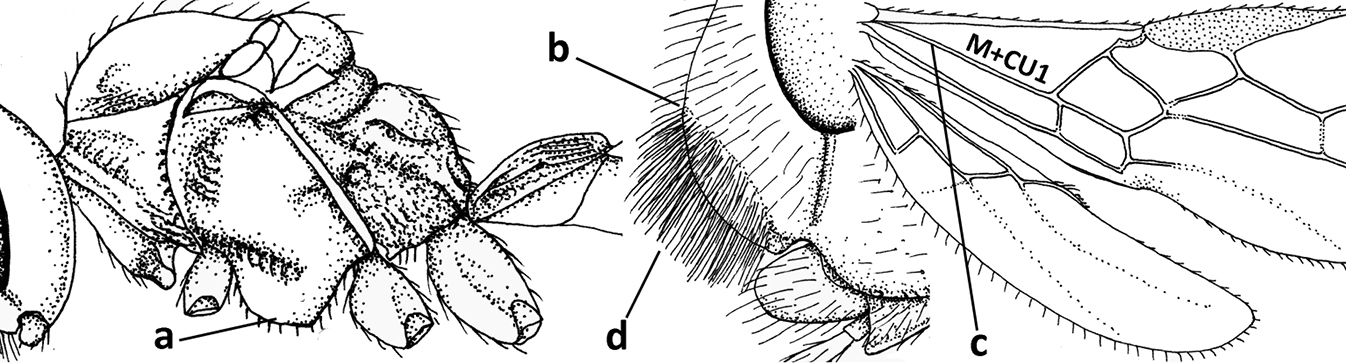
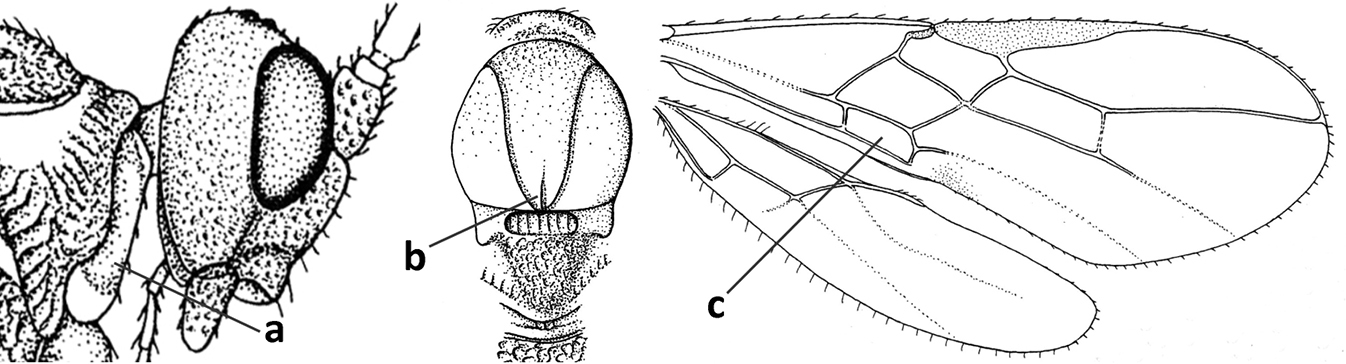
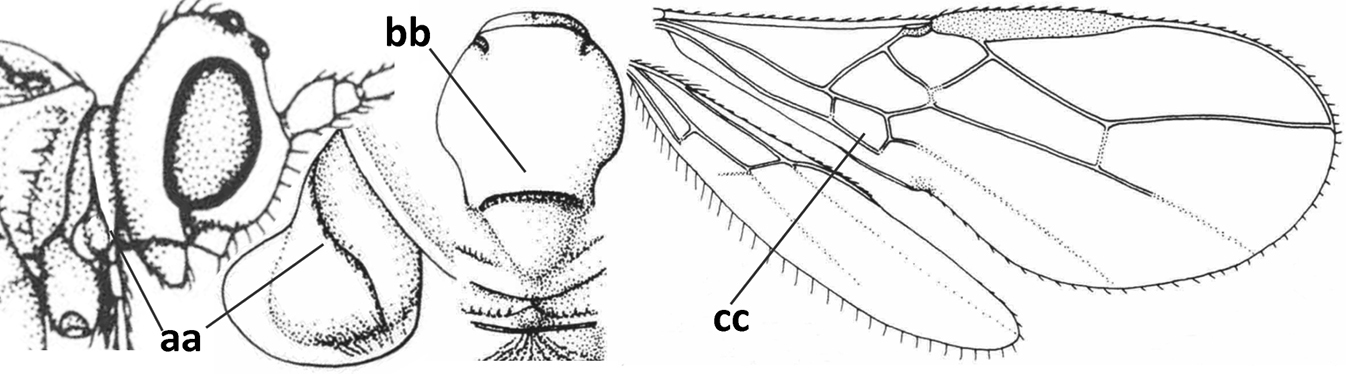
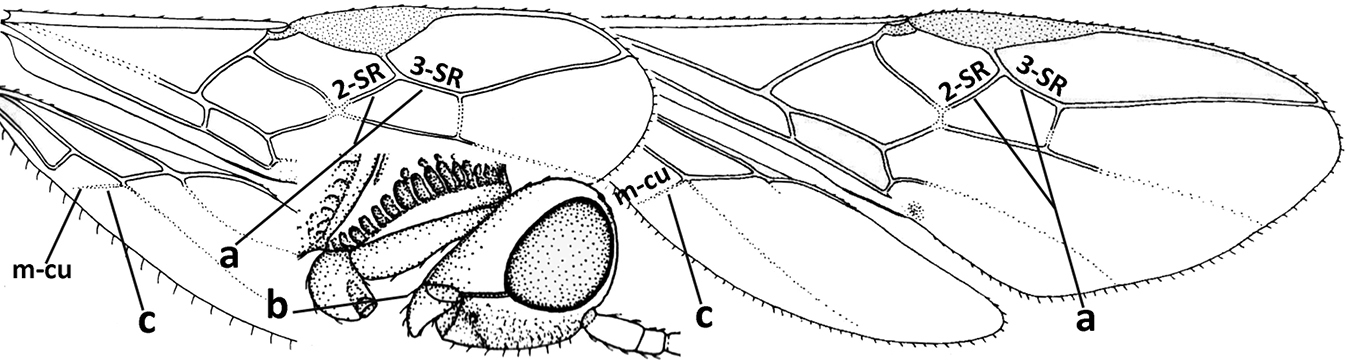
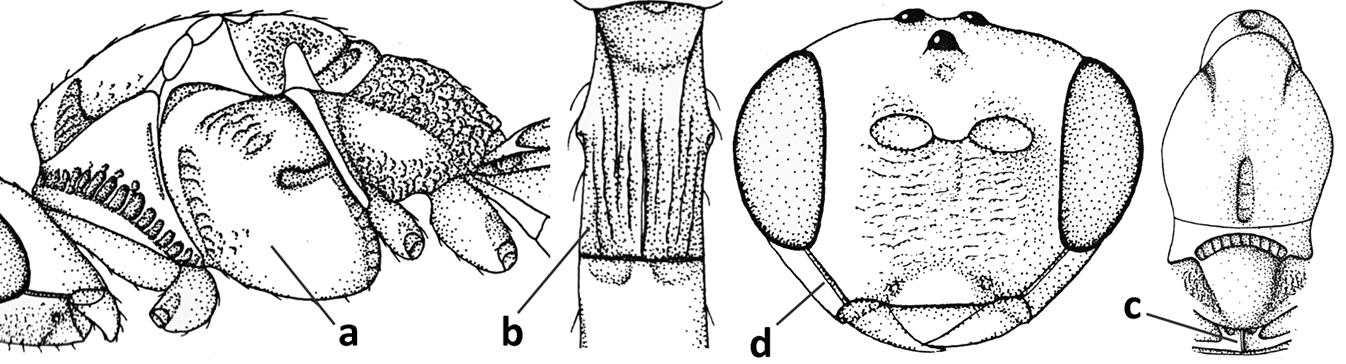
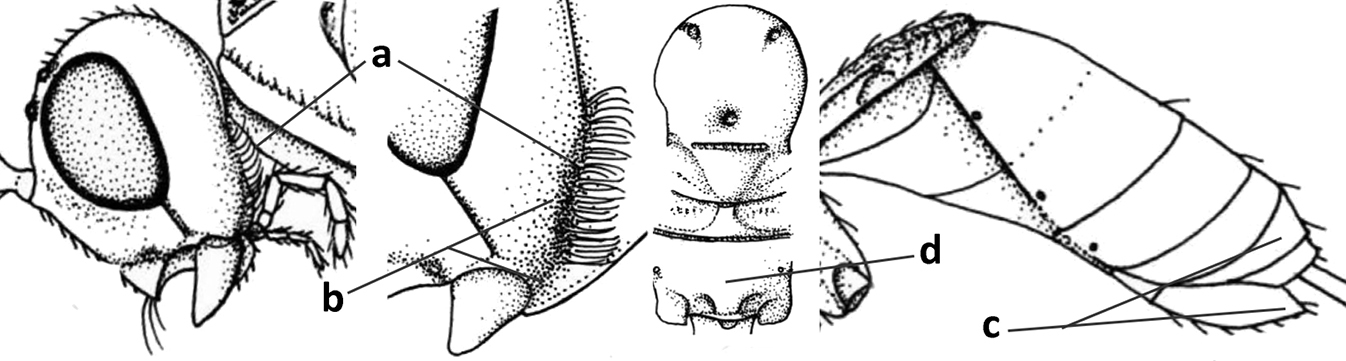
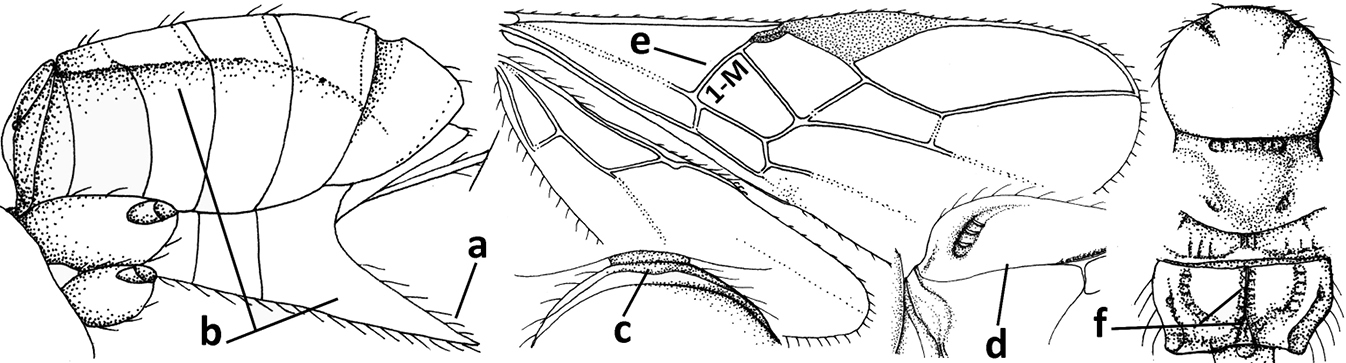
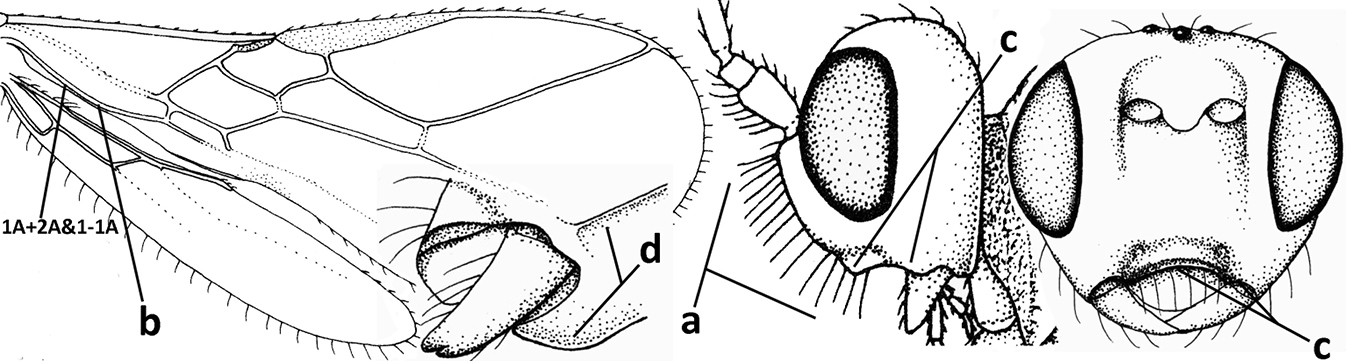
| 1 | Vein 2-SR of fore wing absent (a); first subdiscal cell of fore wing open apically (b); vein cu-a of hind wing absent (c); segments of maxillary palp usually shorter (d); [clypeus wide, short, and impressed; hind wing narrow] | 2 |
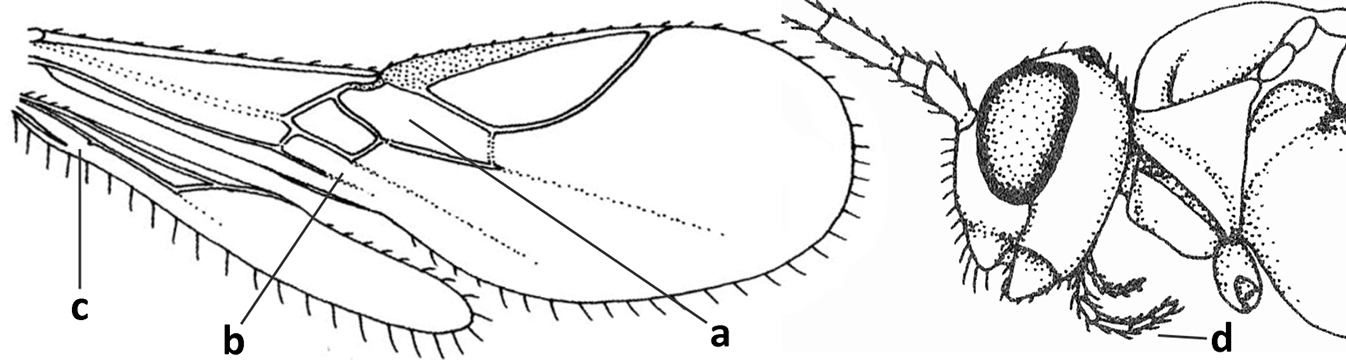
|
||
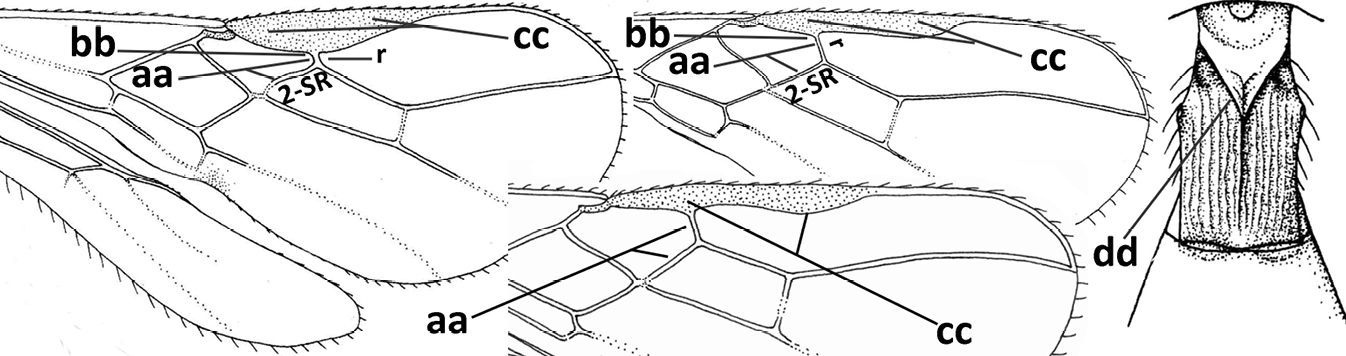
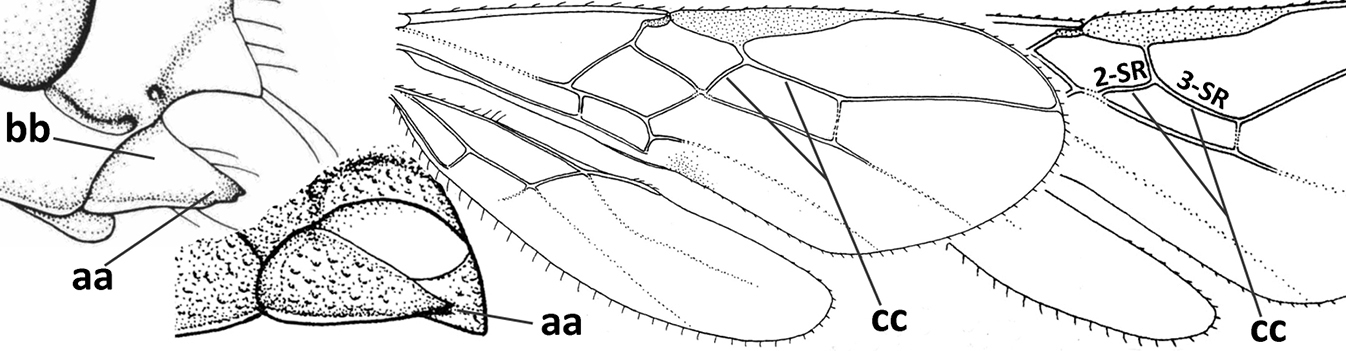
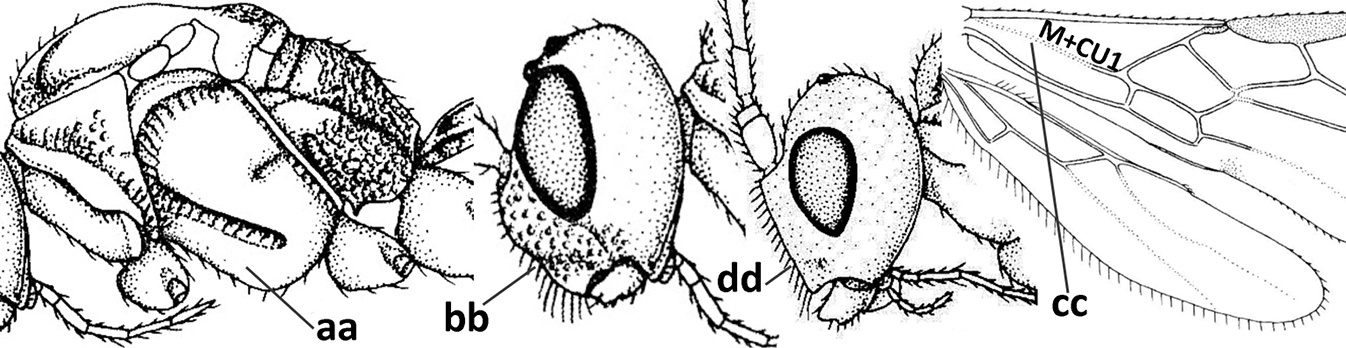
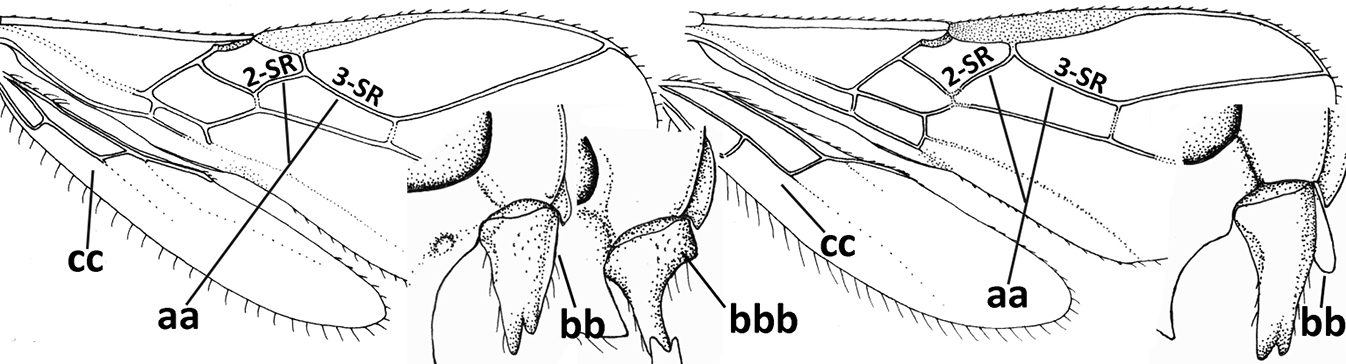
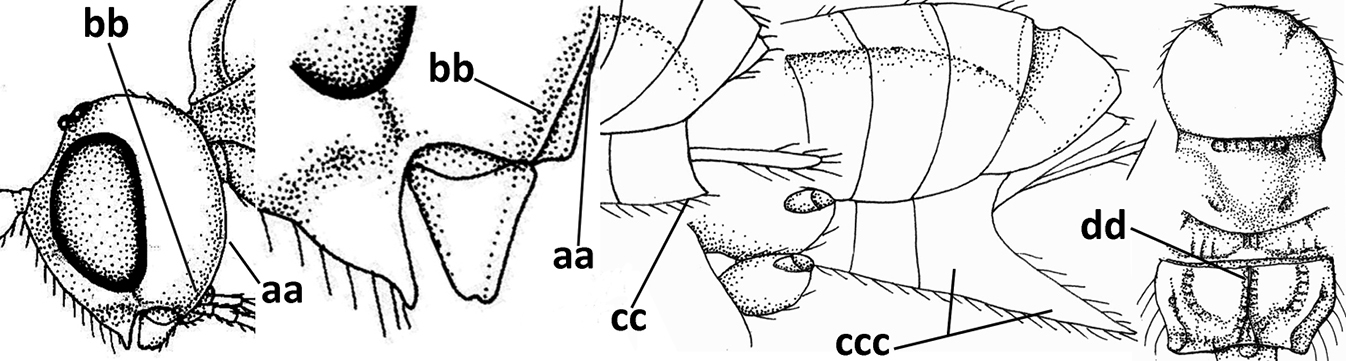
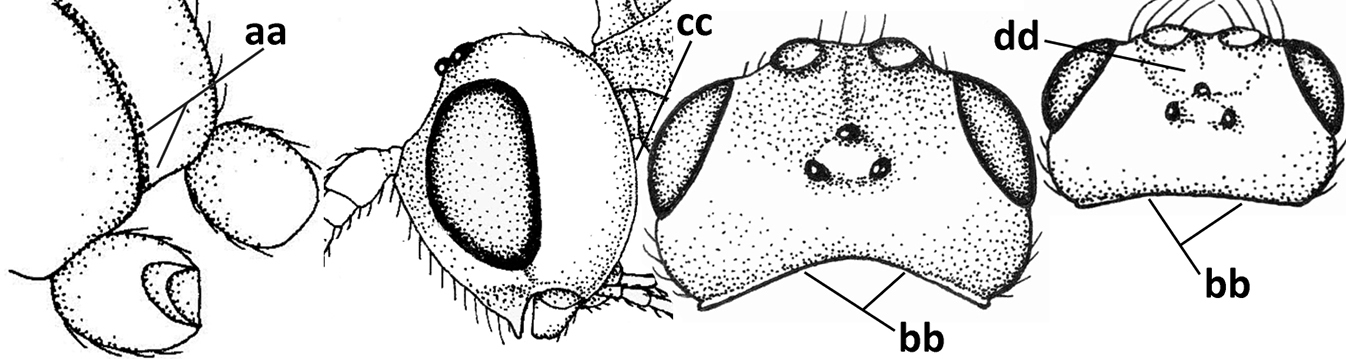
| – | Vein 2-SR of fore wing present (aa), rarely absent; first subdiscal cell of fore wing at least partly closed by vein 3-CU1 apically (bb); vein cu-a of hind wing nearly always present (cc); segments of maxillary palp often elongate (dd) | 3 |
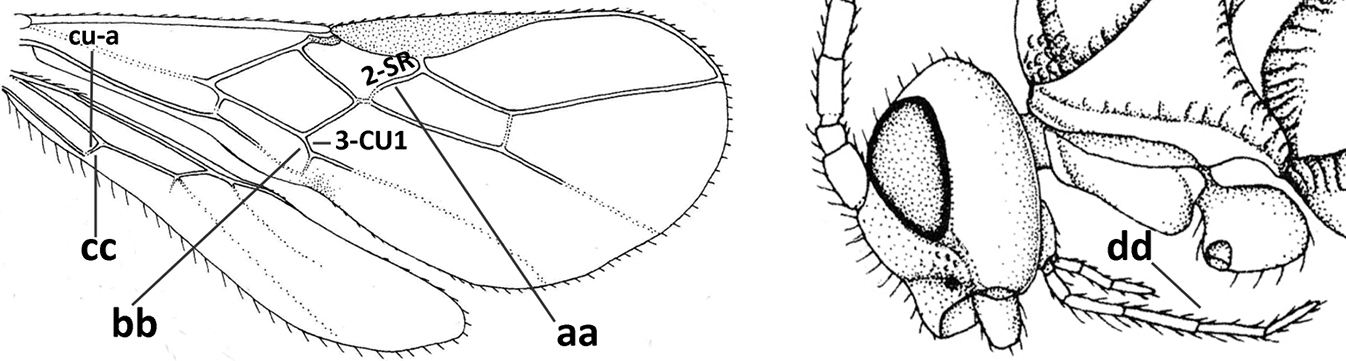
|
||
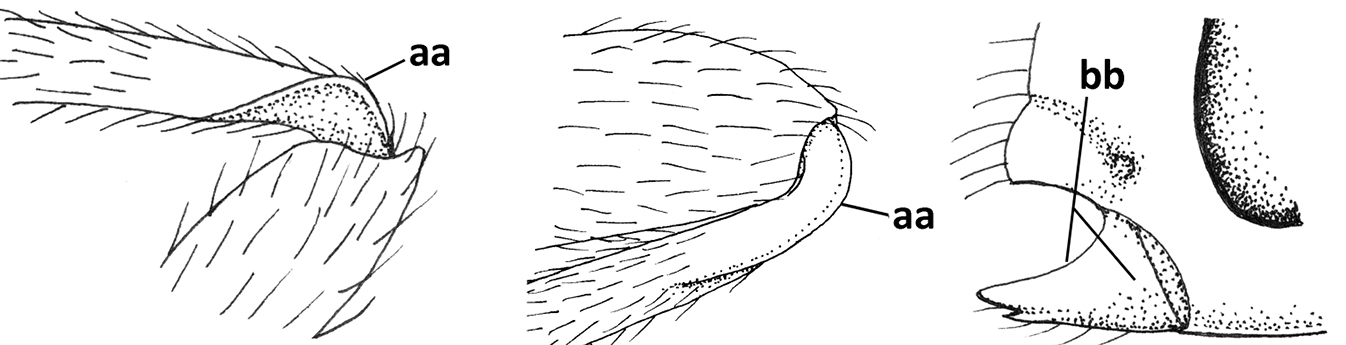
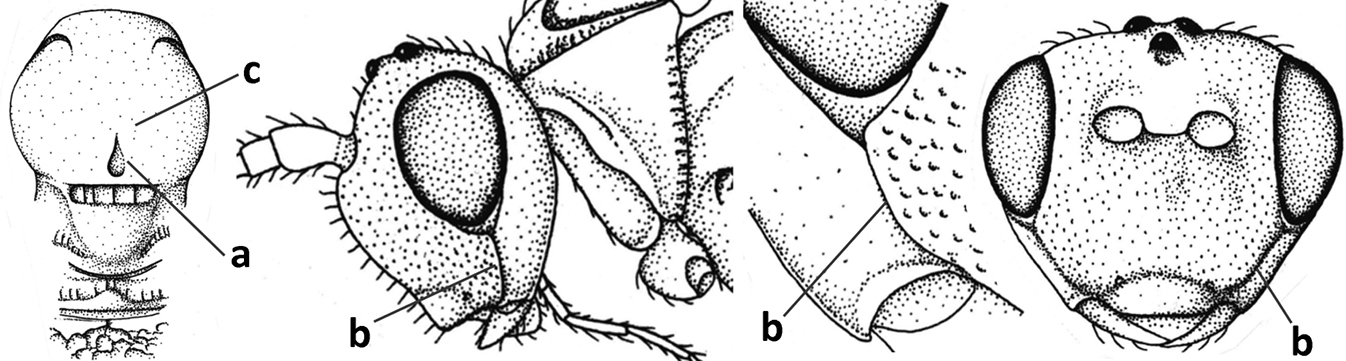
| 2 | Marginal cell of fore wing open apically (a) and long; veins m-cu (b) and r-m (c) of fore wing absent; occipital carina absent laterally (d); [metasomal tergites I and III more or less coriaceous or rugulose; tergites IV–VI largely retracted] | Indiopius Fischer, 1966 |
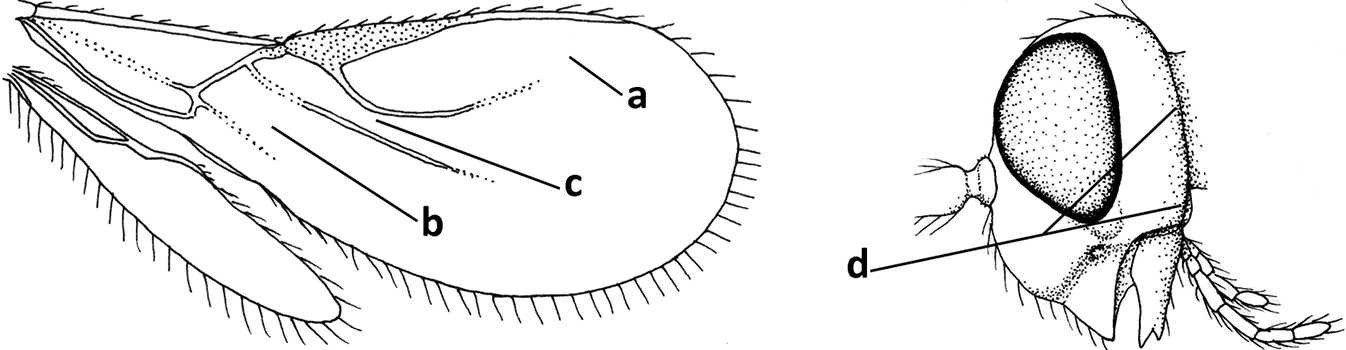
|
||
| – | Marginal cell of fore wing closed apically (aa) and shortened; veins m-cu (bb) and r-m (cc) of fore wing present; occipital carina present laterally (dd) | Pokomandya Fischer, 1959 |
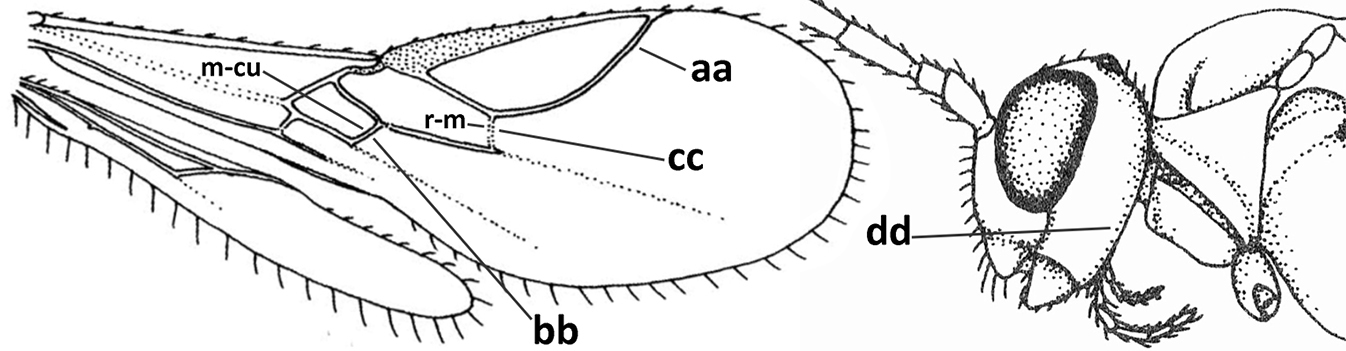
|
||
| 3 | Medio-laterally occipital carina near level of middle of eye strongly curved, resulting in an oblique part of carina or crest (a); vein 1-R1 of fore wing shorter than pterostigma (b); clypeus flat and short (c) | Hoplocrotaphus Telenga, 1950 |
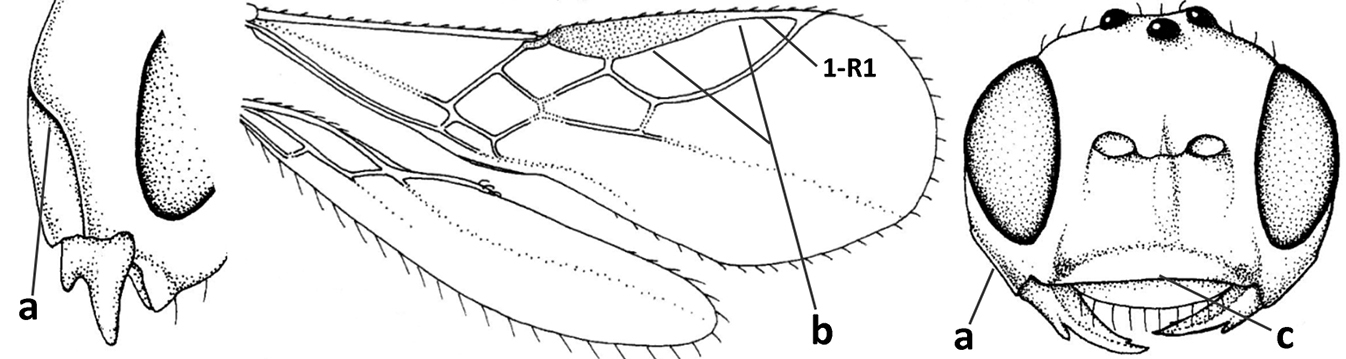
|
||
| – | Medio-laterally occipital carina straight or slightly curved (aa) or carina absent; vein 1-R1 of fore wing usually as long as or longer than pterostigma (bb); if shorter then clypeus more or less convex and longer (cc) | 4 |
|
||
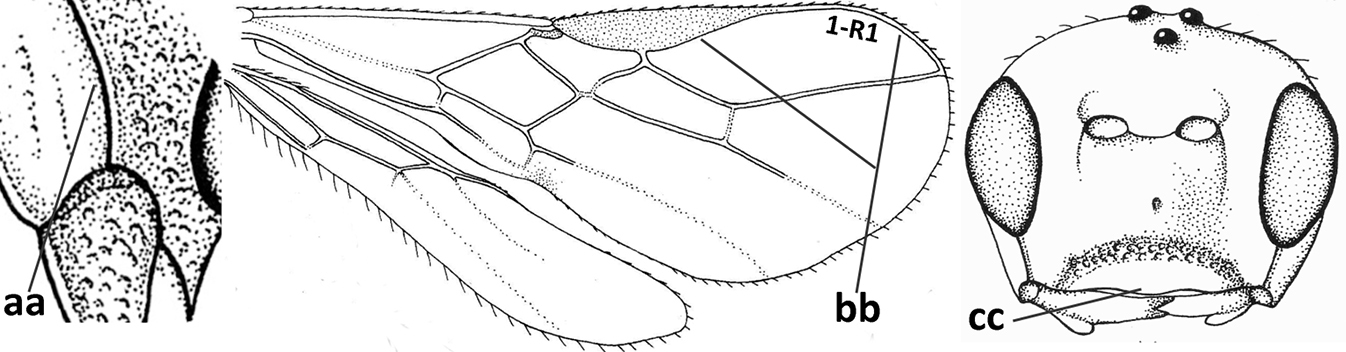
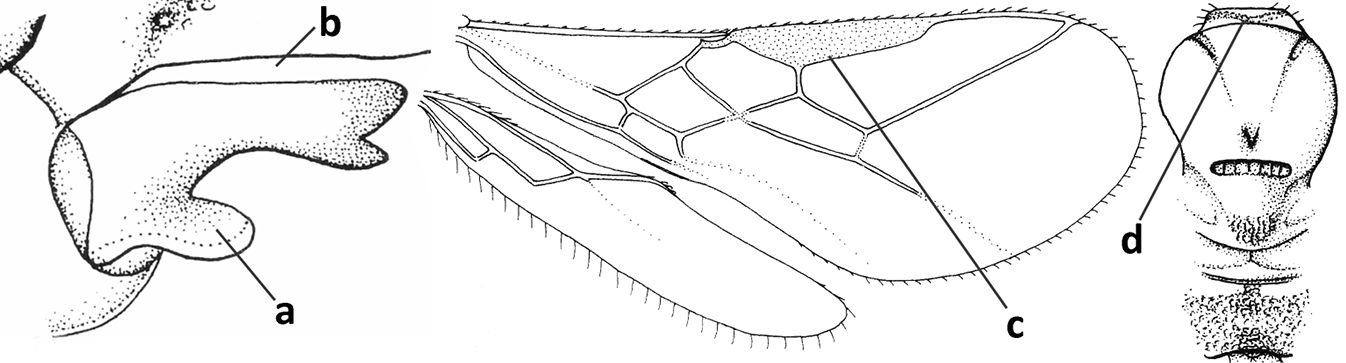
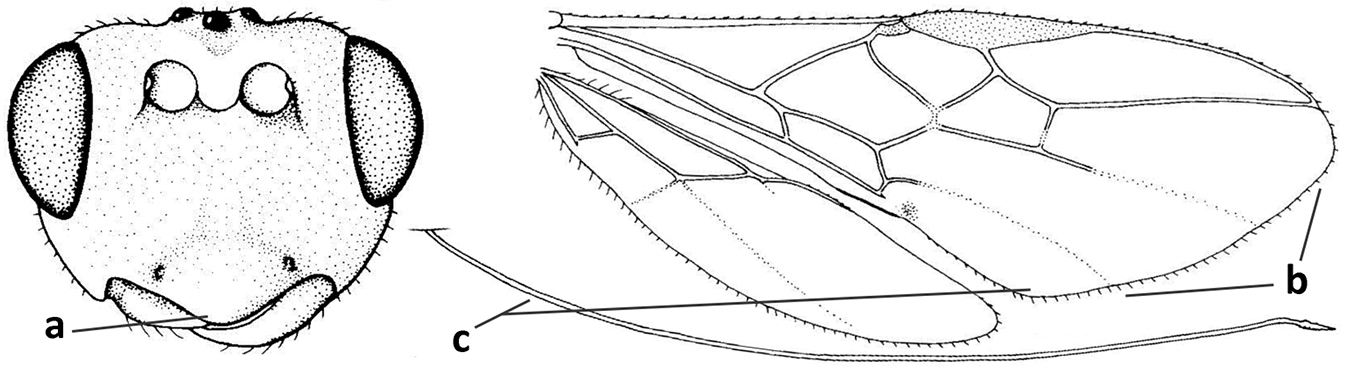
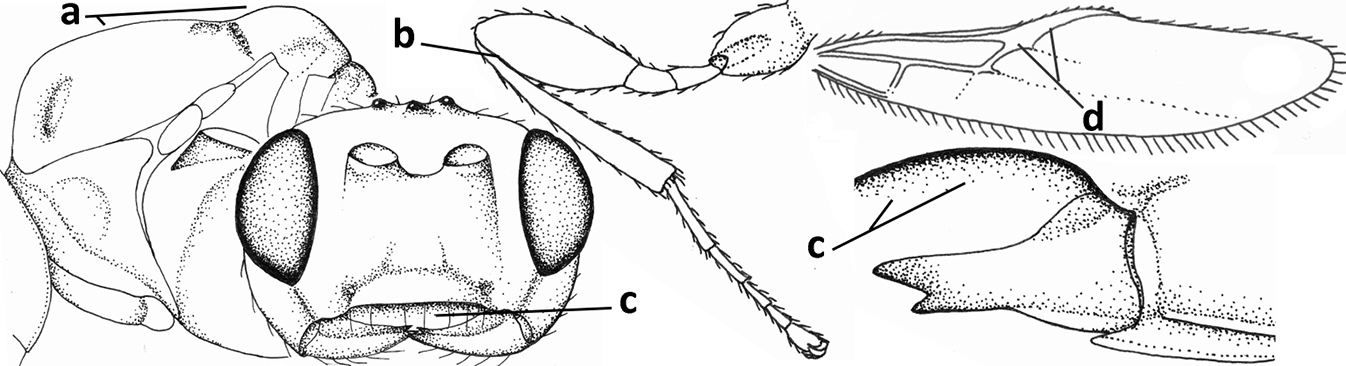
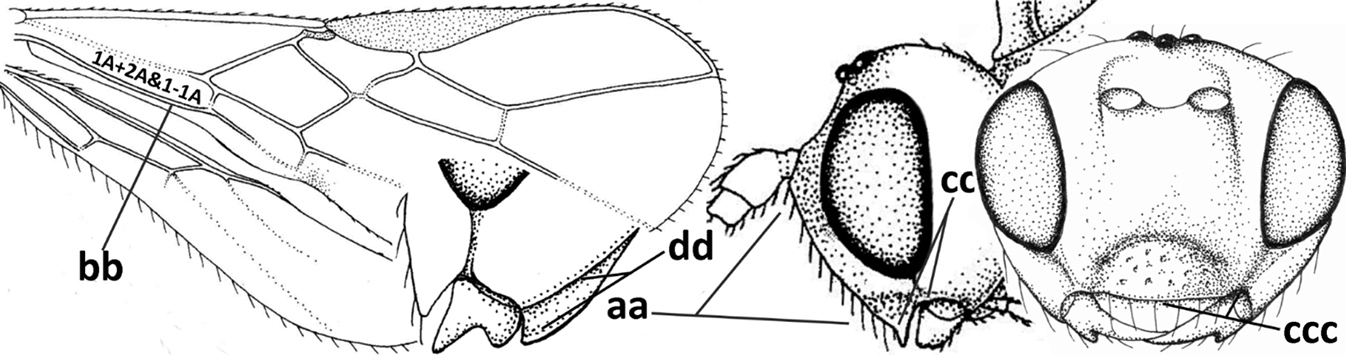
| 4 | Occipital carina completely absent (a) and vein CU1b of fore wing absent (b); convex labrum with long curved setae forming a comb ventrally (observe in oblique view and mandibles at least partly open: c); [body black; malar suture slightly longer than basal width of mandible; anterior tentorial pits close to apical margin of clypeus; ovipositor sheath shorter than apical height of metasoma] | Desmiostoma Foerster, 1863 |
|
||
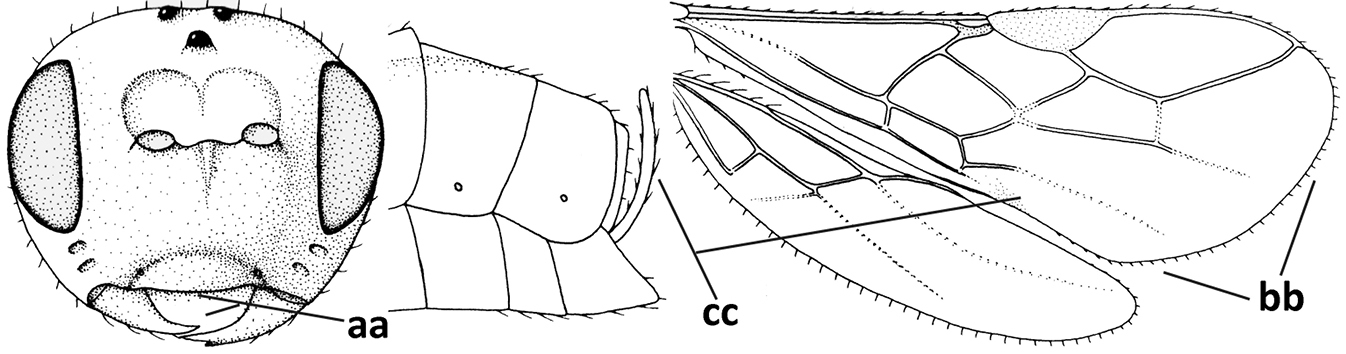
| – | Occipital carina present laterally (aa); if rarely absent then vein CU1b of fore wing present (bb); labrum flattened and without comb of setae ventrally (cc) | 5 |
|
||
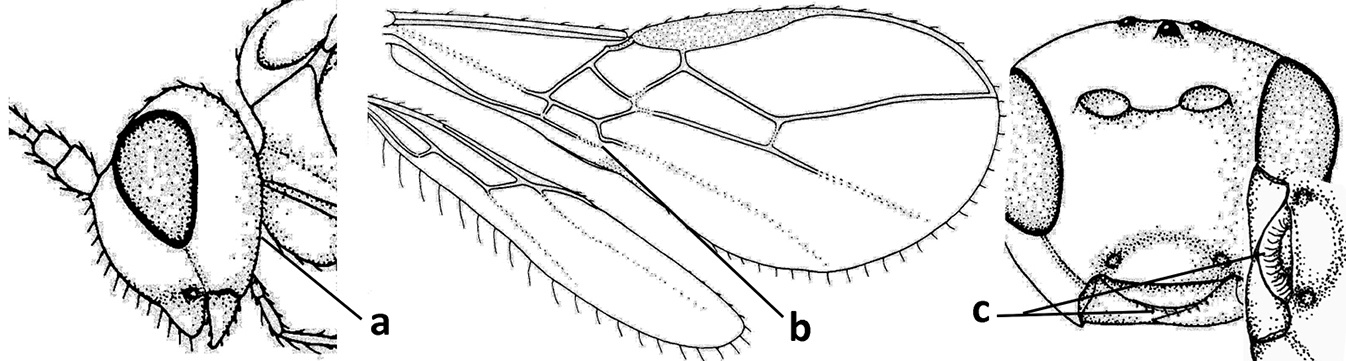
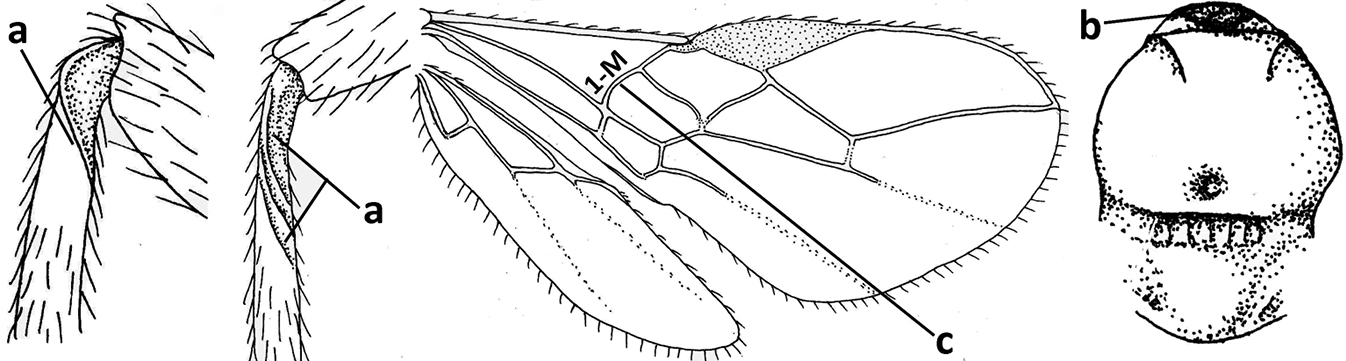
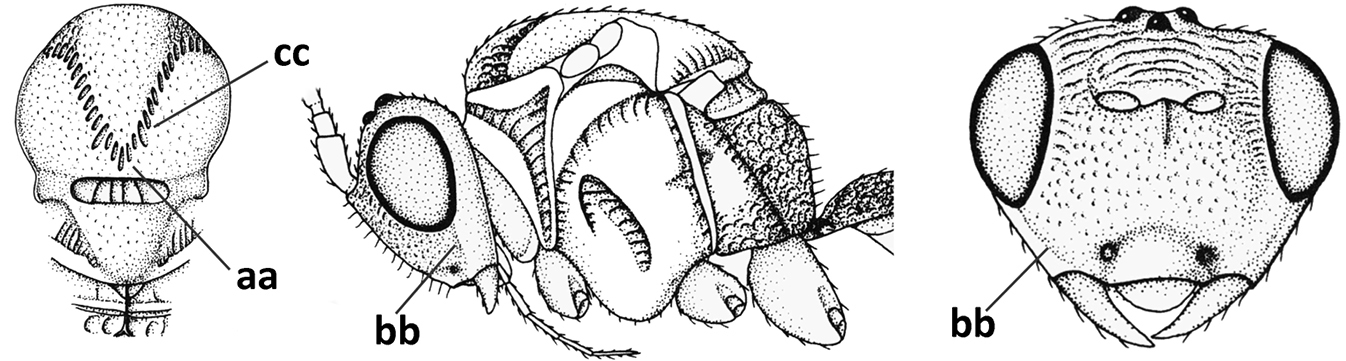
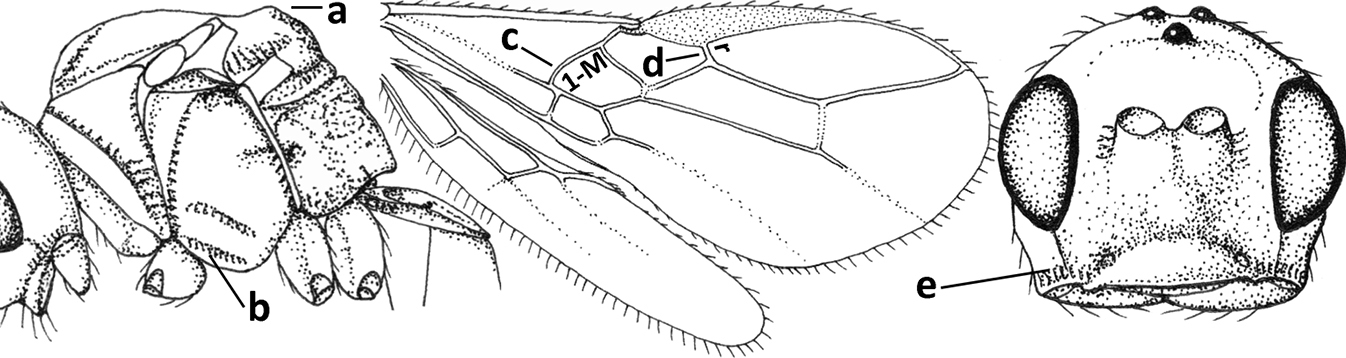
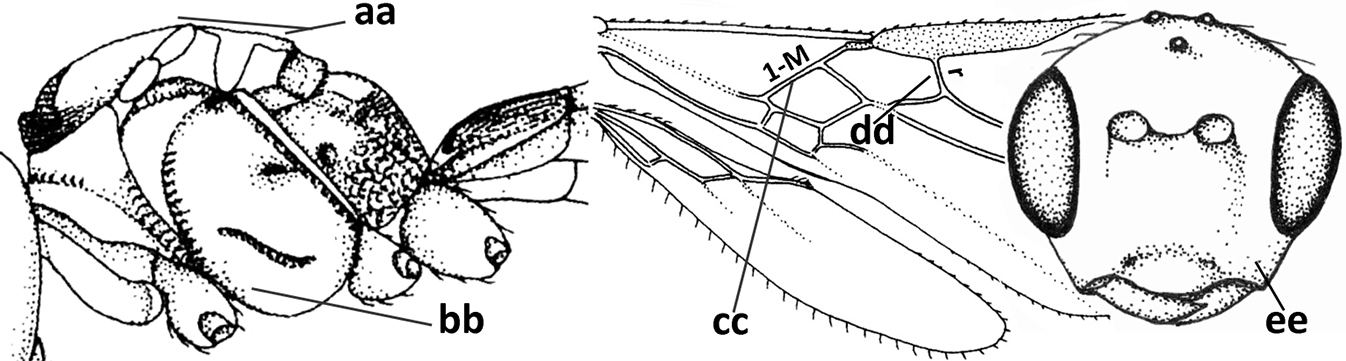
| 5 | Inner side of hind tibia with 1 carina or 3 or 4 carinulae baso-laterally in a glabrous area (a); pronope slit-like, round or elliptical, often large (occupying most of medio-dorsal part of pronotum) and deep (b); vein 1-M of fore wing more or less curved (c); [propodeum often coarsely (reticulate-)rugose and with more or less curved transverse carina dorso-laterally; dorsope absent or slightly impressed because of strongly developed dorsal carinae; hypostomal flange usually large below base of mandible] | 6 |
|
||
| – | Inner side of hind tibia without carina or carinulae baso-laterally and largely setose (aa); pronope usually round and smaller or pronotum only with shallow transverse groove (bb); vein 1-M of fore wing usually straight (cc) | 7 |
|
||
| 6 | Inner side of hind tibia with 3 or 4 fine carinulae (a; rarely 5 or 6 carinulae); mandible distinctly asymmetrical, more narrowed submedially (b) and widened ventro-basally, but less so in O. pactus) | Opiognathus Fischer, 1972 |
|
||
| – | Inner side of hind tibia with one rather lamelliform carina (aa); mandible nearly symmetrical, gradually narrowed submedially (bb) and usually slightly widened ventro-basally | Utetes Foerster, 1863 s.str. |
|
||
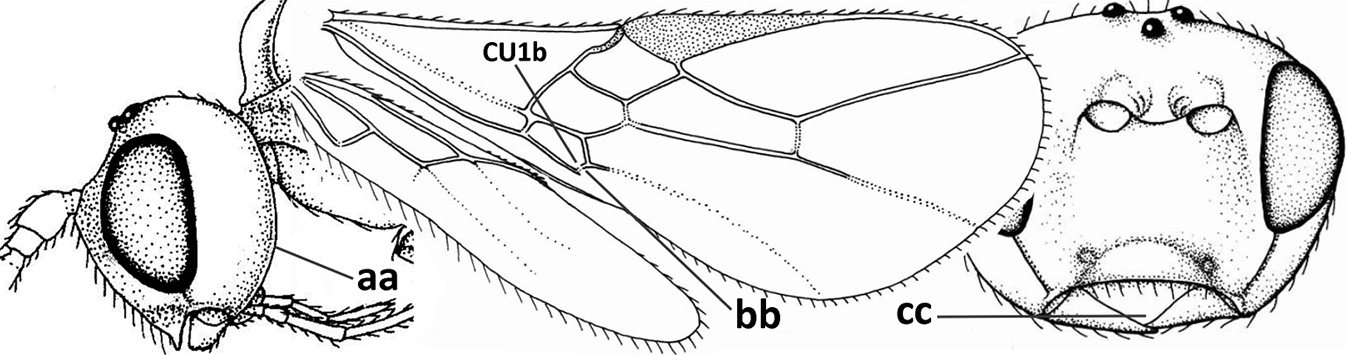
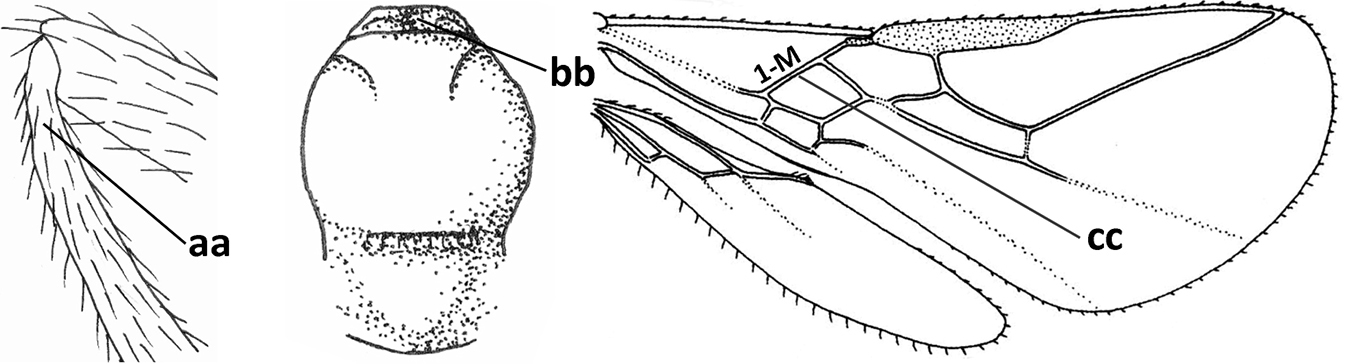
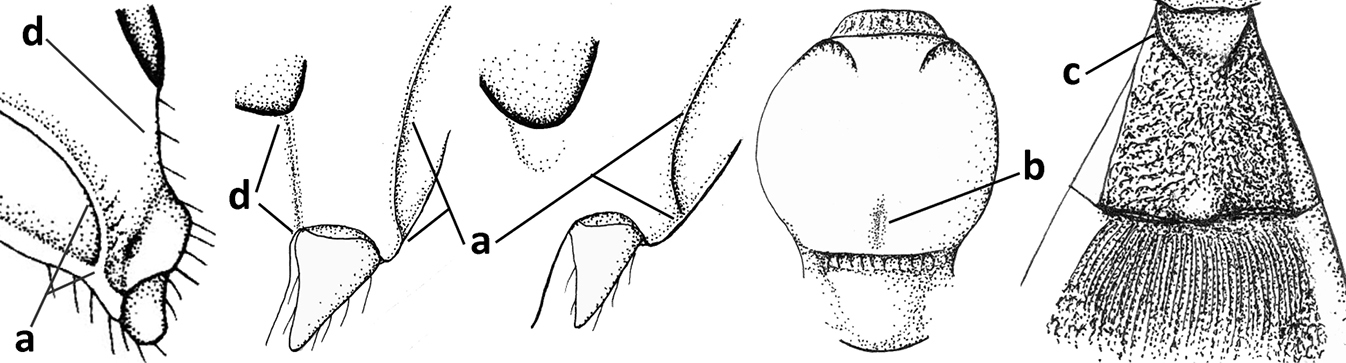
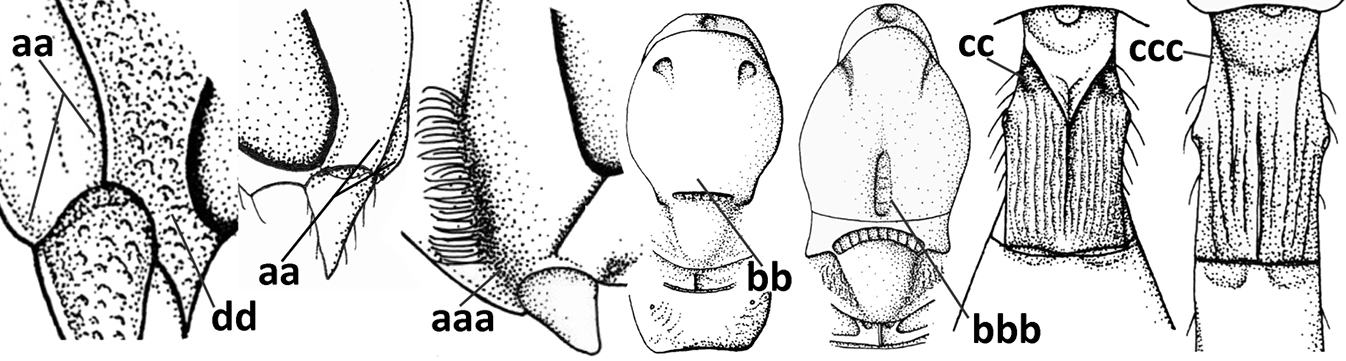
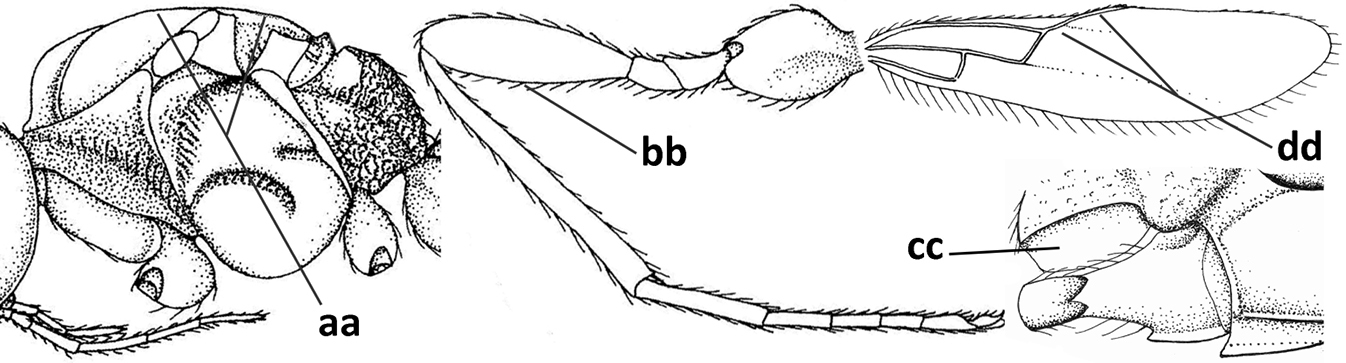
| 7 | Occipital carina above mandibular base curved and (just) meeting hypostomal carina (a); medio-posterior depression of mesoscutum present (b), only rarely absent or obsolescent; dorsope absent (c); malar space comparatively long (d); [if dorsope present and medio-posterior depression of mesoscutum absent, see Atormus van Achterberg] | Apodesmia Foerster, 1863 |
|
||
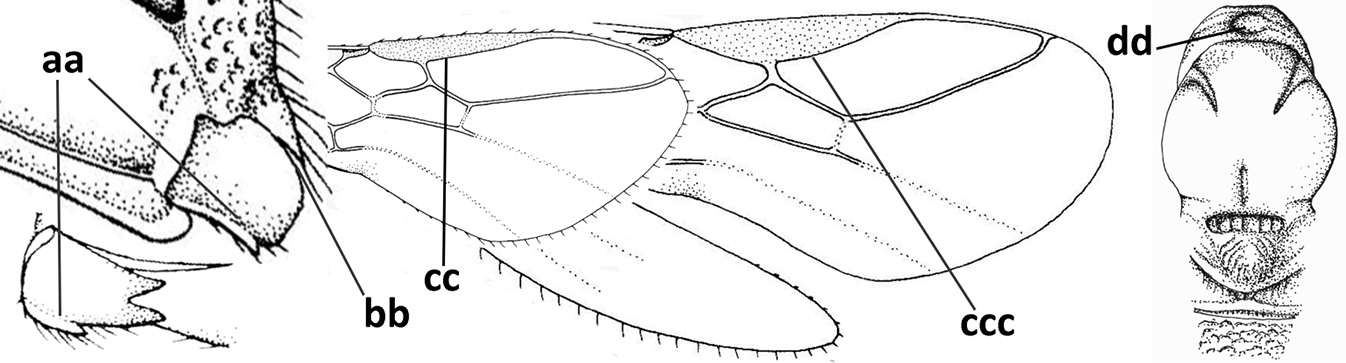
| – | Occipital carina not or slightly curved ventrally and remain removed from hypostomal carina (aa) or occipital carina absent laterally (aaa); medio-posterior depression of mesoscutum absent (bb) or present (bbb); dorsope present (cc) or absent (ccc); if absent then malar space usually short (dd) | 8 |
|
||
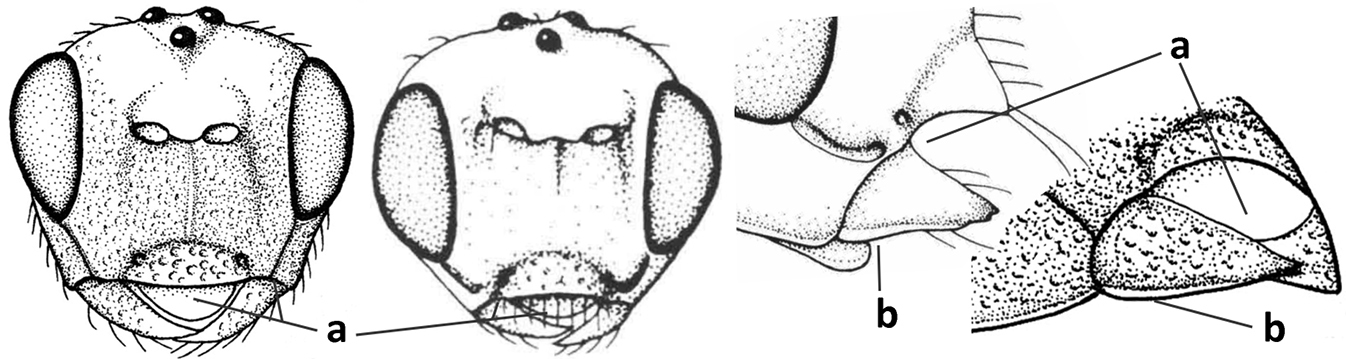
| 8 | Crenulate depression above eye present (a); prepectal carina more or less developed ventrally (b); vein SR1 of fore wing reduced apically, resulting in an open marginal cell (c); hind tibia and tarsus, and tarsal claws very slender (d); [medio-posteriorly scutellum with continuation of lateral elevated area; malar suture absent or obsolescent] | Ademon Haliday, 1833 |
|
||
| – | Crenulate depression above eye absent (aa); prepectal carina absent ventrally (bb); vein SR1 of fore wing completely sclerotised, reaching margin of wing and resulting in a closed marginal cell (cc); hind tibia and tarsus, and tarsal claws medium-sized (dd) | 9 |
|
||
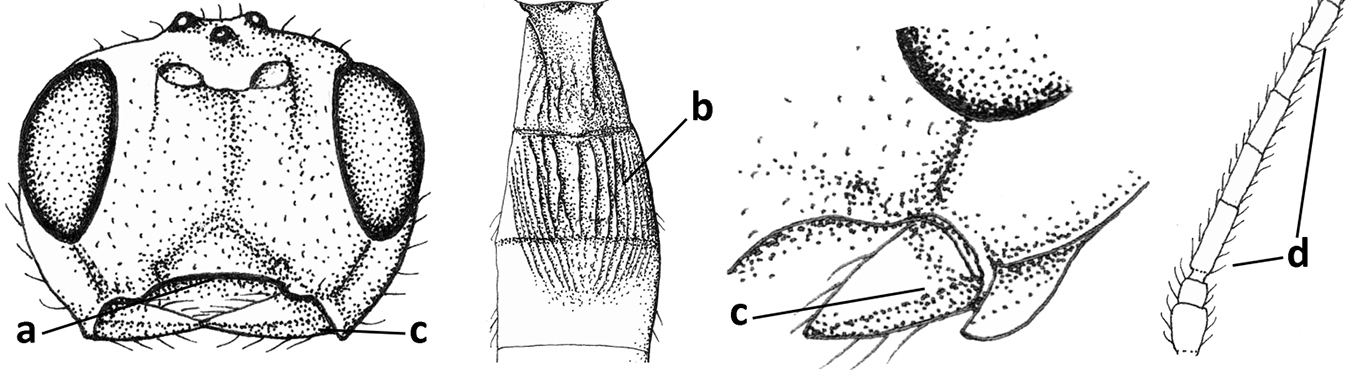
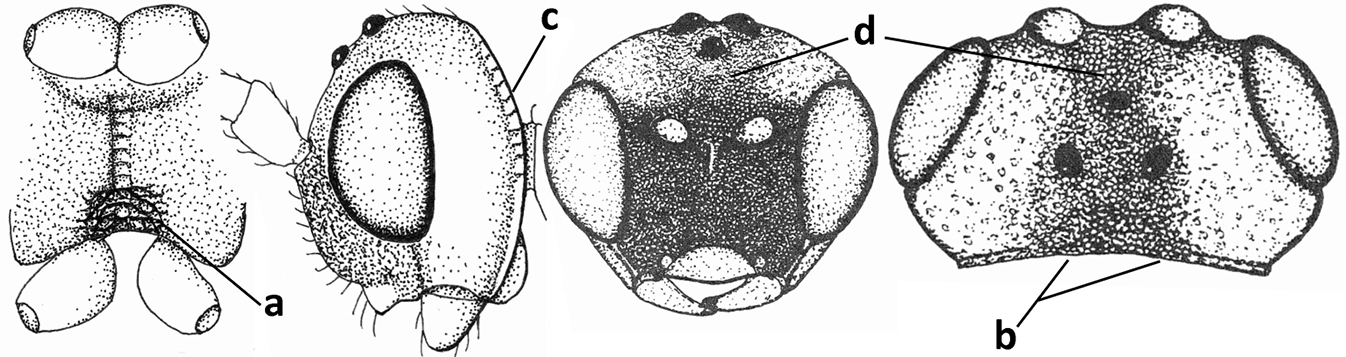
| 9 | Face with pair of tubercles below antennal sockets (a); epistomal suture with pair of large oblique and long pubescent depressions below facial tubercles (b); scape strongly compressed (c); scutellar sulcus narrow (d); [mandible with wide basal tooth] | Cephaloplites Szépligeti, 1897 |
|
||
| – | Face without tubercles (aa); epistomal suture without large depressions (bb); scape at most weakly compressed (cc); scutellar sulcus usually wider (dd) | 10 |
|
||
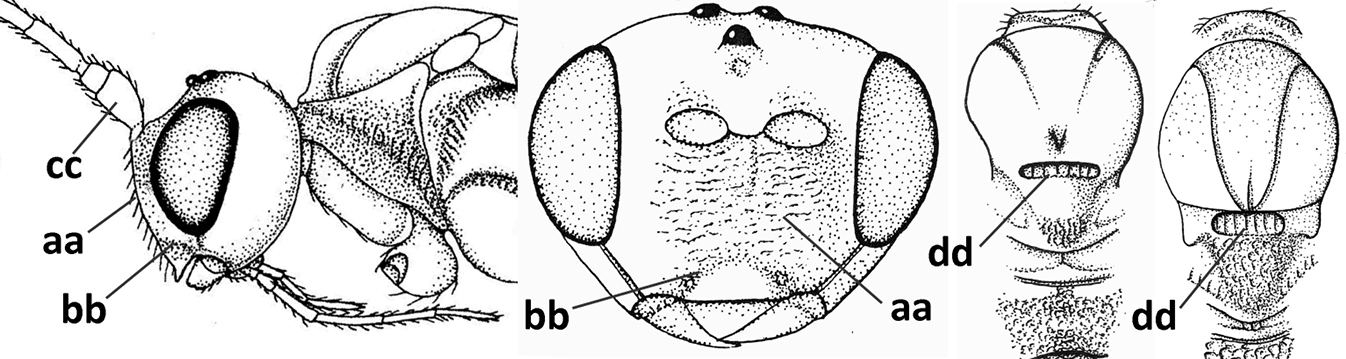
| 10 | Metasomal tergite II distinctly (1.3–1.9×) longer than tergite III and differentiated (a); tergite II bordered posteriorly by a curved second metasomal suture (b); vein 3-SR of fore wing approx. as long as vein 2-SR or slightly longer (c); tergite II longitudinally striate or rugose (d) | 11 |
|
||
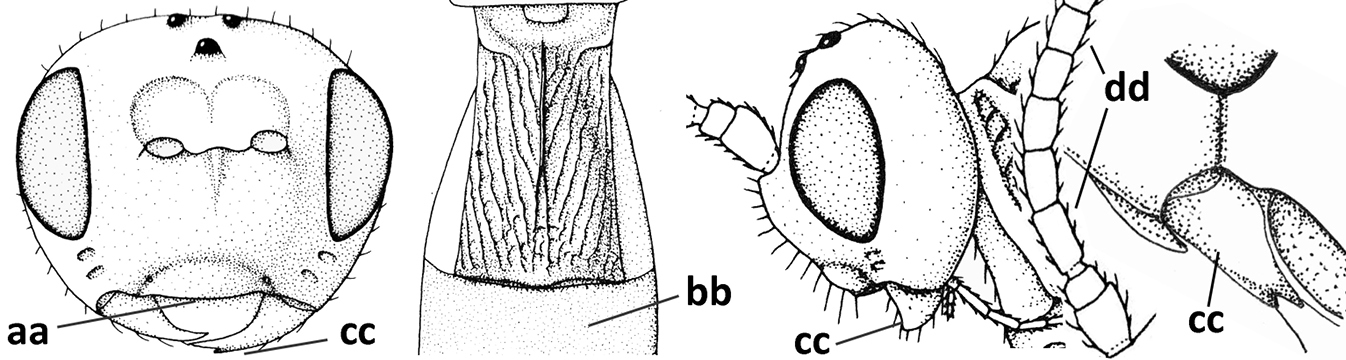
| – | Tergite II 0.7–1.1× as long as tergite III (aa), but frequently tergite II hardly or not differentiated; second metasomal suture nearly straight (bb) or absent (bbb); if metasoma shortened then vein 3-SR of fore wing distinctly longer than vein 2-SR (cc; Opius agnesae) and tergite II smooth or granulate (dd); [if metasoma shortened and propodeum with transverse carina subbasally, see Coleopioides van Achterberg & Li] | 13 |
|
||
| 11 | Around base of middle coxa with a circular carina (a); tergite III without sharp lateral crease (b); occipital carina present latero-dorsally and rather protruding in lateral view (c); head less elongate in anterior view (d); [dorsal carinae of tergite I united basally and medially forming a median carina; mesoscutum smooth laterally] | Bitomoides van Achterberg, 2004 |
|
||
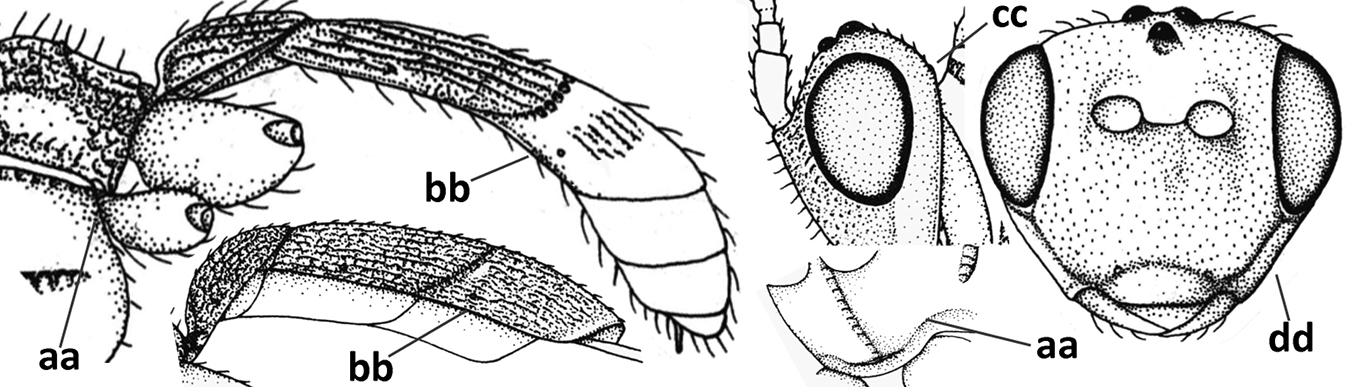
| – | Around base of middle coxa without a circular carina (aa); tergite III with a sharp lateral crease (bb); occipital carina absent latero-dorsally or if present usually less protruding in lateral view (cc); head more elongate in anterior view (dd); [mesoscutum finely crenulate laterally; dorsal carinae of tergite I usually separated basally and medially without a median carina] | 12 |
|
||
| 12 | Medio-posterior depression of mesoscutum present (a); malar suture complete and distinctly impressed (b); posterior 1/2 of notauli entirely or largely absent (c) | Orientopius Fischer, 1966 |
|
||
| – | Medio-posterior depression of mesoscutum absent (aa); malar suture absent or obsolescent (bb); posterior 1/2 of notauli complete (cc); [laterope distinct] | Coleopius Fischer, 1964 |
|
||
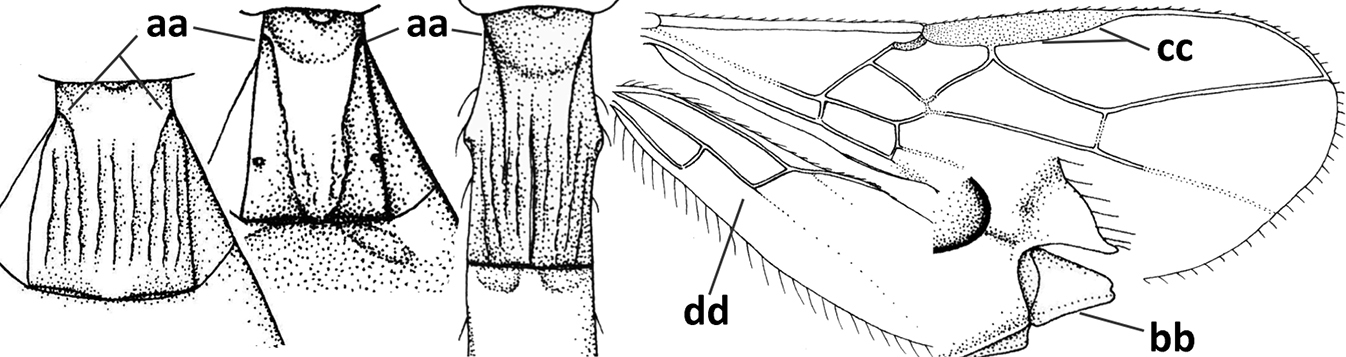
| 13 | Vein r of fore wing approx. as long as vein 2-SR (a), slightly curved, not angled with vein 3-SR and issued near base of pterostigma (b); basal third of pterostigma much narrower than apical third (c); dorsal carinae of metasomal tergite I largely obsolescent (d); [dorsope often minute or obsolescent; spiracles of tergite I situated near middle of notum; malar suture distinct] | Eurytenes Foerster, 1863 s.str. |
|
||
| – | Vein r of fore wing much shorter than vein 2-SR (aa), straight, more or less angled with vein 3-SR and issued distinctly removed from base of pterostigma (bb); basal 1/3 of pterostigma approx. as wide as apical 1/3 or slightly narrower (cc); dorsal carinae of tergite I usually distinctly developed (dd) | 14 |
|
||
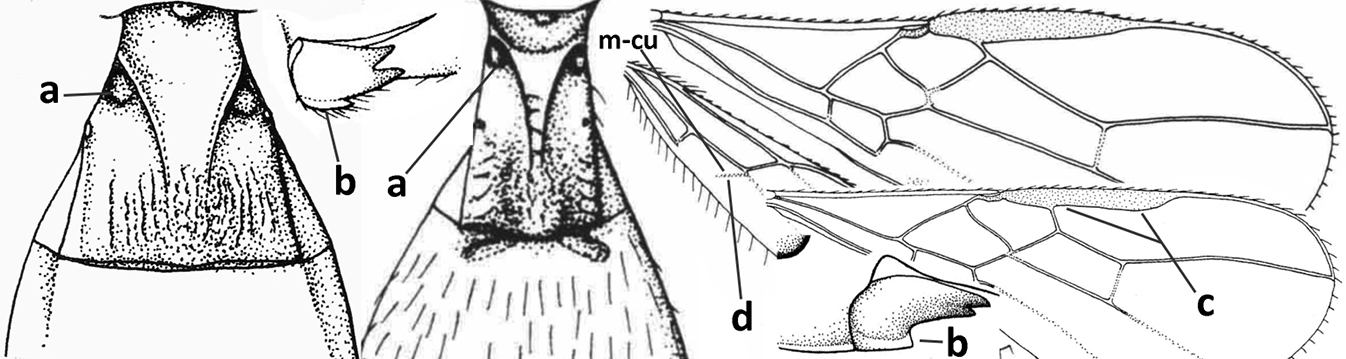
| 14 | Dorsope present (a); if with small dorsope then mandible with a short lamella ventro-basally (b) or pterostigma slightly widened subapically (c); vein m-cu of hind wing usually present, at least as a partly pigmented trace (d); [medio-posterior depression of mesoscutum present, rarely absent or obsolescent; antenna often longer than 1.3× length of fore wing] | 15 |
|
||
| – | Dorsope absent (aa); mandible usually without a short lamella ventro-basally or with a narrow carina along most of mandible ventrally (bb); pterostigma narrowed subapically (cc); vein m-cu of hind wing often absent (dd), but present in Diachasma and Bathystomus; [antenna usually less than 1.3× length of fore wing] | 21 |
|
||
| 15 | Mandible not or slightly twisted medially and its second tooth distinctly visible in lateral view (a), wide apically and its outer side convex (b); vein 3-SR of fore wing 1.3× as long as vein 2-SR or less (b), but 1.2–1.6× in Pseudosteres | 16 |
|
||
| – | Mandible more or less twisted medially and its second tooth hardly or not visible in lateral view (aa), triangular and its outer side often flattened or weakly convex (bb), but convex in Xynobius and Atormus; vein 3-SR of fore wing 1.4–2.5× longer than vein 2-SR (cc) | 18 |
|
||
| 16 | Mandibles asymmetrical, with a large ventro-basal tooth or lobe (a); hypoclypeal depression medium-sized to narrow (b), rarely absent; pterostigma wide elliptical or elongate triangular (c); pronope absent or nearly so (d); [vein 3-SR of fore wing 1.2–1.6× longer than vein 2-SR; vein m-cu of fore wing sometimes curved] | Pseudosteres gen. nov. |
|
||
| – | Mandible symmetrical, without ventro-basal tooth, at most with an acute basal lamella (aa); hypoclypeal depression absent (bb) or narrow; shape of pterostigma variable (cc), if comparatively wide elliptical or triangular (ccc) then usually pronope large (dd); [vein m-cu of fore wing straight] | 17 |
|
||
| 17 | Mesopleuron narrowed ventrally (a); epistomal suture absent (b); basal 1/2 of vein M+CU1 of fore wing entirely or largely sclerotised (c); clypeus with dense long setae, and convex (d); [mandible without distinct ventro-basal carina, but sometimes weakly developed] | Chilotrichia Foerster, 1863 |
|
||
| – | Mesopleuron wider and rounded ventrally (aa); epistomal suture present (bb); if shallow then basal 1/2 of vein M+CU1 of fore wing largely unsclerotised (cc); clypeus with less dense and shorter setae, if setae longer and denser then clypeus flattened (dd) | Biosteres Foerster, 1863 |
|
||
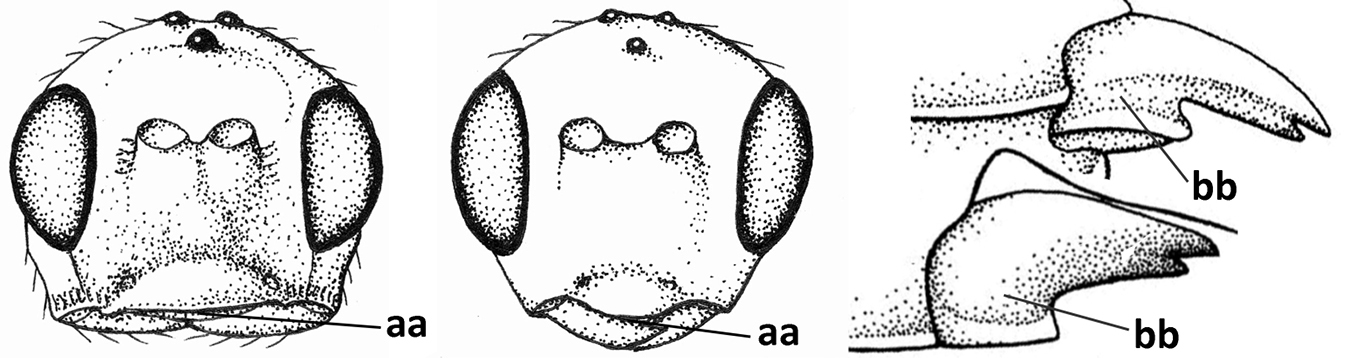
| 18 | Hypoclypeal depression present, large, and medially ventral margin of clypeus near upper level of condyles of mandibles (“subcyclostome condition”; a); mandible symmetrical basally (b) and outer side convex; [malar suture deep] | 19 |
|
||
| – | Hypoclypeal depression absent or narrow, and medially ventral margin of clypeus more or less below upper level of condyles of mandibles (“mouth closed” or nearly so; aa); mandible often widened ventro-basally (asymmetrical: bb) and outer side usually flattened or weakly convex | 20 |
|
||
| 19 | Propleuron without fine oblique carina subapically (a); medio-posterior depression of mesoscutum present (b); first subdiscal cell of fore wing subparallel-sided (c) | Xynobius Foerster, 1863 |
|
||
| – | Propleuron with (oblique or transverse) carina subapically (aa); medio-posterior depression of mesoscutum absent (bb); first subdiscal cell of fore wing widened distally (cc); [metasomal tergite III evenly setose, but sometimes sparsely so; hypostomal flange protruding ventrally] | Atormus van Achterberg, 1997 |
|
||
| 20 | Scutellum elevated above level of mesoscutum (a); below precoxal sulcus with a second finely sculptured sulcus (= sternaulus: b), but sometimes only medially superficially impressed; vein 1-M of fore wing curved (c) and vein r short (d); malar space crenulate ventrally (e); [vein m-cu of fore wing strongly converging posteriorly to vein 1-M or subparallel] | Biophthora Foerster, 1863 |
|
||
| – | Scutellum hardly or not elevated above mesoscutum (aa); sternaulus absent (bb), at most with one groove (= precoxal sulcus) present; vein 1-M of fore wing straight (cc) or nearly so and vein r medium-sized to long (dd); malar space smooth ventrally (ee) | Opiostomus Fischer, 1972 |
|
||
| 21 | Vein 3-SR of fore wing 0.8–1.3× vein 2-SR (a); if rarely 1.3–1.5× then basal 1/2 of mandible symmetrical (b); i.e., without a ventral carina or tooth) and vein m-cu of hind wing present (at least as a distinct partly pigmented trace: c); length of fore wing usually more than 3 mm | 22 |
|
||
| – | Vein 3-SR of fore wing 1.3× vein 2-SR or longer (aa); if 1.0–1.5× then basal 1/2 of mandible with a narrow ventral carina (bb) or baso-ventrally widened (asymmetrical: bbb) and/or vein m-cu of hind wing absent (cc) or obsolescent; length of fore wing usually less than 3 mm | 25 |
|
||
| 22 | Ventral margin of clypeus roundly protruding and without distinct hypoclypeal depression, at most with a narrow slit (a); fore wing pointed apically (b); ovipositor sheath long (c), usually as long as fore wing or longer; parasitoids of Tephritidae | Diachasmimorpha Viereck, 1913 |
|
||
| – | Ventral margin of clypeus almost straight or slightly concave, and with a distinct hypoclypeal depression (aa); fore wing broadly rounded apically (bb); ovipositor sheath short (cc), approx. as long as apical height of metasoma or concealed; parasitoids of other dipterous hosts | 23 |
|
||
| 23 | Precoxal sulcus absent or largely so (a); metasomal tergite I 2.0–2.5× longer than its apical width (b); metanotum with median carina posteriorly (c); malar suture complete and oblique (d); [metasomal tergites II and III of ♀ distinctly compressed; tergite II smooth] | Bathystomus Foerster, 1863 |
|
||
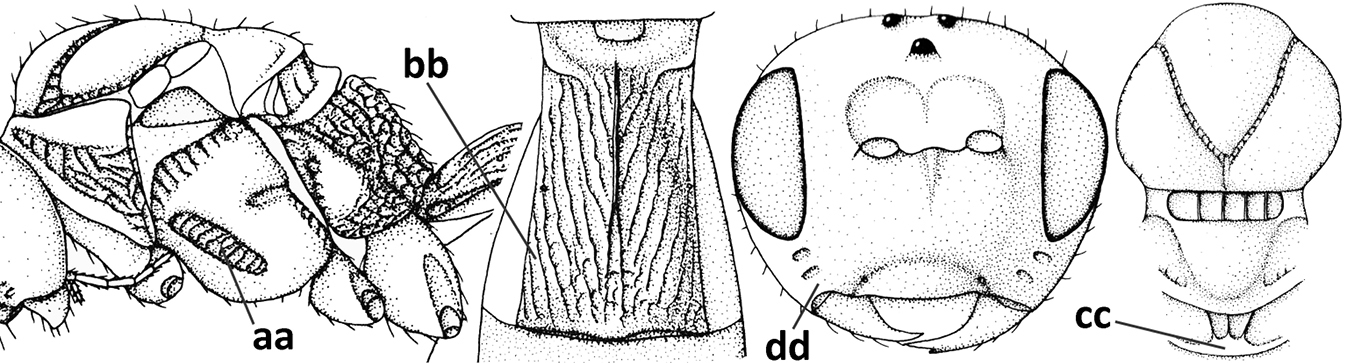
| – | Precoxal sulcus present and often wide on anterior 1/2 of mesopleuron (aa); length of tergite I 1.0–1.5× its apical width (bb); metanotum without median carina posteriorly or weakly developed (cc); malar suture absent or obsolescent (dd); [tergites II and III of ♀ not compressed] | 24 |
|
||
| 24 | Clypeus concave medio-ventrally (a); tergite II of ♀ with coarse striation (b); mandible strongly twisted basally (c); antennal segments slender (d); parasitoids of Psilidae; [mesosoma ~ 1.8× longer than high in lateral view; metanotum finely crenulate posteriorly; eyes diverging ventrally] | Atoreuteus Foerster, 1863 |
|
||
| – | Clypeus straight or protruding medio-ventrally (aa); tergite II of ♀ smooth or with very fine sculpture (bb); mandible not twisted basally (cc); antennal segments robust (dd); parasitoids of Anthomyiidae | Diachasma Foerster, 1863 |
|
||
| 25 | Ventral 1/2 of occiput with conspicuous and more or less curved setae (a); occipital carina widely absent ventrally (b); hypopygium of ♀ acute apically, nearly as long as apical height of metasoma or somewhat shorter (c) and propodeum largely smooth (d) | Cavopius gen. nov. |
|
||
| – | Ventral 1/2 of occiput glabrous (aa); occipital carina present (bb), at most narrowly absent ventrally; hypopygium of ♀ usually obtuse apically (cc), if acute then acute part longer than basal part (ccc) and/or anteriorly propodeum with medio-longitudinal carina (dd) and/or partly sculptured | 26 |
|
||
| 26 | Scutellum distinctly protruding above level of mesoscutum (a); hind femur very robust, 2–3× longer than wide (b); labrum slanted backwards, leaving a large space below clypeus (c); medio-anterior veins of hind wing of ♂ strongly widened (d); [hypopygium of ♀ 0.3–0.5× as long as metasoma and apically usually more or less protruding; dorsal face of propodeum strongly transverse; hind wing narrow] | Psyttoma van Achterberg & Li, 2012 |
|
||
| – | Scutellum at level of mesoscutum (aa); hind femur more slender, 3–5× longer than wide (bb); labrum normal, without large space below clypeus (cc); medio-anterior veins of hind wing of ♂ narrow (dd) | 27 |
|
||
| 27 | Hypopygium of ♀ more narrowed apically (a), acute, 0.3–0.5× as long as metasoma (b); pronope absent or small in dorsal view (c); lateral carina of mesoscutum absent in front of tegula (d); vein 1-M of fore wing straight or nearly so (e); propodeum often with medium-sized medio-longitudinal carina connected to narrow triangular areola (f), but largely obscured by sculpture in P. cyclogaster; [tergite II (if differentiated) distinctly shorter than tergite III or of similar length; hind wing and hypoclypeal depression wide; antenna of ♀ 1.3–1.6× as long as fore wing] | Psyttalia Walker, 1860 |
|
||
| – | Hypopygium of ♀ less narrowed apically (aa) and usually shorter than 0.3× length of metasoma (bb); pronope usually large to medium-sized (cc); if absent then in front of tegula with fine lateral carina of tegula (dd; sometimes area aciculate) or vein 1-M of fore wing curved (ee); medio-longitudinal carina of propodeum usually absent, if present then short and either no or wide areola (ff), rarely carina hardly developed (fff); [metasomal tergite II approx. as long as tergite III] | 28 |
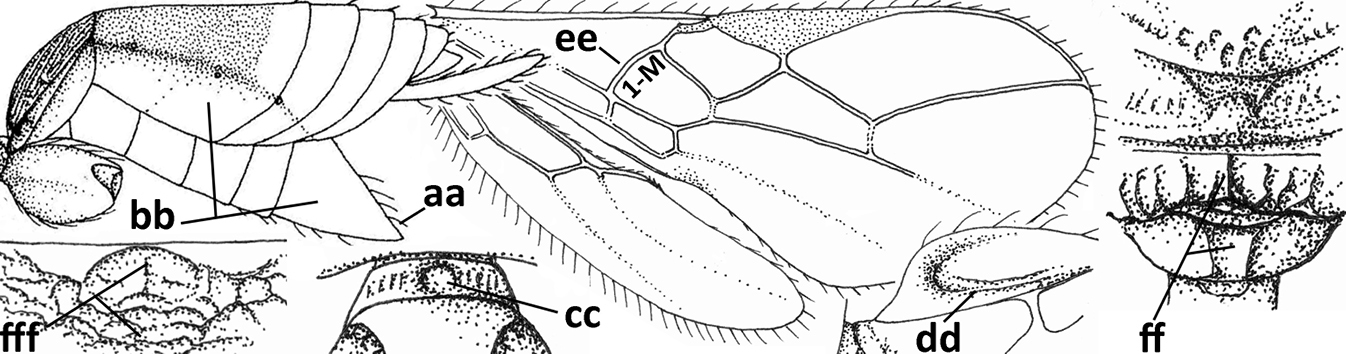
|
||
| 28 | Medio-posterior triangular area of mesosternum coarsely reticulate (a); occipital carina complete medio-dorsally or nearly so (b) and distinctly lamelliform dorso-laterally (c); frons distinctly granulate (d) | Neopius Gahan, 1917 |
|
||
| – | Mesosternum largely smooth medio-posteriorly (aa); occipital carina widely interrupted medio-dorsally (bb); if more or less complete dorsally then occipital carina narrow lateral (cc) and/or frons smooth (dd) | 29 |
|
||
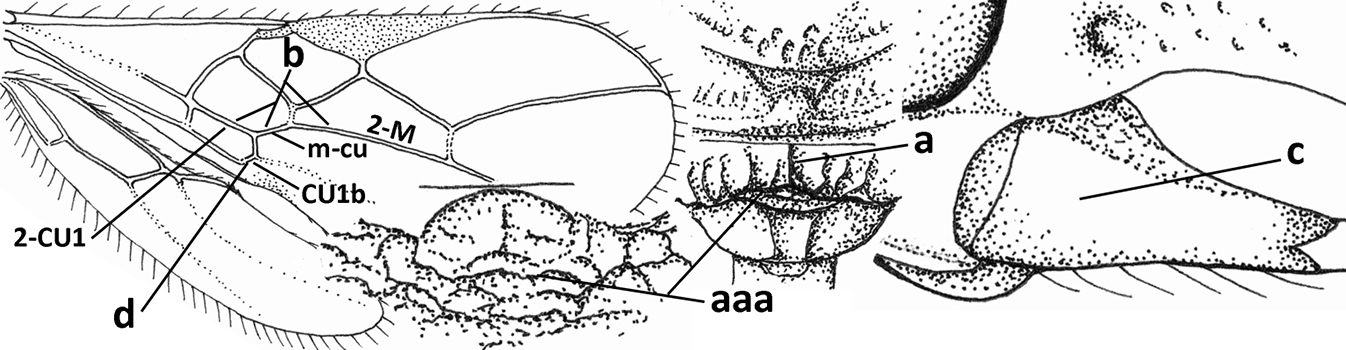
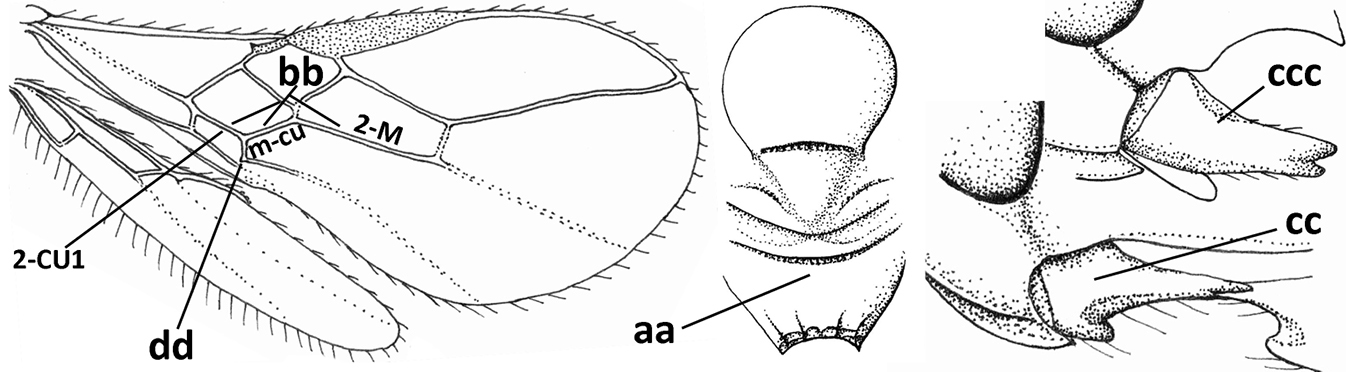
| 29 | Face conspicuously long and densely setose medio-dorsally (a); vein 1A+2A & 1-1A of fore wing distinctly bent and reaching posterior border of wing (b); thick ventral margin of clypeus distinctly concave medio-apically (c; medially distinctly above upper level of mandibular condyles); occipital carina fine and remaining far removed from hypostomal carina ventrally (d); [metasoma (except dark brown tergite I) orange-yellow] | Eutrichopsis Foerster, 1863 |
|
||
| – | Face without conspicuous setosity (aa); vein 1A+2A & 1-1A of fore wing straight or nearly so and remaining removed from posterior border of wing (bb); ventral margin of clypeus below upper level of mandibular condyles (cc) or approx. same level (ccc)) and usually thin; occipital carina usually more developed and closer to hypostomal carina ventrally (dd) | 30 |
|
||
| 30 | Medio-longitudinal carina of propodeum present anteriorly (a), if very short and reduced or obscured by coarse reticulation, then with transverse elements of rugosity subanteriorly (aaa); vein m-cu of fore wing gradually merging into 2-CU1 and sublinear with vein 2-M (b); mandible symmetrical or nearly so (c); vein CU1b of fore wing distinct and medium-sized to small (d); [hypoclypeal depression medium-sized to large; medio-posterior depression of mesoscutum absent and precoxal sulcus distinctly crenulate] | Rhogadopsis Brèthes, 1913 |
|
||
| – | Medio-longitudinal carina of propodeum absent and area smooth (aa), if rugose then without distinct transverse elements; vein m-cu of fore wing angled with veins 2-CU1 and vein 2-M (bb); shape of mandible varies from more or less asymmetrical and strongly narrowed medially (cc) to symmetrical and slightly narrowed medially (ccc); vein CU1b of fore wing short or absent (dd) | Opius Wesmael, 1835 s.l. |
|
||
Taxonomy
Cavopius gen. nov.
Type species
Opius (Agnopius) daghoides Zaykov & Fischer, 1983.
Etymology
From cavus (Latin for hollow) and the generic name Opius Wesmael, because of the long and curved setae make a kind of cave at the back of the head (Fig.
Diagnosis
Antenna with 26–37 segments and 1.1–1.2× as long as fore wing (latter unknown of C. daghestanicus); ventral half of occiput with medium-sized to large area of long conspicuous and usually curved setae (Figs

Distribution
Palaearctic: three species.
Key to species of the genus Cavopius gen. nov
| 1 | Setose part of ovipositor sheath 0.6–0.7× as long as fore wing (Fig. |
C. daghoides (Zaykov & Fischer, 1983) |
| – | Setose part of ovipositor sheath 0.9–1.2× as long as fore wing (Fig. |
2 |
| 2 | Setose part of ovipositor sheath ~ 1.2× as long as fore wing (Fig. |
C. depressorius sp. nov. |
| – | Setose part of ovipositor sheath ~ 0.9× as long as fore wing; tergites IV–VI flat and evenly sclerotised; vein 1-M of fore wing 4× as long as vein 1-SR (Fig. |
C. daghestanicus (Telenga, 1950) |
Cavopius daghestanicus , comb. nov.
Opius daghestanicus
Telenga, 1950: 306;
Opius (Misophthora) daghestanicus;
Opius (Agnopius) daghestanicus;
Opius (Allotypus) daghestanicus;
Type material
Holotype
, ♀ (
Diagnosis
Antenna of ♀ with approximately 28 segments; face laterally black or dark brown; ventral half of occiput less conspicuously setose (Fig.

Biology
Unknown.
Distribution
Moldova, Russia (N. Caucasus, Siberia).
Notes
Photographs of the damaged holotype were very kindly supplied by Konstantin Samartsev (
Cavopius daghoides , comb. nov.
Opius (Agnopius) daghoides Zaykov & Fischer, 1983: 41–44.
Phaedrotoma daghoides;
Type material
Holotype
, ♀ (
Diagnosis
Antenna with 26 segments (♀); face laterally yellowish brown; curved setae of ventral half of occiput conspicuous (Figs

Cavopius daghoides (Zaykov & Fischer), comb. nov., holotype, ♀, Bulgaria (Konush) 11 wings 12 head anterior 13 ventral half of occiput and mandible lateral 14 mesosoma dorsal 15 hind leg 16 head dorsal 17 ventral half of occiput latero-posterior 18 outer hind tarsal claw 19 apex of antenna 20 base of antenna 21 antenna 22 habitus lateral 23 metasomal tergite I dorsal. Scale bar: 1.0× (11, 12, 14–16, 21, 22); 1.5× (23); 1.6× (13, 17); 2.5× (18–20).
Biology
Unknown.
Distribution
Bulgaria, Spain.
Cavopius depressorius sp. nov.
Type material
Holotype
, ♀ (
Diagnosis
Antenna with 37 segments (♀); curved setae of ventral half of occiput conspicuous (Figs

Description
Holotype, ♀, length of body 3.2 mm, of fore wing 3.4 mm.
Head. Antenna with 37 segments and 1.2× as long as fore wing; third segment 1.2× longer than fourth segment, length of third, fourth and penultimate segments 1.5×, 1.2× and 1.4× their width, respectively (Figs
Mesosoma. Length of mesosoma 1.2× its height (Fig.
Wings. Fore wing (Fig.
Legs. Hind femur, tibia and basitarsus 5.3×, 8.7× and 4.8× as long as wide, respectively (Fig.
Metasoma. Tergite I 1.1× as long as wide apically and slightly widened apically, its surface convex medially and largely rugulose-punctate, dorsal carinae rather weakly developed and nearly up to apex of tergite (Fig.
Colour. Black; temple chestnut brown; mandible largely, clypeus, malar space largely, palpi and legs pale yellow; tergite II and following tergites brownish yellow, but membranous parts more or less brown; antenna (but ventro-basally yellowish) dark brown; pterostigma and veins brown; ovipositor sheath dark brown; wing membrane subhyaline (Fig.
Distribution
Korea.
Biology
Unknown.
Etymology
Named after the broadly depressed metasomal tergites IV–VI (Figs
Pseudosteres gen. nov.
Type species
Biosteres adanaensis Fischer & Beyarslan, 2005.
Etymology
From “pseudos” (Greek for “fallacy”) and the generic name Biosteres Foerster, because it is similar to Biosteres, but differs considerably as indicated below. Gender: masculine.
Diagnosis
Hypoclypeal depression usually medium-sized, and medially ventral margin of clypeus above upper level of condyles of mandibles, but depression absent in P. riphaeus and narrow in P. adanaensis (Fig.

Pseudosteres adanaensis (Fischer & Beyarslan), comb. nov., holotype, ♀ Turkey (Adana-Balcali) 34 wings 35 head anterior 36 mesosoma dorsal 37 hind leg 38 base of antenna 39 head dorsal 40 mandible lateral 41 metasomal tergite I dorsal 42 outer hind claw lateral 43 antenna 44 habitus lateral. Scale bar: 1.0× (34, 36, 37, 43, 44); 1.3× (35, 39); 1.5× (41); 2.5× (38, 40, 42).
Distribution
Palaearctic: five species.
Notes
Most species are similar to the genus Biosteres Foerster, 1863, but differ by having a large ventro-basal tooth or lobe and in part of species also by the presence of a medium-sized hypoclypeal depression. Members of Opiostomus Fischer have also the mandibles basally widened and the dorsope developed, but the mandible is distinctly twisted medially, the second submarginal is much longer and the scutellum is smooth posteriorly (but punctate in O. leptostigma (Wesmael)).
Key to species of the genus Pseudosteres gen. nov
| 1 | Antenna of ♀ with ~ 44 segments; vein r of fore wing emitted medially from pterostigma; hypoclypeal depression absent; [vein 3-SR of fore wing ~ 1.2× vein 2-SR; precoxal sulcus distinctly sculptured; scutellum convex and finely rugulose; vein r from middle of pterostigma; mesoscutum with pair of brownish stripes; metasoma (except tergite I) reddish brown; head densely setose dorsally; notauli absent on disc; mesoscutum punctate medially; propodeum coriaceous-rugulose]; Iran, Russia | P. riphaeus (Tobias, 1986) |
| – | Antenna of ♀ with 20–33 segments; vein r of fore wing emitted before middle from pterostigma (Fig. |
2 |
| 2 | Antenna of ♀ with ~ 33 segments (of ♂ with 32 (according to label of holotype, but in description 35) segments; length of body 2.5–3.0 mm and of fore wing ~ 3.0 mm; vein 1-R1 almost reaching apex of fore wing; vein m-cu of fore wing curved; [vein 3-SR of fore wing ~ 1.6× longer than vein 2-SR; ventro-basal lobe of mandible obtuse and distinctly protruding outwards; medio-posterior depression of mesoscutum large; precoxal sulcus distinctly crenulate; clypeus 5× wider than high; orbita of head largely yellow; length of hind femur (♂) ~ 3× longer than wide; vein m-cu of fore wing antefurcal; setose part of ovipositor sheath as long as metasomal tergite I]; England, Georgia, Greece, Ukraine (Crimea), and Asian part of Turkey | P. christenseni (Papp, 1982) |
| – | Antenna of ♀ with 20–25 segments (♂ unknown); length of body 1.5–2.1 mm and of fore wing 1.7–2.3 mm; vein 1-R1 of fore wing remaining distinctly removed from apex of wing (Fig. |
3 |
| 3 | Clypeus slightly sinuate ventrally, largely strongly shiny and comparatively weakly widened medially (Fig. |
P. adanaensis (Fischer & Beyarslan, 2005) |
| – | Clypeus straight ventrally, weakly shiny and distinctly widened medially; vein m-cu of fore wing antefurcal; medio-posterior depression of mesoscutum narrow elliptical; scutellum orange brown or chestnut brown | 4 |
| 4 | Vein SR1 of fore wing ~ 2.5× as long as vein 3-SR; lateral lobes of mesoscutum largely yellowish brown; [tergite I medio-posteriorly weakly striate; scutellar sulcus with distinct carinae; mesopleuron dorsally, mesoscutum, prothorax yellowish brown and metasomal tergite I black]; Ukraine | P. arenaceus (Jakimavičius, 1986) |
| – | Vein SR1 of fore wing ~ 3.5× as long as vein 3-SR; lateral lobes of mesoscutum black; [tergite I medio-posteriorly striate; scutellar sulcus crenulate; mesopleuron largely (except dorsally) and tergite I blackish or dark brown]; Asian Turkey | P. pseudarenaceus (Fischer & Beyarslan, 2005) |
Pseudosteres adanaensis , comb. nov.
Biosteres (Biosteres) adanaensis
Fischer & Beyarslan, 2005: 380–382;
Type material
Holotype
, ♀ (
Diagnosis
Antenna of ♀ with > 25 segments (♂ unknown); hypoclypeal depression narrow; clypeus slightly sinuate ventrally, largely strongly shiny and comparatively weakly widened medially (Fig.
Biology
Unknown.
Distribution
Turkey (Asian part).
Pseudosteres arenaceus , comb. nov.
Opius (Allotypus) arenaceus
Jakimavičius (in Tobias & Jakimavičius), 1986: 63;
Biology
Unknown.
Distribution
Ukraine.
Notes
According to
Pseudosteres christenseni , comb. nov.
Opius (Xynobius) christenseni
Papp, 1982: 185;
Type material
Holotype , ♂ (MTMA), “Greece, Peloponnese, Monemvasia”, “15. iv. 1978, J. Papp”, “Holotypus ♂ Opius (Xynobius) christenseni sp. n., Papp, J., 1980% / ant. 32-art., “Hym. Typ. No. 2844, Museum Budapest”.
Biology
Unknown.
Distribution
England, Greece, Ukraine, Georgia, and Asian Turkey.
Notes
As indicated on the label the holotype has 32 antennal segments, but according to the original description and the redescription the holotype male should have 35 antennal segments.
Pseudosteres pseudarenaceus , comb. nov.
Opius (Allotypus) pseudarenaceus
Fischer & Beyarslan, 2005: 407–409;
Type material
Holotype
, ♀ (
Biology
Unknown.
Distribution
Turkey (Asian part).
Notes
If Opius arenaceus indeed has a distinct dorsope, then P. pseudarenaceus and P. arenaceus are very similar and P. pseudarenaceus may be only a colour variety of the latter. The differences given by
Pseudosteres riphaeus , comb. nov.
Opius riphaeus
Tobias, 1986: 11, 12, 23 [holotype, ♀ (
Opius (Opiostomus) riphaeus; Fischer, 1991: 180–182;
Biology
Unknown.
Distribution
Iran, Russia (Asian part: Central Ural).
Cephaloplites
Cephaloplites
Szépligeti, 1897: 600;
Diagnosis
Antenna comparatively short, approximately as long as fore wing; scapus, fore coxa and trochanter distinctly compressed; face with pair of facial tubercles below antennal sockets more or less developed (Figs


Biology
Parasitoids of Agromyzidae (
Distribution
Palaearctic: three species.
Key to species of the genus Cephaloplites Szépligeti
| 1 | Head black; pair of tubercles of face indistinctly developed, only as a pair of small bumps, invisible in dorsal view of head and slightly visible in lateral view (Fig. |
C. tadzhicus Tobias & Saidov, 1995 |
| – | Head largely reddish or brownish yellow (Fig. |
2 |
| 2 | Setose tentorial depression smaller and closer to base of mandible (Fig. |
C. mocsaryi Szépligeti, 1897 |
| – | Setose tentorial depression larger and further removed from base of mandible (Fig. |
C. gijswijti sp. nov. |
Cephaloplites gijswijti sp. nov.
Type material
Holotype
, ♂ (
Diagnosis
Pair of tubercles of face distinctly developed, distinctly visible in dorsal view of head (Fig.
Description
Holotype, ♂, length of body 1.7 mm, of fore wing 2.0 mm.
Head. Antenna with 25 segments and as long as fore wing; third segment 1.3× longer than fourth segment, length of third, fourth and penultimate segments 3.0×, 2.1× and 1.7× their width, respectively, and apical segment with minute spine (Figs
Mesosoma. Length of mesosoma 1.3× its height (Fig.
Wings. Fore wing (Fig.
Legs. Hind femur, tibia, and basitarsus 3.3×, 7.7×, and 3.0× as long as wide, respectively (Fig.
Metasoma. Tergite I 1.3× longer than its apical width and slightly widened apically, its surface convex medially and largely smooth (only some rugulae posteriorly), dorsal carinae weakly developed and nearly up to apex of tergite (Fig.
Colour. Black; temple, frons largely and face laterally reddish yellow; remainder of face, clypeus, malar space, antenna and tergite II dark brown; palpi brown; coxae and trochanters black or dark brown, remainder of legs brownish yellow; pterostigma and veins brown; wing membrane subhyaline (Fig.
Biology
Unknown.
Distribution
Greece.
Etymology
Named after the aimable collector of the holotype, Martinus Johannes (Theo) Gijswijt (10.xi.1927–27.v.2015), who was one of the major specialists of European Chalcidoidea.
Cephaloplites mocsaryi
Cephaloplites mocsaryi
Szépligeti, 1897: 600–601;
Opius (Hypocynodus) kilisanus
Fischer & Beyarslan, 2005: 403–405;
Type material
Holotype
of C. mocsaryi (♀ from Hungary, Budapest, Zugliget) is lost. Holotype of O. kilisanus, ♂ (

Cephaloplites mocsaryi Szépligeti, ♀, Hungary (Budapest), but apex of antenna of Germany (Stuttgart) 56 wings 57 outer hind claw lateral 58 head anterior 59 face anterior 60 hind leg 61 mesosoma dorsal 62 head dorsal 63 face and mandible lateral 64 mandible lateral 65 metasomal tergite I dorsal 46 apex of antenna 67 habitus lateral. Scale bar: 1.0× (56, 58–62, 65, 67); 1.5× (63, 64); 2.5× (57, 66).
Biology
Parasitoids of Agromyzidae: Agromyza woerzi Groschke (
Distribution
Czech Republic, Germany, Hungary, and Turkey (Asian part).
Notes
The holotype of O. kilisanus has the mandible, malar space, temple ventrally, bases of hind and middle coxae and of trochantelli, mesosoma (except mesoscutum, scutellum, dorsal part of pronotum and mesopleuron) and metasoma (except second and most of tergite III) blackish, the temple somewhat less rounded and narrowed than figured for the female and the antenna with 26 segments. The differences are most likely clinal and considered to fall within the species limits of C. mocsaryi.
Cephaloplites tadzhicus
Cephaloplites tadzhicus Tobias & Saidov, 1995: 683–684.
Type material
Holotype
, ♂ (
Biology
Unknown.
Distribution
Central Asia: Tajikistan.
Notes
The venation of C. tadzhicus is similar to that of the type species (cf. Fig.

Acknowledgements
Many thanks to Sergey Belokobylskij and Konstantin Samartsev (
Additional information
Conflict of interest
The author has declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Funding
No funding was reported.
Author contributions
The author solely contributed to this work, except for the figures supplied by Dr Konstantin Samartsev as indicated in the paper.
Author ORCIDs
Cornelis van Achterberg https://orcid.org/0000-0002-6495-4853
Data availability
All of the data that support the findings of this study are available in the main text.
References
- Ameri A, Talebi AA, Rakhshani E, Beyarslan A, Kamali K (2014) Study of the genus Opius Wesmael (Hymenoptera: Braconidae: Opiinae) in Southern Iran, with eleven new records. Zootaxa 3884(1): 1–26. https://doi.org/10.11646/zootaxa.3884.1.1
- Beyarslan A, Fischer M (2013) Checklist of Turkish Opiinae (Hymenoptera, Braconidae). Zootaxa 3721(5): 401–454. https://doi.org/10.11646/zootaxa.3721.5.1
- Beyarslan A, Gözüaçik C, Güllü M, Konuksal A (2017) Taxonomical investigation on Braconidae (Hymenoptera: Ichneumonoidea) fauna in northern Cyprus, with twenty six new records for the country. Journal of Insect Biodiversity and Systematics 3(4): 319–334. https://doi.org/10.52547/jibs.3.4.319
- Fischer M (1961) Ergänzungen zur Kenntnis der paläarktischen Opius-Arten. Annali del Museo Civico di Storia Naturale di Genova 72: 1–16.
- Fischer M (1964) Die gezüchteten Opiinae der Sammlung Groschke (Hymenoptera, Braconidae). Stuttgarter Beiträge zur Naturkunde 136: 1–12.
- Fischer M (1971) Index to entomophagous insects. Hym. Braconidae. World Opiinae. Le Francois, Paris, 1–189.
- Fischer M (1972) Hymenoptera Braconidae (Opiinae I). (Paläarktische Region). Das Tierreich. 91(1973): 1–620.
- Fischer M (1977) Hymenoptera Braconidae (Opiinae II). (Amerika). Das Tierreich 96: 1–1001.
- Fischer M (1982) Die paläarktische Arten der Subgenera Misophthora Foerster und Agnopius n. des Opius Wesmael sowie über andere Opiinae (Hymenoptera, Braconidae, Opiinae). Folia Entomologica Hungarica 43: 21–37.
- Fischer M (1983) Illustrierte Redeskriptionen von Opiinen aus der UdSSR und neue Bestimmungsschlüssel zu den paläarktischen Arten des Subgenus Utetes Foerster (Hymenoptera, Braconidae). Annalen des Naturhistorischen Museums in Wien 85(B): 13–37.
- Fischer M (1986) Neue Bestimmungsschlüssel für paläarktische Opiinae, neue Subgenera, Redeskriptionen und eine neue Art (Hymenoptera, Braconidae). Annalen des Naturhistorischen Museums in Wien 88/89: 607–662.
- Fischer M (1987) Hymenoptera Braconidae (Opiinae III) - äthiopische, orientalische, australische und ozeanische Region. Das Tierreich 104: 1–734.
- Fischer M (1991) Wiederbeschreibungen und Neubeschreibungen von Opiinae aus der Alten Welt (Hymenoptera: Braconidae). Annalen des Naturhistorischen Museums in Wien 92(B): 139–203.
- Fischer M, Beyarslan A (2005) New maggot wasps from the Turkish fauna (Hymenoptera, Braconidae, Opiinae). Polskie Pismo Entomologiczne 74: 379–421.
- Gadallah NS, Ghahari H, Peris-Felipo FJ, Fischer F (2016) Updated checklist of Iranian Opiinae (Hymenoptera: Braconidae). Zootaxa 4066(1): 1–40. https://doi.org/10.11646/zootaxa.4066.1.1
- Jiménez-Peydró R, Peris-Felipo FJ (2011) Diversity and community structure of Opiinae (Hymenoptera: Braconidae) in the forest estate of Artikutza (Spain). The Florida Entomologist 94(3): 472–479. https://doi.org/10.1653/024.094.0311
- Li X-Y, van Achterberg C, Tan J-C (2013) Revision of the subfamily Opiinae (Hymenoptera, Braconidae) from Hunan (China), including thirty-six new species and two new genera. ZooKeys 268: 1–186. https://doi.org/10.3897/zookeys.326.5911
- Papp J (1981) Braconidae III/A. 13. Opiinae. Fauna Hungariae. Magyarorszag Allatvilaga. 144: 1–163.
- Papp J (1982) Braconidae (Hymenoptera) from Greece, I. Annales Musei Goulandris 5: 183–193.
- Papp J (2004) Type specimens of the braconid species by Gy. Szépligeti deposited in the Hungarian Natural History Museum (Hymenoptera: Braconidae). Annales Historico-Naturales Musei Nationalis Hungarici 96: 153–223.
- Szépligeti G (1897) Cephaloplites, novum genus Braconidarum. Természetrajzi Füzetek 20: 600–601.
- Telenga NA (1950) New species of hymenopterous parasitic wasps of the family Braconidae in USSR (Hymenoptera). Scientific Proceedings of the Institute of Entomology and Phytopatology 2: 293–308. [In Russian]
- Tobias VI (1998) Alysiinae (Dacnusini) and Opiinae. In: Ler PA (Ed.) Key to the insects of Russian Far East. Vol. 4. Neuropteroidea, Mecoptera, Hymenoptera. Pt 3: 1–706. Dal’nauka, Vladivostok, 299–411, 558–656.
- Tobias VI, Jakimavičius A (1986) Alysiinae & Opiinae, 7–231. In: Medvedev GS (Ed.) Opredelitel Nasekomych Evrospeiskoi Tsasti SSSR 3, Peredpontdatokrylye 4. Opr. Faune SSSR. Vol. 147, Section 3, Part 5, 1–308.
- Tobias VI, Saidov NS (1995) New species of braconid wasps (Hymenoptera, Braconidae) from Tajikistan. Entomologicheskoe Obozrenie 74(3): 681–684, 726. [In Russian] [Entomological Review 75(9): 33–37.]
- van Achterberg C (1988) Revision of the subfamily Blacinae Foerster (Hymenoptera, Braconidae). Zoologische Verhandelingen Leiden 249: 1–324.
- van Achterberg C (1990) Illustrated key to the subfamilies of the Holarctic Braconidae (Hymenoptera: Ichneumonoidea). Zoologische Mededelingen Leiden 64(1): 1–20.
- van Achterberg C (1993) Illustrated key to the subfamilies of the Braconidae (Hymenoptera: Ichneumonoidea). Zoologische Verhandelingen Leiden 283: 1–189.
- van Achterberg C (1997) Revision of the Haliday collection of Braconidae (Hymenoptera). Zoologische Verhandelingen Leiden 314: 1–115.
- van Achterberg C (2004) Bitomoides gen. nov. (Hymenoptera: Braconidae: Opiinae) from Europe. Zoologische Mededelingen Leiden 78(21): 331–335.
- van Achterberg C (2009) Can Townes type Malaise traps be improved? Some recent developments. Entomologische Berichten (Amsterdam) 69(4): 129–135.
- van Achterberg C, Salvo A (1997) Reared Opiinae (Hymenoptera: Braconidae) from Argentina. Zoologische Mededelingen Leiden 71(18): 189–214.
- Wharton RA (1987) Changes in nomenclature and classification of some opiine Braconidae (Hymenoptera). Proceedings of the Entomological Society of Washington 89(1): 61–73.
- Wharton RA (1988) Classification of the Braconid subfamily Opiinae (Hymenoptera). Canadian Entomologist 120(4): 333–360. https://doi.org/10.4039/Ent120333-4
- Wharton RA (1997) Generic relationships of opiine Braconidae (Hymenoptera) parasitic on fruit-infesting Tephritidae (Diptera). Contributions of the American Entomological Institute 30(3): 1–53.
- Yu DSK, van Achterberg C, Horstmann K (2016) Taxapad 2016, Ichneumonoidea 2015. Database on flash-drive. Nepean, Ontario, Canada.
- Zaykov AN, Fischer M (1983) Über Opiinae aus Bulgarien (Hymenoptera, Braconidae). Annalen des Naturhistorischen Museums in Wien 85: 39–51.