Research Article |
|
Corresponding author: Elisabeth Geiser ( elisabeth.geiser@gmx.at ) Academic editor: Caroline Chaboo
© 2023 Elisabeth Geiser.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Geiser E (2023) Revision of the Palaearctic species of the genus Plateumaris C. G. Thomson, 1859 (Coleoptera, Chrysomelidae, Donaciinae). In: Chaboo CS, Schmitt M (Eds) Research on Chrysomelidae 9. ZooKeys 1177: 167-233. https://doi.org/10.3897/zookeys.1177.103214
|
Abstract
Ten of the 27 species of Plateumaris Thomson (Chrysomelidae: Donaciinae) occur in the Palaearctic. Due to the intraspecific variation and the large distributions of some species, descriptions exist for at least 80 taxa plus five nomina nuda. The status of each valid species is clarified and the remaining 70 names are allocated as synonyms. New synonymies are P. tenuicornis Balthasar, considered a synonym of P. consimilis (Schrank), P. sulcifrons Weise as a synonym of P. rustica (Kunze), and P. caucasica Zaitzev as a synonym of P. sericea (Linnaeus). Two controversial synonyms are confirmed: P. discolor (Panzer) and P. sericea sibirica (Solsky) are both synonyms of P. sericea. Finally, P. obsoleta Jacobson is a synonym but at present it is not possible to decide whether it belongs to P. shirahatai Kimoto or to P. sericea. Forty-one new country records are added, compared with the Catalogue of Palaearctic Coleoptera published in 2010; 28 records are based on recently published records and 13 are first records for a specific country.
Key words
Identification key, leaf beetles, new country records, new synonymies, reed beetles, revision, Systematics, taxonomy
Introduction
The genus Plateumaris Thomson, 1859 belongs to the subfamily Donaciinae (reed beetles) which is part of the beetle family Chrysomelidae (leaf beetles). This family includes more than 37,000 (probably at least 50,000) species in more than 2,500 genera, making up one of the largest beetle families (
Most life stages in Chrysomelidae (larvae and imagines) are terrestrial; however, the larvae of the Donaciinae develop submerged on roots of aquatic plants. These larvae breathe by tapping the aerenchyma of the plant with two hollow abdominal hooks which are connected to their tracheal system. Therefore, they can stay permanently under water. Monophyly of Donaciinae is supported by these special morphological and physiological adaptions as well as by molecular data (
This number of ten species in the Palaearctic is not without controversy; in fact, different authors recognise nine to 19 species in the region (Table
A revision of the Palaearctic species of Plateumaris was needed, not only for the Palaearctic catalogue (
Knowledge of species distributions is necessary for systematic revisions. Therefore, I evaluated records from museum specimens and the literature. For the catalogue (
Materials and methods
For this revision, approximately 1500 adult specimens including six type specimens from 16 museum collections were examined. These museums are indicated below, together with other museums which are cited as depositories.
Museum acronyms
BMNH The Natural History Museum [formerly British Museum (Natural History)], London, UK (Michael Geiser, Maxwell V.L. Barclay, Keita Matsumoto, Dmitry Telnov)
NHRS Swedish Museum of Natural History, Stockholm (Johannes Bergsten)
SDEI Senckenberg German Entomological Institute, Müncheberg, Germany (Thomas Schmitt, Stephan Blank, Mandy Schröter)
ZMHB Natural History Museum, Berlin, Germany (Johannes Frisch, Bernd Jaeger)
Other abbreviations
A1, A2, … Number of an antennomere.
ab. aberratio.
coll. collection: the location where the specimen is stored.
det. determinavit: name of the person who identified this specimen.
ex. specimen(s).
ex coll. previous collection where the specimen was stored.
f. forma.
ICZN International Code of Zoological Nomenclature.
leg. legit: name of the person who collected this specimen.
T1, T2, … number of a tarsomere.
var. variatio.
vid. vidit: name of the person who confirmed the identification.
[] Text in square brackets [] are additions or comments by the author; in records data they do not form part of the label text tagged to the specimen.
[leg.] name of the person who probably collected this specimen.
[det.] name of the person who probably identified this specimen.
[new in PalCat] This indicates that the country mentioned was not listed in
[first record] If a country record was not published in detail until now, it is indicated as “first country record”. The same applies for a part of a country if it is treated as separate unit within a country, according to
// If original text is cited from different labels tagged to the pin of one specimen, the double slash indicates the text separation between these labels.
Data sources
Museum specimens
Except for Plateumaris sericea, it is very difficult to deliberately catch Plateumaris specimens in their habitat. Many voucher specimens were gained as by-catch in field studies motivated by other goals. The study of the variation of characters from many different sites of the whole distribution area is important for systematic revisions. Therefore, museum collections that have existed for a long time and where specimens from many different locations are stored are essential for such studies, but even smaller collections could provide a pleasant surprise to find some voucher specimens of a rare species from an interesting site.
The specimens in the museum collections were sorted in boxes with a species label. The compilation was usually done by non-specialists, who sorted the various collections donated to the museum into the overall collection, regardless of whether they were correctly identified or not. The vouchers of P. bracata, P. consimilis, P. rustica, and P. sericea were usually correctly identified. Specimens of other Plateumaris species are rare, and these were all examined in detail. Some of them already had identifier labels. If not, I tagged my label on the needle with the information “genus species Author det. E. Geiser Year”. I also did that when a specimen was misidentified, leaving the original identifier label on the needle. When a species was difficult to identify I used a similar label, with “vid. E. Geiser Year” to state that I have confirmed the identification. Specimens of the four species mentioned above were also studied carefully for the purpose of this revision. Here I did not tag my label to all of them, but mainly on vouchers that showed some variation of the typical characters or were collected in an interesting location.
The importance of museum collections has increased in recent years. Some areas of the Palaearctic region are inaccessible today for political or security reasons like Xinjiang in western China, or Syria, or Afghanistan. Fortunately, some natural history field studies dating more than 100 years ago enabled vouchers to be deposited in European museums. These historic specimens are an invaluable source for further studies. Additionally, it is more difficult today to collect Donaciinae and therefore Plateumaris species because man-made changes of limnic and wetland environments are contributing to the decline of habitats and, therefore, of species. Many previous habitats no longer exist and the only chance to study specimens from these districts is to examine the museum vouchers.
Main literature sources
In addition to the museum specimens studied, the author relied on certain important publications for additional data, mainly
Original descriptions and type specimens
The study of original taxonomic descriptions is essential for any systematic revision. Many of the species or subspecies were first described in Latin with very specific phrases used in scientific entomological scripts in the 18th and 19th centuries. Some original descriptions are in German, French, or Russian; some are multilingual, starting with Latin for the main characters of a presumed new species, and then more details were added in the native language of the author. Therefore, to overcome these challenges,
The study of type specimens is also essential in revisions, and I was able to examine six type specimens. Many types from the 18th and 19th centuries could not be located or no longer exist. Another problem is that specimens in collections may be labelled as “type” or indicated as types by red or red-bordered labels but are not actually type specimens.
Synonyms
For each species, in “Taxonomic history and synonymies” most synonymies are explained, where changes were made (
Distribution data
For each species, the countries, or the part of a country, with reliable records are listed. Detailed information on the data source is given for the countries or part of a country which are new in
Results
Identification keys
Many previous researchers have published keys to the Palaearctic species of Plateumaris, but these keys often use characters that are difficult to see from the beginning. Although only ten species occur in the Palaearctic region, some specimens are not easy to identify even for experienced coleopterologists. The new key reduces the species designation available for a particular specimen. The detailed characters and the pictures provided in the species sections should allow definitive identification. Because only two species occur in the whole Palaearctic region, the separation of the West and East Palaearctic in different keys leads to more reliable identifications. If the information on the specimen location label is very imprecise (e.g., “Siberia” or “Russia”) then both keys need to be applied.
Plateumaris species which occur in the West Palaearctic
| 1 | Antennae and legs entirely metallic, same colour as pronotum and elytra. Sometimes the antennomeres can be reddish near the joints. At the legs small reddish parts may be occur near the joints or, exceptionally, on the tibiae or tarsomeres. Pronotum and elytra in various metallic colours (Fig. |
P. sericea |
| – | Antennae and legs yellow reddish brown, some parts more or less darkened | 2 |
| 2 | Pronotum and elytra black or with dark metallic lustre, elytra elongate, their length twice as long as wide, side contour of elytra parallel, not convex, the largest of all species (Fig. |
P. bracata |
| – | Elytra not twice as long as wide | 3 |
| 3 | Pronotum cordate, upper side with metallic lustre in different colours or entirely black, elytra 1.5–1.8× longer than wide, never 2× longer than wide, side contour of elytra not parallel but slightly convex (Figs |
P. consimilis |
| – | Pronotum not cordate | 4 |
| 4 | Pronotum distinctly flattened, almost quadrate, only slightly constricted at the basis, with flat disc and indistinct anterior tubercles, upper side bronze or black with greenish, bluish, or purplish metallic lustre, colour of pronotum and elytra mostly the same but can also differ significantly (Fig. |
P. rustica |
| – | Pronotum neither distinctly cordate nor flattened, outline subquadrate, slightly longer than wide, basal part narrowed, slightly cone-shaped, anterior tubercles distinctly visible or almost entirely smooth, upper side cupreous or bronze, sometimes metallic green, blue, purple, or non-metallic brown (Fig. |
P. weisei |
Remarks: Some specimens of P. weisei are difficult to distinguish from P. consimilis and P. rustica. The variation of several characters is sometimes within the same range. It occurs also within the distribution area of P. consimilis and P. rustica in southeast Finland and some parts of Russia, though these three species are very rare here. To avoid misidentification, use all the details and figures provided in the species sections.
Plateumaris species which occur in the East Palaearctic
| 1 | Femora and tibia entirely metallic in colour | 2 |
| – | Femora and tibia partly metallic or dark, femoral base usually rufous | 3 |
| 2 | Median line of pronotum deep, male pygidial apex usually emarginate; apex of median lobe of male genitalia without subapical corner; apex of median ejaculatory guide of endophallus rounded (Figs |
P. sericea |
| – | Median line of pronotum indistinct, male pygidial apex usually truncate; apex of median lobe of male genitalia with subapical corner; apex of median ejaculatory guide of endophallus notched (Figs |
P. shirahatai |
| 3 | Metafemoral tooth present but blunt, pronotum neither distinctly cordate nor flattened, outline subquadrate, slightly longer than wide, basal part narrowed, slightly cone-shaped, anterior tubercles distinctly visible or almost entirely smooth, upper side cupreous or bronze, sometimes metallic green, blue, purple, or non-metallic brown (Fig. |
P. weisei |
| – | Metafemoral tooth well developed | 4 |
| 4 | Body robust in shape, with conspicuously short elytra and legs, elytra length is only 1.6× longer than width, at least always less than 1.7, pronotum finely and densely punctate; colour black or bronze-metallic, also with a bluish or greenish lustre, elytra shiny (Fig. |
P. akiensis |
| – | Elytra and legs not conspicuously short, normal, ratio of elytral length to width> 1.7 | 5 |
| 5 | Pronotum cordate, surface shiny as if varnished, most parts without wrinkles or punctures, anterior tubercles protruding but smooth, gently narrowed with a shallow, transverse groove behind them, dorsal colouration variable (Fig. |
P. constricticollis |
| – | Pronotum not cordate, covered with rugae and punctures | 6 |
| 6 | Antennae and legs mostly rufous, sometimes apically darkened, femora reddish on the basal half and metallic-dark on the apical half (Fig. |
P. roscida |
| – | Legs usually rufous, sometimes apical area of femora dark (Fig. |
P. amurensis |
Remarks: The three species found in the east of Russia and northeast China, P. amurensis, P. roscida, and P. weisei, are not easy to distinguish because their external characters sometimes overlap. To distinguish P. amurensis from P. weisei see also Table
Opinions on species delimitation in the literature
Although only a few Palaearctic Plateumaris species exist (compared with other beetle genera), there are many differing opinions between authors. Almost every comprehensive publication about Palaearctic Plateumaris shows a different number of species (Table
The highest number of species is recorded by
Two major books were published in 2010: Warchalowski treated 12 species in his identification key and Silfverberg listed 16 species (and some more subspecies) in
Comparison of major publications: names of Palaearctic species of the genus Plateumaris Thomson including the different opinions about valid species and synonyms.
| Plateumaris species names | Palaearctic Region | Parts of Palaearctic Region | ||||||
|---|---|---|---|---|---|---|---|---|
|
|
Warchalowski 2010 |
|
|
|
Japan: |
Russia: Bienkowski 2014 | China & Korea: |
|
| P. affinis | * | syn of P. rustica | syn of P. rustica | * | * | — | syn: P. rustica | — |
| P. akiensis | * | * | * | o: described in 1984 | o: described in 1984 | * | — | — |
| P. amurensis | * | * | “syn of weisei by Goecke which is probably correct” | o: clarified 2001 to be different from P. weisei | syn of P. weisei | — | * | * |
| P. bracata | * | * | * | * | * | — | * | o: first record 2023 |
| P. caucasica | * | o | possible syn of P. roscida | * | o | — | ssp of P. sericea | — |
| P. consimilis | * | * | * | * | * | — | * | — |
| P. constricticollis | * including 3 ssp | * | * | * | * | * including 3 ssp. | — | — |
| P. discolor | * | syn of P. sericea | syn of P. sericea | * | * | syn of P. sericea | * | — |
| P. mongolica | syn of P. weisei | syn of P. weisei | probable syn of P. weisei | * | * | syn of P. weisei | — | * |
| P. obsoleta | * | * | probable syn of P. sericea | * | * | — | * | — |
| P. roscida | * | * | * | * | * | — | * | as P. annularis |
| P. rustica | * | * | * | * | * | — | * | — |
| P. sachalinensis | syn of P. weisei | * | probable syn of P. weisei | * | o: described in 1973 | syn of P. weisei | * | — |
| P. sericea | * | * | * | * | * | * | * | as P. socia |
| P. shirahatai | * | as syn of P. obsoleta | * | * | o: described in 1971 | * | * | o: described in 1971 |
| P. sibirica | ssp of P. sericea | ssp of P. sericea | o | o | syn of P. sericea | ssp of P. sericea | ssp of P. sericea | — |
| P. socia | syn of P. sericea sibirica | syn of P. sericea sibirica | probable syn of P. sericea | * | o | syn of P. sericea | — | * |
| P. sulcifrons | * | * | probable syn of P. rustica | * | * | — | — | — |
| P. tenuicornis | * | o | probable syn of P. consimilis | * | * | — | — | — |
| P. weisei | * | * | * | * | * | * | * | o |
| Number of valid species | 16 | 12 | 9 | 17 (+2) | 13 (+4) | |||
Taxonomic accounts
Plateumaris
Plateumaris C. G. Thomson, 1859: 154.
Euplateumaris Iablokoff-Khnzorian, 1966: 121.
Juliusina Reitter, 1920: 41.
Type species and localities
Plateumaris C. G. Thomson, 1859: Donacia nigra Fabricius, 1792: Germania.
= Prionus bracatus Scopoli, 1772: Carniola [now Slovenia].
Euplateumaris Iablokoff-Khnzorian, 1966: Leptura sericea Linnaeus, 1758: Europe.
Juliusina Reitter, 1920: Prionus bracatus Scopoli, 1772: Carniola [now Slovenia].
Timeline of taxonomic history and synonymies
1758: Linnaeus described the genus Leptura with 22 species. Two of these species belong to the (later established) Donaciinae: Donacia aquatica and Plateumaris sericea (
1760: Linnaeus mentioned Leptura sericea in his ‘Fauna Suecica’ with the same diagnosis as in
1762: Geoffroy erected the genus Prionus for the species Cerambyx coriarius Linnaeus, 1758. It now belongs to the Cerambycidae as does the genus Leptura.
1772: Scopoli described Prionus bracata.
1775: The genus name Donacia was erected by Fabricius. He described Donacia crassipes and Donacia simplex and assigned Leptura aquatica L., 1758 to the genus Donacia, but he did not change the genus name of Leptura sericea L., 1758. Other Plateumaris species were described as Leptura, e.g., Leptura consimilis Schrank, 1781. It is remarkable that these early entomologists had already assigned them to a genus other than Donacia.
1796: The category “Chrysomelidae” was established between order and genus for insects by Latreille.
1802: Latreille established the coleopteran family Chrysomelidae.
1837: Kirby established the subfamily Donaciinae.
1859: The genus name Plateumaris was erected by C. G. Thomson (translation in
1920: Reitter split the Palaearctic Plateumaris into two subgenera: Plateumaris sensu stricto and Juliusina. He assigned P. sericea and P. discolor to Plateumaris s. str. and he described P. annularis in a footnote to P. sericea. In the same footnote he assigned P. obsoleta to P. annularis, but not as a synonym, and he placed P. amurensis near to P. discolor. The subgenus Juliusina contained P. bracata, P. consimilis, P. rustica, and P. affinis. To the latter species he assigned P. sulcifrons and P. mongolica. Moreover, Reitter made no designation of type specimens to either of the new subgenera.
Type species designations
The names Plateumaris Thomson, 1859 and Juliusina Reitter, 1920 are based on the same type species (Prionus bracatus) which makes the subgenus Juliusina a synonym of Plateumaris in the sense of the whole genus, which was endorsed by
Taxonomic status of Plateumaris
Most European authors accepted the genus Plateumaris, but North American authors were reluctant and oscillated between the use of this name at generic and subgeneric ranks within the genus Donacia. This ambiguity led to confusion about the genus name Plateumaris, worsening the existing confusion over the name Plateumaris of the Palaearctic species (see above).
Previous subgeneric classifications
In central Europe, the widely used key of
Diagnosis of the genus Plateumaris Thomson, 1959
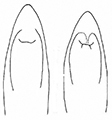
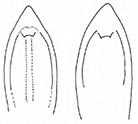
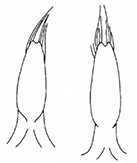
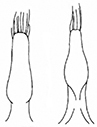
- Sutural margin of elytron explanate apically, sutural interval sinuates distinctly before apex, lower sutural margin broadly exposed (Fig.
1A ); - Elytral apex rounded, inner angle sharp, no outer angle protruding;
- First abdominal segment as long as the others combined;
- Host plants are typically Cyperaceae, but also a few other wetland plants.
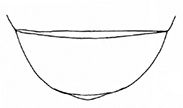
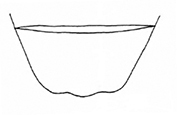
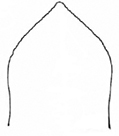
The aedeagus of Plateumaris species and some other representatives of the Donaciinae consists of a median lobe which contains the endophallus and the lateral parameres (Fig.

Biology
Reed beetles live on plants in wetlands. The larvae develop attached to the roots in the sediment and live as sap suckers gnawing a hole into the root. They breathe by tapping the aerenchyma of the plant using two hollow abdominal hooks, which are connected to their tracheal system; therefore, they can live even in anoxic mud (
The larval host plants are mostly members of Cyperaceae, but include some Juncaceae and Poaceae (e.g., Phragmites australis (Cav.) Trin. ex Steud.). Acorus calamus L. (Araceae), Caltha palustris L. (Ranunculaceae), and lris versicolor L. (Iridaceae) are also mentioned as food plants (
Plateumaris species prefer habitats of wet sedge meadows, peat bogs, and fens, in contrast to Donacia species which live on aquatic plants with emerging parts and in contrast to Macroplea species which live on totally submerged aquatic plants.
Historical biogeography
Plateumaris species are found only in the Holarctic region with ten species in the Palaearctic and 17 species in the Nearctic (
Checklist and distribution
A summary of the distribution of the Plateumaris species in the Palaearctic region is shown in Table
Checklist and distribution of the Plateumaris species in the Palaearctic region.
| Plateumaris species | Distribution | |
|---|---|---|
| 1 | P. akiensis Tominaga & Katsura, 1984 | Japanese endemic: records only from Hiroshima prefecture so far |
| 2 | P. amurensis Weise, 1898 | Russia: East Siberia and Far East |
| 3 | P. bracata (Scopoli, 1772) | West Palaearctic except of south Mediterranean including Kazakhstan and west Siberia |
| 4 | P. consimilis (Schrank, 1781) | Europa and west Siberia |
| 5 | P. constricticollis (Jacoby, 1885) | Endemic of the Japanese archipelago |
| 6 | P. roscida Weise, 1912 | East Siberia and Far East, from northern China to Amur River region, Lake Baikal, and the Sakha (Yakutia) Republic |
| 7 | P. rustica (Kunze, 1818) | West Palaearctic |
| 8 | P. sericea (Linnaeus, 1758) | Palaearctic |
| 9 | P. shirahatai Kimoto, 1971 | East Palaearctic: Russian Far East, Japan, and South Korea |
| 10 | P. weisei (Duvivier, 1885) | Northern Europe and northern Asia, Japan, and South Korea |
Palaearctic region: Plateumaris sericea has the largest distribution area of any Donaciinae species: it is recorded from Ireland and Great Britain to the whole of continental Europe, North Africa, and almost all Asian countries which belong to the Palaearctic region. Plateumaris weisei occurs from northern Europe to east Asia, from Siberia to northern China, Japan (Hokkaido), and South Korea.
Western Palaearctic region: Three species occur only in Europe and in Asia west of Lake Baikal: P. bracata, P. consimilis, and P. rustica.
Eastern Palaearctic region: Five species occur only in Asia east of Lake Baikal: P. amurensis, P. roscida, and P. shirahatai, and two species are endemic to the Japanese archipelago, P. akiensis and P. constricticollis.
Palaearctic species of the genus Plateumaris
Plateumaris akiensis
Plateumaris akiensis Tominaga & Katsura, 1984: 25.
Type locality
Japan, Honshu, Shinkawa-tameike, altitude 770 m, Nishi-yawata-hara, Geihoku-cho, Yamagata-gun, Hiroshima Prefecture.
Type material
Holotype
: Japan • ♂; Honshu, Hiroshima Prefecture, Yamagata-gun, Geihoku-cho, Nishi-yawata-hara, Shinkawa-tameike; 770 m a.s.l.; 13 Jun 1982; I. Hiura leg.;
Taxonomic history
Because this species was not described until 1984 it is possible that some small Plateumaris specimens from Japan are misidentified as another species in old collections.
Diagnosis
Habitus (Fig.

Description
There are comprehensive descriptions in
Size : 6.4–7.6 mm.
Colour : Colour of pronotum and elytra entirely metallic black or reddish coppery, some specimens with a bluish or greenish lustre.
Head : Supraocular furrow indistinct, frontal tubercle weakly convex.
Antennae : Antennae short and robust, slightly shorter than half as long as the body, A1 dark, A2–A11 entirely reddish brown or apical antennomeres with darker parts. A1 is longest, A2 is shortest, A3–A11 are only slightly longer than A2, A3 ≥ A4 ≤ A5.
Pronotum : Slightly longer than wide, outline more or less cordate, gradually narrowed backwards with anterior margin strongly produced forward; dorsal surface with feebly raised anterior and posterior tubercles, disc densely but finely punctate, median line shallow, with a conspicuous broad collar along the posterior margin.
Elytra : Robust and short, 1.6× as long as wide, with arched outer margin, rows of punctures regular, their interstices smooth, usually sparsely but sometimes densely punctate.
Legs : Legs short, reddish brown, partly dark rufous or black, especially the apical half of the femora, metafemur robust in shape, with a tooth; T2 shorter than its width, T1 > T2 < T3.
Pygidium : Entirely rufous, but apical part black, apex pubescent, shallowly emarginate in both sexes.
Aedeagus
: See Fig.
Similar species
Plateumaris akiensis looks similar to P. constricticollis but can be identified by its shorter elytra. In P. constricticollis the elytral length is distinctly > 1.6× longer than wide.
Biology
The adults feed on pollen of Carex sp. and Scirpus juncoides (
Distribution
Endemic species found in Honshu, Japan. The only records so far are from Hiroshima and Shimane prefectures in south-west Honshu.
Material examined
Five specimens from Hiroshima prefecture, stored in BMNH,
Plateumaris amurensis
Plateumaris amurensis Weise, 1898: 179.
Type locality
Amur [assumed Russia, further details unknown].
Type material
Types could not be located so far. In his original description (translation in

Taxonomic history and synonymies
Diagnosis
Metafemur with a prominent, blade-like tooth, apical part of median lobe of aedeagus gradually narrowed towards the apex.
Description
Size : 7.1–7.7 mm.
Colour : Pronotum and elytra bronze, cupreous, also which greenish reflex.
Head : Supraocular furrow indistinct, vertex pubescent with deep median line. Antennae entirely rufous, sometimes apex darkly rufous; A4 = 1.6× A2, A5 longest and ~ 2.5× as long as wide.
Pronotum : More or less quadrate, anterior part slightly widened by shallow anterior tubercles, disc shiny, coarsely punctate, rugose, sometimes with microsculpture in major part of the disc, basal sulcus prominent with rugae and dense punctures, median groove indistinct.
Elytra : Sparsely rugose, shiny, densely punctate on disc.
Legs
: In most specimens rufous, sometimes apical area of femora dark, metafemur with a prominent, blade-like tooth (Fig.
Pygidium
: Apex pubescent, shallowly emarginate or sometimes truncate in male and rounded in female. Last sternite entirely coppery but apex at middle part rufous, apical shape variable in male, acute in female (Table
Ovipositor
: Elongate, both sides paralleled, apical angle acute, subapical corner with teeth, apex remarkably prominent (Table
Aedeagus
: With median lobe, acute but slightly rounded at the apex; cap of tegmen gradually narrowed distally, notched, or sometimes rounded at apex (Fig.
Similar species
The most similar species is Plateumaris weisei and the main distinguishing characters are shown in Table
Biology
Host plant and larvae are unknown.
Distribution
East Palaearctic only. Records exist for Asia: Russia: Transbaikalia, Republic of Sakha (Yakutia): southern part of river Lena; Amur Oblast, Khabarovsk Krai, Primorsky Krai, Sakhalin, Kurile Islands. Sometimes P. amurensis is mentioned from Japan (e.g.,
Material examined
15 specimens from different localities in East Siberia and Far East.
Plateumaris bracata
Prionus bracatus Scopoli, 1772: 100.
Donacia abdominalis Olivier, 1800: 9.
Plateumaris bracatus var. fairmairi LeGrand, 1861a: 265.
Donacia nigra Fabricius, 1792: 117.
Donacia palustris Herbst, 1784: 100.
Leptura violacea Pallas, 1773: 724.
Type locality
Plateumaris bracatus: Carniola, a historical region which comprised parts of present-day Slovenia.
Type material
Type specimens of P. bracatus do not exist anymore. Sadly, Scopoli’s collection of insects from Carniola decayed during his life time. He committed this collection to a printer in Vienna who apparently did not store it adequately. Before all the pictures were printed, Scopoli complained in his 1773 letter to Linnaeus (

Taxonomic history and synonymies
In nearly all publications the species name has been misspelled “braccata”. The spelling in the original description by
Donacia abdominalis (Olivier, 1800):
Plateumaris fairmairi was first described by
Fabricius described Donacia nigra by 1792 (see
Diagnosis
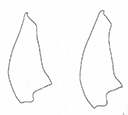
This is the largest Plateumaris species, body length: 8.0–12.0 mm (Fig.
Head, pronotum and elytra entirely black or with a weak blue, purple or green metallic lustre, antennae and legs reddish brown in most specimens, elytra elongate, ~ 2× as long as wide, side contour of the elytra very straight, almost parallel.
Description
Size : 8.0–12.0 mm.
Colour : Dark, entirely black or at the most with weak blue, purple or green metallic lustre.
Head : Entirely black or with weak metallic lustre. Fine punctures and very fine wrinkles. Antennae minimum as long as the half length of the beetle, basal part of the antennomere always reddish (with rare exceptions), apical part dark; ½ A1 ≥ A2, ½ A3 ≥ A2, A4 ≥ A3, A1 ≈ A4, A5 … A11 ≈ A4.
Pronotum : Cordate, ahead distinctly wider than behind, anterior tubercles only slightly convex, slightly pubescent. Disc with fine punctures, median line well developed, sometimes shortened, but also very shallow and almost invisible in some specimens.
Elytra : Base of elytra with short, distinct setae in most specimens, elytral disc rugose, punctures delicate, not deep, interstices strongly transversely wrinkled; contour of the margin very straight, almost parallel; elytra elongate, ~ 2× as long as wide, ratio of length to width = (1.8–2.1): 1.
Legs : Colour variation from complete reddish brown to dark apical parts and completely dark legs, femora basally very broad, metafemur with broad tooth, robust in male, in female feeble or indistinct.
Aedeagus
: see Fig.
Similar species
The most similar species is Plateumaris consimilis, which is smaller (6.0–9.2 mm) on average, its elytra are shorter with a ratio of length to width ≤ 1.8, and the outer contour of the elytra are slightly rounded, not parallel.
Biology
Plateumaris bracata is monophagous on Phragmites australis (Cav.) Trin. ex Steud., the common reed, Poaceae (
Distribution
All parts of Europe except southern Europe and north Scandinavia, continuing to central Asia, including southern parts of Russia and western Siberia. Records exist for Europe: Austria, Belgium, Bosnia-Herzegovina [new in PalCat], Belarus, Bulgaria, Croatia [first record], Czech Republic, Denmark, Estonia, Finland, France, Germany, Great Britain, Hungary, Ireland, Italy, Latvia, Lithuania, Luxembourg, Moldavia, Montenegro [first record], The Netherlands, Norway, Poland, Romania, Russia (central and south parts of European Russia), Serbia [new in PalCat], Slovakia, Slovenia, Sweden, Switzerland, Ukraine.
Asia: Azerbaijan, China (Xinjiang [first record]), Georgia, Iran [new in PalCat], Kazakhstan, Kyrgyzstan [new in PalCat], Russia (south Siberia [new in PalCat], west Siberia).
New country records additional to Silfverberg (2010)
Bosnia-Herzegovina:
Croatia • 3 ex.; Dalmatia; E. Geiser 2019 det.; coll. Frey in
Montenegro • 2 ex.; Montenegro; E. Geiser 2019 det.; coll. Frey in
Serbia:
China • 2 ex.; Xinjiang, “Ost-Turkestan, Bagratsch-Kul” [Bosten-Lake], Kurla; May 1902; Hauser leg.; E. Geiser 2019 det.;
Iran • 2 ex.; Khorasan-e Razavi province, Sabzevar; 36°12′N, 57°35′E; 1024 m a.s.l.; 23. Aug 2012 (
Kyrgyzstan:
Russia: South Siberia (
Material examined
More than 100 specimens from different localities throughout the distribution area.
Plateumaris consimilis
Leptura consimilis Schrank, 1781: 155.
Plateumaris consimilis f. aerea Bechyné, 1942: 234, 236 [infrasubspecific name].
Leptura assimilis Schrank, 1781: 156.
Plateumaris consimilis f. coerulea Bechyné, 1942: 234, 236 [infrasubspecific name].
Donacia discolor Hoppe, 1795: 45 [homonym].
Plateumaris consimilis f. flavipes Bechyné, 1942: 234, 236 [infrasubspecific name].
Plateumaris consimilis f. nigripes Bechyné, 1942: 234, 236 [infrasubspecific name].
Donacia rufipes Olivier, 1791: 292.
Plateumaris tenuicornis Balthasar, 1934: 128 [syn. nov.].
Donacia variabilis Kunze, 1818: 39.
Plateumaris consimilis f. violacea Bechyné, 1942: 234, 236 [infrasubspecific name].
Plateumaris consimilis f. viridis Bechyné, 1942: 234, 236 [infrasubspecific name].
Type locality
Plateumaris consimilis: unknown, but possibly in Austria (the country in 1781 was much larger than today) because the original description is in a book titled ‘Enumeratio insectorum Austriae indigenorum’.
Type material

Taxonomic history and synonymies
Bechyne (1942) detailed statistics about the different colours and subtle structures on the pronotum of P. consimilis, but without convincing results. He named colour variations as “forma” but conceded that there also existed combinations of colours in between. These names are indicated above for the sake of completeness, but they are irrelevant to systematics.
Donacia discolor was described by
Plateumaris consimilis orientalis was described by
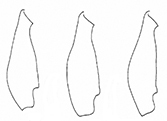
Diagnosis
Pronotum cordate, anterior tubercles slightly convex. Upper side with metallic lustre in varying colours, antennae, and legs at least partly reddish brown. Elytra 1.5–1.8× longer than wide.
Description
Size : 6.0–9.2 mm.
Colour
: Very variable, upper side greenish, bluish, cupreous, bronze, or black with metallic lustre, some black specimens almost without metallic lustre (Fig.
Head : Same colour as pronotum. Antennae approximately half the body length or slightly longer, Antennomeres reddish brown at least at the basal part, the apical part can be darkened. ½ A1 ≥ A2, ½ A3 ≥ A2, A4 ≥ A3, A1 ≈ A4, A5 … A11 ≈ A4.
Pronotum : cordate, ahead wider than behind, anterior tubercles only slightly convex, disc uniformly punctate, median line obsolete to fine but distinct.
Elytra : 1.5–1.8× longer than wide, never twice as long as wide, side contours slightly convex, not parallel, elytra rugose and punctulate.
Legs : Colour variation from completely reddish brown to only reddish at the joints, femora basally very broad, metafemora with sharp or broad tooth.
Aedeagus
: The shape varies between the short, more rounded form in P. bracata and the elongated acute form of P. rustica (Fig.
The most similar species are Plateumaris bracata and P. rustica: P. bracata has longer elytra, ~ 2× as long as wide and the side contour of the elytra is parallel, not convex (Fig.
Biology
Although P. consimilis is one of the common Plateumaris species, its larva was not described until 2014 by Medvedev and Muravitsky. The larvae and adults feed on Carex sp. (Cyperaceae). Also, Juncus articulatus (Juncaceae) and Caltha sp. (Ranunculaceae) are mentioned as food plants of adults, on which they feed on the pollen (
Distribution
Western Palaearctic: mainly continental Europe up to southern Sweden, very rare in south and east Europe and west Siberia. Records exist for: Europe: Albania [first record], Austria, Belarus, Belgium, Bosnia-Herzegovina, Bulgaria, Croatia, Czech Republic, Denmark, France, Germany, Hungary, Italy, Latvia, Liechtenstein [new in PalCat], Lithuania, Luxembourg, The Netherlands, North Macedonia [new in PalCat], Poland, Romania, Russia (central part of European Russia), Serbia [new in PalCat], Slovakia, Slovenia, Spain, Sweden, Switzerland, Ukraine.
Asia: Georgia [first record], Russia (west Siberia), Turkey [new in PalCat].
New country records additional to Silfverberg (2010)
Albania [first records] • 3 ex.; Qarku i Kukësit, Kula e Lumës, “Albanien Expedition, Kula Ljums”; 18–28 May 1918; H. Goecke 1956 det., E. Geiser 2019 vid.;
Liechtenstein:
North Macedonia:
Serbia: Gavrilovic and Curcic (2011).
Georgia [first record] • 2 ex.; Mtskheta, “Transcaucasia, Mazcheta, pr. Tiblisi”; 4–23 Jun 1987; Wrase and Schülke leg.; E. Geiser 2019 det.;
Turkey:
Remarks
Plateumaris consimilis does not occur in the East Palaearctic which has also been confirmed recently by
Material examined
More than 200 specimens from different localities throughout the distribution area.
Plateumaris tenuicornis , syn. nov.
Type locality
Bosnia-Hercegovina: Dol. Tuzla, Bosnia.
Type material
Holotype
of Plateumaris tenuicornis. Bosnia-Herzegovina • 1 ex.; Bosnia, Dol. [Dolina?] Tuzla; Em. Fritsch leg.;
Remarks
Because Bechyné only indirectly treated P. tenuicornis as a synonym, and because Askevold only suggested that P. tenuicornis should be considered as a probable new synonym, I determined that P. tenuicornis Balthasar, 1934 is a new synonym of P. consimilis (Schrank, 1781) according to

Plateumaris constricticollis
Donacia constricticollis Jacoby, 1885: 192.
Plateumaris constricticollis babai Chûjô, 1959: 2.
Donacia constricticollis constricticollis Jacoby, 1885: 192 (= Plateumaris constricticollis kurilensis L. N. Medvedev, 1978b: 83).
Plateumaris constricticollis toyamensis Tominaga & Katsura, 1984: 27 (= Plateumaris constricticollis chugokuensis Tominaga & Katsura, 1984: 28).
Type localities
Plateumaris constricticollis: Japan; P. constricticollis babai: Honshu: Niigata Prefecture, Yoshigahira, Shitada-mura; P. constricticollis toyamensis: Tsubura-ike, alt. 690 m, Kamiichi-machi, Naka-niikawa-gun, Toyama prefecture; P. constricticollis chugokuensis: Koiga-kubo, alt. 570 m, Tessei-cho, Atetsu-gun, Okayama prefecture.
Type material
Holotype of P. constricticollis: Japan • 1 ♂; Hokkaido, Lake Junsai, N of Hakodate; 43°7'N, 145°6'E; 28–30 Jul 1880; G. Lewis leg.; BMNH 1910-320. The holotype was examined in 2019.
Holotype
of P. constricticollis babai: Japan • 1 ♀; Honshu, Niigata Prefecture, Yoshigahira in Mt. Sumon; 25 Jun 1955; K. Baba [leg.]; “P. constricticollis babai Chȗjȏ” M.
Holotype
of P. constricticollis toyamensis: Japan • 1 ♂; Honshu, Toyama-Prefecture, Kamiichi-machi, Naka-niikawa-gun, Nakanomata, Tsubura-ike; 690 m a.s.l.; 20 Jun 1983; K. Katsura leg;
Holotype
of P. constricticollis chugokuensis: Japan • 1 ♂; Honshu, Okayama prefecture, Tessei-cho, Atetsu-gun, Koiga-kubo; 570 m a.s.l.; 13 Jun 1982; O. Tominaga leg.;
Taxonomic history and synonymies
Specimens of P. constricticollis reveal a remarkable variation in body size and colouration, pronotal disc, and even genital structures. This resulted in the description of four subspecies. Further studies concluded that there were two subspecies in addition to the nominate species, and therefore the other two subspecies names are synonyms (
Chûjô (1959) was the first to describe a subspecies, P. c. babai from Niigata Prefecture. In 1978, Medvedev described P. c. kurilensis from Kunashiri, the southernmost Kurile Island, near Hokkaido. This subspecies was synonymised with P. c. constricticollis by
Diagnosis
The characters common to all subspecies are the following: surface very shiny (Fig.
Description
Size : 6.6–11.9 mm.
Plateumaris constricticollis and its subspecies have been thoroughly studied by Japanese colleagues, with detailed descriptions of their morphological characters and their variations, as well as phylogeny, biogeographical history, biology, and molecular analyses (
The similar species Plateumaris weisei is not shiny or glossy, its pronotum not cordate, and the legs are longer and slenderer.
Biology
Distribution
Plateumaris constricticollis is endemic to the Japanese Archipelago (Hokkaido, Honshu, and Kunashiri).
Plateumaris constricticollis constricticollis: southernmost Kurile Island Kunashiri, Hokkaido and northern part of Honshu until prefecture of Yamagata (
Plateumaris constricticollis babai: Honshu: prefectures Fukushima, Niigata, Nagano, Gunma, Tochigi, Ibaraki, and Chiba.
Plateumaris constricticollis toyamensis: Honshu: prefectures Toyama, Gifu, Ishikawa, Aichi, Hyogo, and Okayama.
Material examined
Approximately 60 specimens from Japan.
Plateumaris roscida
Plateumaris roscida Weise, 1912: 77.
Plateumaris annularis Reitter, 1920: 41.
Type localities
Plateumaris roscida: Russia, Transbaikalia (Zabaykalsky Krai): Tschita; Plateumaris annularis: Russia, Far East, Khabarovsk Krai, Nikolayevsk-on-Amur.
Type material
Holotype of P. roscida: Russia • 1 ♂; East Siberia, Transbaikalia, Tschita; ZMHB. Label text: “Typus [red]// roscida m. //coll. J. Weise //Zool. Mus. Berlin //Holotype ♂ Plateumaris roscida Weise [red]//Plateumaris roscida Weise det. I.S. Askevold 1989”.
No location label is tagged to the holotype, but the type locality “Tschita” is indicated in the original description by
Holotype of Plateumaris annularis: Russia • 1 ex.; Far East, Khabarovsk Krai, Nikolayevsk-on-Amur, “Amur region, Chabarowsk, Nikolajewsk”; L. Graeser leg.; depository unknown. At first stored in coll. W. Koltze, current depository presumably SDEI, but needs confirmation.
Taxonomic history and synonymies



Diagnosis
Upper side bronze, bluish, or purplish, flat-lustrous, habitus similar to P. sericea, antennomeres reddish basally and darkened apically, pronotum with flattened anterior tubercles, femora reddish on basal half and dark on apical half. Aedeagus with a conspicuous elongated peak.
Description
Size : 6.7–9.7 mm.
Colour : Bronze or dark with bluish or purplish lustre.
Antennae : Slender, annulated, antennomeres basally rufous, apical dark or metallic, A2 < A3 < A4.
Pronotum : Almost quadratic or slightly longer than wide, anterior tubercles flattened, disc coarsely and closely punctate with fine wrinkles, median groove narrow or indistinct, in posterior part short and slightly deeper, then forked into two horizontal grooves near the bottom line.
Elytra : Oblong, with shallow impressions, coarsely and densely rugose on most of surface, punctures regular, strong, and deep, interstices wrinkled, interstices ~ 2–4× puncture diameter.
Legs : In most specimens the femora are reddish on basal half and dark metallic on apical half, tibia reddish with dark parts, tarsomeres dark or with reddish basal part. Some specimens with entirely reddish legs. Metafemora with prominent, thorn-shaped tooth in most specimens.
Pygidium of females with an apical notch, males broadly emarginate.
Aedeagus
: Median lobe with a conspicuous elongated peak, cap of tegmen with a deep apical notch (Fig.
Two similar species are Plateumaris sericea and P. shirahatai which differ: in P. roscida the pronotum and its tubercles are flattened, legs with large reddish parts, and the aedeagus has a conspicuous elongated peak.
Biology
Distribution
East Palaearctic: East of Lake Baikal to Far East, the Sakha (Yakutia) Republic, Amur region in Russia and north-east China (Harbin, Heilongjiang). Records exist for Asia: northern China (Heilongjiang, Inner Mongolia) [new in PalCat], Russia (East Siberia, Far East).
New country records additional to Silfverberg (2010)
China – Heilongjiang • 1 ex.; Harbin; 3 Jul 1952; E. Geiser 2021 det.; BM1953-715, (BMNH). • 2 ex.; Xinkai (Khanka) Lake, Bathing beach area; 45°21'52"N, 132°18'55"E; 11 Jun 2018; among strandline detritus; R.B. Angus, F.L. Jia, Z.L. Liang leg., E. Geiser 2021 det.; BMNH.
China: Heilongjiang and Inner Mongolia:
Material examined
More than 30 specimens from different localities throughout the distribution area.
Plateumaris rustica
Donacia rustica Kunze, 1818: 31.
Donacia abdominalis Bedel, 1891: 218 [nomen nudum].
Donacia affinis Kunze, 1818: 37.
Plateumaris rustica ab. forojulensis Gortani, 1906: 20 [infrasubspecific name].
Donacia fusca Zschach, 1788: 27 [nomen oblitum].
Donacia pallipes Kunze, 1818: 35.
Plateumaris rustica var. picipes Weise, 1898: 180.
Donacia planicollis Kunze, 1818: 34.
Donacia rustica Schüppel, 1818: 31 [nomen nudum].
Plateumaris sulcifrons Weise, 1900: 267 [syn. nov.].
Type locality
Germany, surroundings of Berlin [Kunze, 1818: 31: “in der Gegend von Berlin”].
Type material
Type specimens missing.
Taxonomic history and synonymies
Some authors, like
The names P. abdominalis Bedel, 1891 and P. (Donacia) abdominalis Olivier, 1795 [1800 is correct, see above for P. bracata] were erroneously attributed to P. rustica. The name P. abdominalis is frequently mentioned as a synonym for P. rustica or P. affinis as occurs in the key by Jacobson (1892): “abdominalis Oliv.” with P. affinis as its synonym.
Donacia affinis was also described in
Plateumaris forojulensis was described by
Donacia fusca was regarded as synonymous with P. affinis, but it is a nomen oblitum (
Plateumaris pallipes was assigned as a synonym of P. affinis and P. planicollis as a synonym of P. rustica. As the original descriptions of
Plateumaris picipes was described by
Diagnosis

Description
Size : 6.5–9.0 mm.
Colour : Upper side bronze or black with greenish, bluish, or purplish metallic lustre, colour of pronotum and elytra mostly the same but can also differ significantly. Antennae and legs entirely or partly reddish brown.
Head : Frons with deep or shallow groove, longitudinal calli distinctive or flattened.
Antennae : Filiform, each antennomere yellow or reddish at the basis, darkened at the apex, extent of darkened zone very variable, 2nd antennomere 2–3× smaller than other antennomeres which are approximately equal in length, only the 3rd antennomere is sometimes slightly smaller than the others: (2–3)× A2 = A1 = A4 … A11; A3 ≤ A4.
Pronotum : Almost quadratic, only at the basis slightly constricted, with flat disc and indistinct anterior tubercles; surface shiny or alutaceous, disc smooth with small shallow dots, more or less densely dotted, median line varies from imperceptible to distinctive.
Elytra : Punctures very delicate, interstices with slight transversal rugae, interstices 2–4× puncture diameter. Ratio of elytral length to width: 1.7–2.0.
Legs : Yellow reddish, sometimes partly or almost entirely darkened, piceous, but always with reddish joints (var. picipes Weise, 1898). Femora basally broad, metafemoral tooth very variable, mostly prominent, in some (mostly female) specimens very small or imperceptible. There is no geographic correlation concerning the size of the tooth.
Aedeagus
: Median lobe distinctly elongated, apex acute (Fig.
There are two similar species. Plateumaris consimilis has the pronotum distinctly cordate and the disc is not flattened. The pronotum of P. weisei is trapeziform and slightly longer than wide. In the territories where their distribution areas are overlapping (European part of Russia) it can be distinguished from P. weisei by the quadratic shape of the pronotum.
Also, the aedeagi of these species are clearly different (compare Fig.
Biology
The larvae are oligophagous on Carex sp. and other Cyperaceae. Adults feed on leaves and stems, not on pollen (
Distribution
West Palaearctic region: throughout Europe, further in Algeria, Turkey, Iran, and west Siberia. Records exist for Europe: Austria, Belarus, Belgium, Bosnia-Herzegovina [new in PalCat], Bulgaria, Croatia, Czech Republic, Denmark, Estonia, Finland, France, Germany, Great Britain, Hungary, Ireland, Italy, Crimea [new in PalCat], Latvia, Liechtenstein, Lithuania, Luxembourg, Montenegro [first record], The Netherlands, Norway, Poland, Romania, Russia: northern, central, and southern parts [new in PalCat] of European Russia), Serbia [new in PalCat], Slovakia, Slovenia, Spain, Sweden, Switzerland, Ukraine.
North Africa: Algeria.
Asia: Iran [first record], Russia (west Siberia), Turkey [new in PalCat].
New country records additional to Silfverberg (2010)
Bosnia-Herzegovina:
Crimea: Listed in Catalogue: Beetles of the Krym (pers. comm. S. Mosiakin 2019).
Montenegro • 11 ex.; Poljane north-west of Podgorica “Pojane”; P. rustica E. Geiser 2019 det.;
Russia • 1 ex.; Southern European territory, town Samara Nikolayevsky Uyezd; May 1916; Bostanzhoglo leg.; Zoological Museum of Moscow State University, Russia. Remark: private record by Bieńkowski 2016.
Serbia:
Iran • 1 ex.; “Persien, Elbrus Gebirge” [Elbrus mountains]; Donacia affinis H. Goecke det., Donacia rustica E. Geiser 2019 det.;
Turkey • 25 ex.; Bolu Province, Abant Daği [mountain], Abant Gölü [lake]; 1298 m a.s.l.; 31 May 1999; J. Voříšek and J. Kodada leg.; P. rustica E. Geiser 2021 det.; BMNH [ex coll. J. Voříšek]. Bolu province and Kahramanmaraş province (
Remarks
Plateumaris rustica was unknown from Turkey. There was no record in the “Checklist of leaf beetles of Turkey” (
Material studied
More than 200 specimens throughout the West Palaearctic region.
Plateumaris sulcifrons , syn. nov.
Type locality
Turkey, Kahramanmaraş province: Süleymanlı “Zeytun” [old name].
Type material
Holotype : Turkey • 1 ♀; Kahramanmaraş province, Süleymanlı “Zeytun” [old name]; 37°53'N, 36°50'E; O. Staudinger leg. Probably collected in 1872 (see below for details). Depository unknown.
Remarks
Besides, there was a confusion about the locus typicus.
Otto Staudinger (1830–1900) was a German entomologist who went on numerous collecting trips or promoted them, but insects were not collected in Malta, neither on his own journeys nor on his commissioned trips. In 1872 he visited the Cilician Taurus (
Unfortunately, the first description does not indicate where these specimens are stored. It is unlikely that Weise returned the type(s) to Staudinger, who was then working on his Lepidoptera Catalogues in his last years. Weise’s private collection and especially the Chrysomelidae part are stored in the Museum für Naturkunde in Berlin (ZMHB), but no Plateumaris specimen labelled “sulcifrons” could be found there, despite the search by J. Frisch in 2019. I screened the Plateumaris collection in 2023 but found no specimen that could be considered the type.
Like other species, where only the type specimen is known, P. sulcifrons was suspected to be a synonym of a well-known species. Weise wrote that it is similar to P. rustica and P. affinis.
Unfortunately, I could not examine the type specimen. To confirm the arguments of
The characters which should distinguish P. sulcifrons from P. rustica are either the same or within the variation range of P. rustica. Therefore P. sulcifrons is a synonym of P. rustica. This was also mentioned in
Comparison of the characters of Plateumaris sulcifrons (as described by
| Characters of P. sulcifrons | Characters of P. rustica with comments |
|---|---|
| Length: 8–9 mm | Length of P. rustica: 6.5–9.0 |
| Description based on female specimens | Females are usually larger than males in Plateumaris species, therefore 8–9 mm matches very well |
| Slightly convex | Same as P. rustica |
| Upper side bronze-coloured, below jet-black, silky grey | The same colours occur in some specimens of P. rustica |
| Antennae, tibiae and tarsomeres dark reddish | Same as P. rustica |
| Frons with a wide and deep groove | In P. rustica frons with deep or shallow groove, narrow or broad, longitudinal calli of every side of the groove distinctive or flattened |
| Prothorax square, very finely pubescent, very slightly constricted before the base, disc almost flat, shiny, rather densely punctured, middle groove anterior and posterior deepened impressed, the tubercles on both sides almost imperceptible, slightly smooth | These are typical characters of a pronotum of P. rustica, the median groove varies from imperceptible to distinctive |
| Elytra with dotted stripes, interstices narrow | Same as P. rustica |
| Femora unarmed | Tooth of metafemur mostly prominent, in some female [sic!] specimens very small or imperceptible |
| Similar to P. rustica and P. affinis but more elongated | Without type specimens it is impossible to estimate what means “more elongated” as the ratio length to width varies in specimens of P. rustica |
Plateumaris sericea
Leptura sericea Linnaeus, 1758: 397.
Donacia aenea Olivier, 1791: 292.
Donacia armata Paykull, 1799: 194.
Donacia asiatica Faldermann, 1837: 322.
Donacia sericea var. atropurpurea Westhoff, 1882: 256.
Plateumaris caucasica Zaitzev, 1930: 111 [syn. nov.].
Plateumaris discolor discolor f. coelicolor Bechyné, 1945: 89 [infrasubspecific name].
Donacia comari Suffrian, 1846: 84.
Plateumaris discolor discolor f. cupraria Bechyné, 1945: 89 [infrasubspecific name].
Donacia discolor Panzer, 1795: 216.
Donacia festucae Fabricius, 1792: 116.
Donacia geniculata C. G. Thomson, 1866: 123.
Plateumaris imitatrix Apfelbeck [nomen nudum].
Plateumaris intermedia Apfelbeck, 1912: 239.
Plateumaris discolor kratochvili f. isocoelicolor Bechyné, 1945: 89 [infrasubspecific name].
Plateumaris discolor kratochvili f. isocupraria Bechyné, 1945: 89 [infrasubspecific name].
Plateumaris discolor kratochvili f. isolacordairei Bechyné, 1945: 89 [infrasubspecific name].
Plateumaris discolor kratochvili f. isopurpuricena Bechyné, 1945: 89 [infrasubspecific name].
Plateumaris discolor kratochvili f. isoviolacea Bechyné, 1945: 89 [infrasubspecific name].
Plateumaris discolor kratochvili Bechyné, 1945: 89.
Donacia lacordairii Perris, 1864: 300.
Donacia laevicollis C. G. Thomson, 1866: 125.
Plateumaris sericea ab. levigata Csiki, 1953: 120 [infrasubspecific name].
Donacia sericea var. luctuosa Westhoff, 1882: 256.
Donacia micans Panzer, 1795: 9.
Plateumaris discolor var. nigrita Schilsky, 1908: 603.
Plateumaris nipponensis Nakane, 1963: 18.
Donacia nymphaeae Fabricius, 1792: 116.
? Plateumaris obsoleta Jacobson, 1894: 243.
Donacia palustris Schilling, 1838: 99 [homonym].
Donacia proteus Kunze, 1818: 23.
Plateumaris discolor discolor f. pseudoviolacea Bechyné, 1945: 89 [infrasubspecific name].
Plateumaris discolor discolor f. purpuricena Bechyné, 1945: 89 [infrasubspecific name].
Donacia sibirica Solsky, 1871: 245.
Plateumaris socia S.-H. Chen, 1941: 9.
Plateumaris slovacica Balthasar [nomen nudum].
Plateumaris discolor ab. tatrica Balthasar, 1934: 130 [infrasubspecific name].
Donacia sericea var. tenebricosa Westhoff, 1882: 256.
Donacia violacea Hoppe, 1795: 44 [homonym].
Plateumaris sericea ab. viridis Csiki, 1953: 120 [infrasubspecific name].
Type locality and type material
Because Linnaeus described sericea (Leptura) in 1758 no type specimen was designated. He stated that it “occurs in Europe” which is correct.
Remarks
Plateumaris sericea exhibits the highest variability in colour among Donaciinae. The upper side colour is metallic and can be green, golden green, blue, purple, red, violet, bronze, black and all shades in between. This is one of the causes so many “variations” were described which were often used like subspecies names. Additionally, throughout the whole distribution area, some specimens show a reddish base at the antennomeres. Also, few specimens exist with a reddish part near the joints of the femora, tibiae or tarsomeres. In (most) identification keys P. sericea is characterised by “antennae and legs entirely metallic“, which is usually correct. Only in recent keys it is mentioned that there can also be reddish parts at some joints. Therefore, these “not entirely metallic” specimens supported the idea that specimens with a reddish spot belong to a different species or at least subspecies. I examined many specimens from the whole distribution area and determined their morphologic characters inclusive of the aedeagus shape are within the variety range of P. sericea.
In large European collections, where Asian specimens are stored, many of these Asian specimens show a red base of their antennomeres. Perhaps, these specimens were preferentially collected and stored whereas “entirely metallic“ specimens were considered as common and not worth keeping.


Schematic sketch of the endophallus. Median ejaculatory guide blackened A Plateumaris sericea (and specimens identified as P. discolor) with apex of median ejaculatory guide rounded. The endophalli of specimens from Poland, Italy and Japan all look the same as in this sketch B Plateumaris shirahatai with apex of median ejaculatory guide notched (drawings by G. Geiser based on photographs from
Taxonomic history and synonymies
The correct data of the first description is Leptura sericea (Linnaeus, 1758): 397, and not “Linnaeus, 1760: 196” as it was printed in
The genus name Donacia was erected later in 1775 by Fabricius. There he described Donacia crassipes and Donacia simplex and assigned Leptura aquatica L., 1758 to the genus Donacia, but, significantly, he did not change the genus name of Leptura sericea.
Plateumaris asiatica was described as Donacia asiatica by
Plateumaris caucasica Zaitzev, 1930: syn. nov., see below.
Plateumaris discolor kratochvili f. coelicolor was described by
Plateumaris discolor discolor f. cupraria was described by
Plateumaris discolor (Panzer, 1795): confirmed synonym, see below.
Plateumaris imitatrix: This name with the author “Apfelbeck” can be found on several museum specimens from Bosnia-Hercegovina (
Plateumaris intermedia was described by
Plateumaris discolor kratochvili was described by
Plateumaris discolor kratochvili forma isocoelicolor, also the forma isocupraria, forma isolacordairei, forma isopurpuricena and forma isoviolacea were described by
Plateumaris levigata was described by Csiki in 1953 as an aberration of P. sericea.
He wrote: “Plateumaris sericea ab[erratio]. levigata[sic!] nom[en]. nov[um]. pro violacea Gyll. (nec Pall., nec Hoppe)“. Plateumaris “levigata” is not a spelling error although “laevigata” is more common, but both spellings were used in classical Latin for the word “smoothed”, so “levigata” is correct. Anyway, this is an infrasubspecific name.
Plateumaris micans was described as Donacia micans by Panzer in 1795 and not in 1796 according to
Plateumaris nipponensis was described by
Plateumaris nymphaeae:
Plateumaris obsoleta Jacobson, 1894: questionable synonym, see below in P. shirahatai.
Plateumaris palustris was described as Donacia palustris by Schilling in 1838 on page 99 and not in 1837 on page 104. It is a homonym because
Plateumaris discolor kratochvili forma pseudoviolacea and forma purpuricena were described by
Plateumaris sibirica (Solsky, 1871), confirmed synonym, see below.
Plateumaris socia was described by
Plateumaris sericea slovacica Balthasar: In the coll. generalis in
Plateumaris discolor ab. tatrica was described by
Plateumaris violacea was described by
Csiki described 1953 “Plateumaris sericea ab[erratio]. viridis nom[en]. nov[um]. pro micans Panz. (nec Hoppe)“, but that does not matter because this is an infrasubspecific name.
General remarks on the morphology and distribution of Plateumaris sericea
Plateumaris sericea has the largest distribution area of all Plateumaris species. It is no surprise that it is also genetically very variable (
Diagnosis
Legs and antennae usually entirely metallic, some specimens with reddish parts near the joints; pygidium of females rounded, in some specimens slightly emarginate, that of males emarginate; apex of median ejaculatory guide rounded.
Description
Size : 6.5–10.5 mm.
Colour : Plateumaris sericea shows the greatest colour variety among all Donaciinae species: The whole beetle can be bronze, green, blue, black, cupreous, purple, red, yellow, and all shades in between. Antennae and legs are mostly completely metallic, but there are some specimens with red base of the antennomeres and even with red parts of the legs, usually at the tibiae or tarsomeres.
Head : Same colour as pronotum, supraocular furrow indistinct; vertex with a median line, antennomeres always apically darkened, either completely dark metallic or the basal part reddish to varying degrees, A3 slightly longer than A2, A4 2× as long as A2 in most specimens. A3 ≥ A2, A4 = 2× A2.
Pronotum : Outline more or less quadrate, in some specimens longer than wide; anterolateral tubercles prominent but sometimes flattened, the disc varies from alutaceous and impunctate to finely or coarsely punctate with deep transverse wrinkles, the median line can be clear and deep or only a very shallow furrow.
Elytra : Disc rugose, rows of punctures, shape, and apex typical like in other Plateumaris species.
Legs : Entirely metallic and same colour as upper side. Rarely, some specimens show reddish parts near the joints, mostly on the tibiae or tarsomeres, metafemora of most specimens with a prominent, blade-like tooth, but some specimens with an indistinct or without any tooth.
Pygidium : Emarginate in males, usually rounded but sometimes shallowly emarginate in females.
Aedeagus
: Examples of its variability are shown in Fig.
There are no reliable external characters to distinguish P. sericea from P. shirahatai. The only reliable feature can be found at the endophallus. The apex of the median ejaculatory guide of the endophallus is notched in P. shirahatai (Fig.
Biology
Plateumaris sericea feeds on Carex sp., Juncus sp., Eriophorum sp., Scirpus sp. and related plant species. For details and identification of the larvae see
Distribution
Plateumaris sericea has not only the largest distribution area of all Plateumaris species but also of all Donaciinae species. It occurs in the whole Palaearctic region. Any lack of records in some parts of its area is most probably due to a lack of collection trips there. Records exist for Europe: Austria, Belgium, Bosnia-Herzegovina [new in PalCat], Belarus, Bulgaria, Croatia [first record], Czech Republic, Denmark, Estonia, Finland, France, Germany, Great Britain, Greece [first record], Hungary, Ireland, Italy, Latvia, Liechtenstein, Lithuania, Luxembourg, Crimea [first record], Montenegro [first records], The Netherlands, North Macedonia [first record], Norway, Poland, Romania, Russia (north, central, and south parts of European territory), Serbia [new in PalCat], Slovakia, Slovenia, Spain, Sweden, Switzerland, Turkey [new in PalCat], Ukraine.
North Africa: Algeria [new in PalCat].
Asia: Armenia, Azerbaijan, China (Beijing, Hebei, Zhejiang), Georgia, Iran, Japan, Kazakhstan, Mongolia, North Korea [new in PalCat], Russia (west, east, and south Siberia [new in PalCat], Far East), South Korea [new in PalCat], Turkey [new in PalCat].
New country records additional to Silfverberg (2010)
Bosnia-Herzegovina:
Bosnia • 1 ex,; Livno [north of Buško Jezero]; collected from Cladium mariscus; Donacia imitatrix Apfelbeck [V. Apfelbeck det.], Plateumaris sericea E. Geiser 2019 det.;
Croatia • 1 ex.; Pakrac [town in western Slavonia]; Plateumaris discolor ab. lacordairii Z. Kaszab det., Plateumaris sericea E. Geiser 2020 det.,
Crimea • 1 ex.; Sebastopol; W. Pliginsky [leg.?]; Plateumaris discolor ab. lacordairii W. Balthasar [det.?], Plateumaris sericea E. Geiser 2020 det.; SMNC. Remark:
Greece • 1 ex.; Thessalia, Pindos mountains, Dessi, Kalambaka, Pertouli, 1110 m; 23. May 2001; A. & F. Riedel leg.; E. Geiser 2023 det; SMNS.
Montenegro • 1 ex.; Crna Gora, Žabljak; 18. Jul 1934; Dr. J. Fodor leg.; J. Bezděk 2017 det.;
North Makedonia • 6 ex.; Delčevo; 3 Jun. 1982; I. Rozner leg.; J. Bezděk 2017 det.;
Serbia: Gavrilovic and Curcic (2011).
Turkey: Many records from European and Asian territory in
Algeria:
North Korea:
South Korea:
Russia (South Siberia):
Material examined
More than 500 specimens from different localities, labelled as various species or subspecies throughout the distribution area.
Plateumaris caucasica , syn. nov.
Type locality
Russia: Stavropol and Dagestan.
Type material
Type series : Russia • 4 ex; Ciscaucasia, Stavropol; Apr 1905; DM Maljuzhenko leg.; Russia • 5 ex; Daghestan, Chasav-jurt; E. Koenig leg.
Remarks
According to
As I explained in “General remarks on synonyms of Plateumaris sericea” above, this is a typical example of establishing a new “species” on colour characters. The other mentioned “different” characters are completely within the variation range of P. sericea or characteristic of this species. The morphology of the aedeagus is also completely within the variation range of P. sericea. In the same area also typical P. sericea (that is: with completely metallic antennae and legs) could be found, the colour variation form “P. sericea caucasica” could not even be a subspecies.
Zaitzev assumed that Plateumaris caucasica is also “very close” to P. annularis, because both have a red base at their antennomeres and legs which are partly reddish brown. To the credit of Zaitzev it is necessary to mention that he had doubts if P. caucasica is really a new species or a synonym to P. annularis. He suggested to treat it as a new species until further knowledge is available about the East Siberian Plateumaris species.
Plateumaris annularis was synonymised by
First, it is actually biogeographically implausible that these two species should be synonyms. Second, the pygidium is emarginate in P. roscida and not emarginate in P. caucasica in both sexes. Third, the aedeagi of P. roscida and P. caucasica are strikingly different. For the median lobe of P. roscida see Fig.
Material examined
More than 30 specimens from the Caucasus region (north and south) labelled as “P. caucasica”, “P. sericea caucasica” or “P. roscida” which were all clearly P. sericea.
Plateumaris discolor
Type location
Germany.
Type material
The holotype is unknown.
Remarks
Plateumaris discolor was described by
Several characters are used to distinguish P. discolor from P. sericea. The first are the antennomeres: in P. discolor A3 and A4 are a little bit longer than A2, whereas in P. sericea A3 is 1.5× as long as A2 and A4 is twice as long as A2. In fact, the length of the antennomeres is very variable, therefore the difference between “a little bit” and “one and a half” is not clear.
The second is the pronotum disc: in P. discolor it is more punctured and transversely wrinkled whereas in P. sericea it is very finely sculptured. However, the structure of the pronotum disc varies in both “species” in its sculpture and shows an intermediary form in many cases.
The third is the median lobe of the aedeagus, which is also very variable (Fig.
Due to these variations, there are no reliable morphological characters to distinguish P. discolor from P. sericea. Other evidence suggests that they may be separate species: P. discolor is reported to be assigned to acid soil and peat bogs where the larvae develop on Carex, Juncus and related plants, whereas P. sericea prefers various wetland habitats with alkaline soil (
Molecular studies by
Material examined
More than 100 specimens labelled “P. discolor” from different localities in Europe.
Plateumaris sibirica
Type locality
Russia, Irkutsk.
Type material
Remarks
Plateumaris sibirica was described as Donacia sibirica by
Eventually, the original description only mentions characters which are typical for P. sericea. It has been regarded as a synonym to P. sericea by
I examined more than 60 specimens identified as P. sericea sibirica, mainly from the BMNH,
The original description was mostly cited as Solsky (1872). It was described in “Horae Societatis Entomologicae Rossiae” volume 8 comprising the years 1871 and 1872. There it was described in the part of 1871 according to
Plateumaris shirahatai
Plateumaris shirahatai Kimoto, 1971: 1.
Plateumaris macropenis Nakane, 1999: 45.
? Plateumaris obsoleta Jacobson, 1894.
Type localities
Plateumaris shirahatai: Japan, Honshu, Yamagata Prefecture, Shizu, Gassan; Plateumaris macropenis: Japan, Honshu, Oze.
Type material
Holotype of P. shirahatai: Japan • 1♂, Yamagata Prefecture, Shizu, Gassan; 17 Jun 1960; K. Shirahata leg.; Entomological Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka. The holotype was not examined.
Paratype. Japan • 3 ♀; same data as for the holotype; Japanese Insect Collection No. 21963,
Holotype of P. macropenis. Japan • 1 ♀; Honshū, Oze; 15 Jul. 1950; H. Hasegawa leg.; Plateumaris macropenis T. Nakane det.; Laboratory of Systematic Entomology, Faculty of Agriculture, Hokkaido University, Sapporo, Japan.
Taxonomic history and synonymies
Plateumaris macropenis Nakane, 1999 was synonymized by
? Plateumaris obsoleta Jacobson, 1894: see below.
Diagnosis
Pronotal disc rugose, antennae, and legs entirely metallic, although in some specimens the basis of the antennomeres is reddish, A3 = 1.5–2× A2, tooth on metafemur sharp blade-like or obtuse, pygidium of females rounded, in some specimens slightly emarginate, pygidium of males emarginate or truncate, median process of endophallus notched.

Description
Size : Males 6.5–7.3 mm, females 7.8–8.2 mm.
Colour : Upper side colour very variable: blackish, blue, green, bronze, cupreous, purple, same colours as P. sericea. Antennae and legs same colour as upper side, in some specimens with reddish parts near the joints.
Head : Rugulosely punctate and pubescent, frontal calli convex, interocular area with a longitudinal median furrow.
Antennae : Entirely metallic with same colour as dorsum, antennomeres in some specimens basically reddish, filiform, A1 robust, club-shaped, A1 = 2× A2, A3 ≅ 2× A2, A3 < A4 ≤ A5.
Pronotum : Slightly longer than broad, gradually narrowed posteriorly, dorsal surface with a pair of distinctly raised antero-lateral tubercles, and with a triangular depression medio-basally, disc punctate with transverse rugae, median line indistinct, shallowly furrowed.
Elytra : Interstices of the rows of punctures with close oblique or transverse corrugations and showing a rugged appearance.
Legs : Entirely metallic, same colour as dorsum, in some specimens small reddish parts at the base of the joints, tooth of metafemur prominent and blade-like but also in some specimens obtuse.
Pygidium : Apex pubescent, apical shape in females rounded, in some specimens slightly emarginate, in males emarginate or truncate.
Male genitalia
: Median lobe of aedeagus (Fig.
Remarks
The only reliable feature to distinguish P. shirahatai from P. sericea is the notched apex at the median ejaculatory guide of the endophallus. The habitus of P. shirahatai looks also very similar to P. roscida, but the latter always has large red parts on the legs and antennae, and their aedeagi are remarkably different (Figs
Biology
The larvae feed on Carex sp. (Narita, 2003). Adults were collected on the florescence of Carex sp. (
Distribution
East Palaearctic species. The distribution area of P. shirahatai is situated completely within the eastern area of P. sericea. Unfortunately, it is almost impossible to distinguish P. shirahatai from P. sericea without male genitalia. Both species share the same colour spectrum and same variation of the other external characters.
Records exist for Asia: China (Jilin) [new in PalCat], Japan (Hokkaido and Honshu), Mongolia [new in PalCat], Russia (Far East: Primorsky Krai, Sakhalin), South Korea, South Kuril (Etorofu).
New country records additional to Silfverberg (2010)
China (Jilin):
Mongolia: • 2 ♂♂, 1 ♀; central Mongolia, Terelj; 47°59'24"N, 107°27'E; 28 Jun 2004; M. Hayashi leg.; M. Hayashi det.; Hoshizaki Institute for Wildlife Protection, Izumo, Japan. Remark: Some parts of this data are published in
This recent record from central Mongolia shows clearly that the distribution area is not known until now. As P. shirahatai was described only in 1971 it is likely that some specimens from the East Palaearctic stored in collections may be identified as P. sericea.
Material examined
20 specimens from Hokkaido and Honshu.
Plateumaris obsoleta
Type locality
Russia, Far East, Primorsky Krai, Bay of Posyet.
Type material
Holotype
: Russia • 1 ♀; Far East; Primorsky Krai; Bay of Posyet;


Remarks
At first, I intended to synonymise P. obsoleta with P. sericea based on studies of the type material and description, but doubts remained that it is more likely that P. obsoleta is a synonym with P. shirahatai. I am sure that P. obsoleta, described based on one female specimen and never recorded again in more than 100 years, is a synonym. However, I cannot prove if it belongs to P. sericea or to P. shirahatai because it is impossible to distinguish these two species by external morphological characters. These two species differ only by subtle morphological differences in the apical part of the endophallus (Fig.
Plateumaris obsoleta was described by Jacobson (1894) (see

The controversy about the sex of this specimen can now be solved: the apical part of the ovipositor protrudes, which Jacobson misinterpreted as a part of the aedeagus. Although
Perhaps it will be possible in the near future to solve this problem without destroying this single specimen with more elaborate methods than historical DNA analysis. The solutions used to extract the DNA may be destroy the connecting membranes between the chitinous parts. Nowadays, nobody can guarantee that this specimen would NOT be damaged! Additionally, it is very questionable whether the results will be clear enough. Usually, the DNA in old, stored insects is fragmented and cannot be sufficiently reconstructed to make the decision to which species the specimens belong. Plateumaris sericea and P. shirahatai are closely related, which was proofed by DNA analysis (
Plateumaris weisei
Donacia weisei Duvivier, 1885: cxvi.
Donacia borealis Mannerheim [nomen nudum].
Plateumaris hirashimai Kimoto, 1963: 13.
Donacia (Plateumaris) mongolica Semenov, 1895: 267.
Plateumaris morimotoi Kimoto, 1963: 13.
Plateumaris consimilis orientalis Shavrov, 1948: 49.
Plateumaris sachalinensis L. N. Medvedev, 1973: 876.
Type localities
Plateumaris weisei: Siberia. Original label text: “Sibérie coll. Duvivier“; Plateumaris consimilis orientalis: Far East, Vladivostok, Sedanka, Russia; Plateumaris hirashimai: Hokkaido, Ashoro in Tokachi, Japan; Plateumaris mongolica: North Mongolia, Borcha-Urga, Mongolia; Plateumaris morimotoi: Hokkaido, Tenninkyo Mt. Daisetsu, Japan; Plateumaris sachalinensis: Far East, Sakhalin, Yuzhno-Sakhalinsk, Russia.
Type material
Type of Plateumaris weisei: 1 syntype, Siberie coll. Duvivier; Museum Paris coll. H. Clavareau 1932, vid. I.S. Askevold 1984 (
Type series of P. consimilis orientalis: Russia • 3 ♀; Far East, Vladivostok, Sedanka; 19 Jun. 1937 [present depository unknown].
Type of P. hirashimai: Japan • 1 ♀; Hokkaido, Ashoronuma in Tokachi; 28 Jul. 1949; R. Matsuda leg.; collection Entomological Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka, Japan.
Type of P. mongolica: Mongolia • 1 ♂; valley of the river Borcha, from Urga to the East; 6 Jul. 1894, B. Kaschkarow leg.; collection Semenov [present depository unknown].
Type of P. morimotoi: Japan • 1 ♂; Hokkaido, Tenninkyo Mt. Daisetsu; 27 Jul. 1955; K. Morimoto leg.; collection Entomological Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka, Japan.
Type of P. sachalinensis: Russia • 1 ♂; Far East, Sakhalin, Yuzhno-Sakhalinsk; 12 Jul. 1955; collection of N.N. Filippov [present depository unknown]. Paratype: Russia • 1 ♂; Far East, Sakhalin, Yuzhno-Sakhalinsk; 10 Jul. 1955; [red label:] Paratype Plateumaris sachalinensis Medvedev, Plateumaris weisei Duv. E. Geiser 2021 det.;
The photographs of the syntype of P. weisei and the paratype of P. sachalinensis were examined.

Taxonomic history and synonymies
This list of synonyms and their shifting positions (see below) indicate the main systematic problems with P. weisei. First, it is difficult to distinguish it from other Plateumaris species. The variety of conspicuous morphological characters (colour, relative length of antennomeres, shape and structure of pronotum, shape of metafemoral tooth, etc.) overlap with other species. Second, the locality name in the first description “Siberie” is anything but precise. Third, P. weisei has a particularly wide distribution range, from northern Fennoscandia through European Russia and Siberia to Far East, Mongolia, Northern China, the Korean peninsula, and Japan. Altogether this resulted in the new descriptions of Plateumaris species when a specimen was found outside Siberia with slightly different characters.
Donacia borealis (Mannerheim), nomen nudum: the first who recognised that a specimen of still undescribed Plateumaris weisei belonged to a new species was Carl Gustav Mannerheim, a Finnish entomologist (1797–1854). He labelled a specimen (possibly more than one specimen, but I only found this one) from “Lapponica” with “Donacia borealis”, which clearly is P. weisei (vid. E. Geiser 20 Jul. 2022). This specimen is stored in the coll. Mannerheim (
Plateumaris hirashimai was first described by
Donacia mongolica was described by
Plateumaris morimotoi was first described by
Plateumaris consimilis orientalis was described by
In the coll. Frey (
Plateumaris sachalinensis was described by
After the study of the original description
Diagnosis
Pronotal disc finely rugose and punctured, sometimes with microsculpture, median line obsolete, sometimes shallowly furrowed (similar to P. shirahatai), metafemur with a small, not blade-like tooth, usually rufous at the base; aedeagus with apex of median lobe arced on both sides, gradually narrowed apically, without a median lip.
Description
Size : Males 6.2–7.0 mm, females 6.8–8.0 mm.
Colour : Most specimens dorsally cupreous or bronze, sometimes metallic green, blue, purple, or non-metallic brown.
Head : Eyes small, supraocular furrow indistinct, vertex pubescent with deep median line, antenna entirely rufous in most specimens but in some specimens darkly rufous or apically metallic, antennomeres: A5 longest in second to A6 and ca 3.5× as long as wide, A4 ca 2.2× as long as A2.
Pronotum : Outline subquadrate, slightly longer than wide, basal part narrowed, slightly cone-shaped, anterior tubercles distinctly visible or almost entirely smooth, disc more or less punctate, rugulose, median groove indistinct or shallowly furrowed.
Elytra : Transverse rugae between the rows of punctures, especially on interstices 1–4.
Legs : Yellow-reddish, in some specimens more or less darkened, femur, tibia, and tarsomere pubescent, outer apical angles of pro- and mesotibiae with a spine, outer apical angles of metatibiae with a small spine, metafemoral tooth mostly blunt or moderate.
Aedeagus
: Apex of median lobe arced on both sides, gradually narrowed apically, without a median lip, cap of tegmen rounded at apex (Fig.
The main different features between P. amurensis and P. weisei are shown in Table
Biology
Larvae were found at the roots on Carex sp. (
Distribution
Plateumaris weisei is a Trans Palaearctic species, it occurs from northern Fennoscandia through Siberia to the Far East, northern China, the Korean peninsula, and Japan. Also, it occurs in a broad span of latitudes, from the arctic polar circle (67°N) to 35°N in Korea. Records exist for Europe: Sweden, Finland, Russia (north and central part of European Russia).
Asia: China (Heilongjiang [new in PalCat], Inner Mongolia), Japan (Hokkaido), Mongolia, Russia (west, east, and south Siberia [new in PalCat], Far East), South Korea [new in PalCat].
In Japan records exist only from Hokkaido so far (
New country records additional to Silfverberg (2010)
China • 1 ex.; Heilongjiang, “Manchuria“ Harbin; Plateumaris weisei E. Geiser 2020 det.; SDEI [coll. K.-H. Mohr]. Remarks:
Russia • 5 ex.; South Siberia, Angara (near Baikal); I. Askevold 1985 det., E. Geiser 2019 vid.; Sharp-coll. 1905 – 313, BMNH;
South Korea:
Material examined
More than 80 specimens from Europe and Asia.
Discussion
The changes in
A primary objective was to declutter doubtful species and ambiguous synonymisations to answer these questions: how many species of Plateumaris exist in the Palaearctic region, what are their names, and how should the remaining 70 names used in a description for a Plateumaris taxon be allocated to the valid species? This was successful in most cases. Additionally, some holotypes could be tracked down, e.g., of P. tenuicornis, which could be identified unambiguously as synonymous with P. consimilis. This also confirmed
In general, this study largely confirmed the systematic status published by
Many problems could be solved unambiguously, but not all P. obsoleta may be a synonym of P. shirahatai: it could in fact be a synonym of P. sericea, which also occurs at the same locality and cannot be distinguished from P. shirahatai by external characters alone.
A problem does remain concerning the synonymisation of P. discolor with P. sericea. They are not distinguishable morphologically, even if some identification keys suggest that they are. The allocations to their ecological requirements and food plants are contradictory: because P. sericea shows very high genetic variability and has a very large distribution area, it may consist of cryptic species, possibly indicating evolution in progress.
I am reluctant to designate neotypes at this point. Before choosing specimens as neotypes one should be certain that the holotype or type series do not exist. Some museum collections, where I expect to find a missing holotype, could not be visited in the last years due to Covid 19, but this should be rectified in the near future. Additionally, the specimens for neotypes should be from the same location or as near as possible to where the original type or type series were collected, and this require visits to museums in which such specimens are stored.
Finally, I do not want to sweep the most severe problem in researching Palaearctic Plateumaris species under the carpet: the majority of specimens for systematic studies is stored in museums. It is not very hopeful to try to catch specimens in the field. Most museum specimens are by-catches acquired by luck: there exists specimens from expeditions which took place 100 and more years ago from large areas of Asia. From areas like Syria, Afghanistan, and large parts of China such Xinjiang Province, the only information we can access now is from museum specimens. The application of historical DNA methods seems to be helpful but is problematic: its success is questionable, mostly due to the severe fragmentation of the DNA in old specimens. Also, damaging these few very precious specimens by the extraction solution is likely. Perhaps, in the future, there will be gentler methods developed for such studies.
Even without restrictions for field studies caused by politics, in Europe, too, it is difficult to obtain new samples. Many colleagues who know that I am working on Donaciinae have tried to catch specimens during their own field trips in the last years. Besides P. sericea, all other species are rare because of many changes of the limnic environment during the last 100 years. Water pollution is not such an issue as it was 40 years ago, but man-made changes to diverse limnic habitats, especially bank straightening and drainage, has probably led to the extinction of Plateumaris populations that need specific ecological conditions to survive and thrive.
Acknowledgements
This generic revision was possible only with the help of many colleagues, particularly important in the last years with travel and entry restrictions. The colleagues responsible for the museum collections I visited are indicated in the list of museum abbreviations. Many thanks for their support during my on-site studies and their useful suggestions. Further essential help was obtained from the following colleagues: Jan Bezděk (Brno, Czechia) provided me with rare literature and his huge knowledge of tricky systematic and nomenclatorial details. I received answers from Horst Kippenberg (Herzogenaurach, Germany) and from Ron Beenen (Nieuwegein, Netherlands) whenever I asked them questions. Johannes Bengsten (Stockholm, Sweden) provided me with unpublished phylogenetic trees of P. sericea and P. discolor. I thank Klaas Reißmann (Kamp-Lintfort, Germany) for interesting discussions about P. discolor.
I also thank the following for support with images: Keita Matsumoto (London, United Kingdom), who provided me with excellent photos from specimens of the coll. Voříšek (BMNH); Alexey Moseyko (St. Petersburg, Russia), who tracked down the holotype of P. obsoleta and sent to me essential pictures and other helpful information; Peter Vdáčný, Vladimir Janský, and Martin Sečanský (all in Bratislava, Slovakia) helped to find the holotype of P. tenuicornis. From Masakazu Hayashi (Izumo, Japan) I received pictures of P. shirahatai from Primorsky Kray and other useful information, including permission to use drawings from his publications.
Gloria Geiser (Salzburg, Austria) made several drawings. Irmgard Geiser (Salzburg, Austria) helped with cryptic locality label texts written in Albanian. Helena Shaverdo (Vienna, Austria) deciphered many handwritten labels in Russian. Remigius Geiser senior (Salzburg, Austria) made meticulous corrections and improved the manuscript with tough discussions. Finally, I would like to thank the editorial team, especially Caroline Chaboo (Lincoln, Nebraska, USA) who also significantly improved the language, Yuri Mikhailov (Yekaterinburg, Russia) and two anonymous reviewers for their useful advice, and Nathalie Yonow (Swansea, United Kingdom) for the linguistic proof.
Additional information
Conflict of interest
The authors have declared that no competing interests exist.
Ethical statement
No ethical statement was reported.
Funding
The research in the
Author contributions
The author solely contributed to this work.
Author ORCIDs
Elisabeth Geiser https://orcid.org/0009-0005-4536-8938
Data availability
All of the data that support the findings of this study are available in the main text.
References
- Alonso-Zarazaga MA, Evenhuis NL (2017) Revised dating of Panzer’s Faunae insectorum Germanicae initia. Deutschlands Insecten (1792–1810), based primarily on the Leipzig Book Fair Catalogs. Sherbornia 4(1): 1–8. https://zoobank.org/References/3b5a713d-6aa8-4a3b-82d8-09505bcdb92e
- An SL (2019) Two new records of the subfamily Donaciinae (Coleoptera, Chrysomelidae) from Korea. Journal of Asia-Pacific Biodiversity 12(1): 63–65. https://doi.org/10.1016/j.japb.2018.11.005
- Anonymous [“S.“] (1901) [Obituary] Dr. Otto Staudinger †. Deutsche entomologische Zeitschrift Iris 13: 341–358. https://www.biodiversitylibrary.org/item/45903#page/363/mode/1up
- Apfelbeck V (1912) Komponente balkanske faune iz roda Chrysomelidae. Zoogeografski prinos fauni balkanskog Poluostrova. Glasnik Zemaljskog Muzeja u Bosni i Hercegovini 24: 235–263.
- Askevold IS (1990) Reconstructed phylogeny and reclassification of the genera of Donaciinae (Coleoptera: Chrysomelidae). Quaestiones Entomologicae 26(4): 600–664.
- Askevold IS (1991) Classification, Reconstructed Phylogeny, and Geographic History of the New World Members of Plateumaris Thomson, 1859 (Coleoptera: Chrysomelidae: Donaciinae). Memoirs of the Entomological Society of Canada 157(S157): 5–175. https://doi.org/10.4039/entm123157fv
- Balthasar V (1934) Ein Beitrag zur Kenntnis der palaearktischen Donaciini. Entomologisches Nachrichtenblatt (Troppau) 8(4): 128–130. https://www.zobodat.at/publikation_volumes.php?id=62568
- Bates HW (1883) Supplement to the Geodephagous Coleoptera of Japan, chiefly from the collection of Mr. George Lewis, made during his second visit, from February, 1880 to September, 1881. The Transactions of the Entomological Society of London 1883(3): 205–290. https://doi.org/10.1111/j.1365-2311.1883.tb02947.x
- Bechyné J (1942) O variabilitě Plateumaris consimilis Schrank. (Col. Donaciidae). De variatione Plateumaris consimilis Schrank. (Col. Donaciidae). Sborník entomologickèho odděleni zemědělské Musea v Praze (= Acta Entomologica Musei Nationalis Pragae) 20: 232–237. [In Czech and Latin] https://www.aemnp.eu/acta-entomologica/volume-20/482/o-variabilite-plateumaris-consimilis-schrank-col-donaciidae-de-variatione-plateumaris-consimilis-schrank-col-donaciidae.html
- Bechyné [=Bechynĕ] J (1945) Příspĕvek k poznání našich mandelinek. Additamenta ad cognitionem Phytophagorum centralieuropaeorum. Entomologické Listy 8: 87–91.
- Bedel L (1891) Faune des coléoptères du bassin de la Seine Tome V (1889–1901), 423 pp. https://www.biodiversitylibrary.org/item/276952#page/66/mode/1up
- Bezděk J, Sekerka L (in press) Catalogue of Palaearctic Coleoptera. Volume 6/2. Revised and updated Second Edition. Chrysomeloidea II (Megalopodidae, Orsodacnidae, Chrysomelidae). Brill, Leiden.
- Bieńkowski AO (2014) Reed Beetles (Coleoptera: Chrysomelidae: Donaciinae) [in Russian]. Publishing Company Mukhametov GV, Livny, 380 pp.
- Bieńkowski AO, Orlova-Bieńkowskaja MJ (2004) Morphology, systematics and host plants of Palaearctic Donaciinae larvae. In: Jolivet P, Santiago-Blay JA, Schmitt M (Eds) New Developments in the Biology of Chrysomelidae. SPB Academic Publishing, The Hague, 481–502. https://doi.org/10.1163/9789004475335_042
- Borowiec L (1984) Zoogeographical study on Donaciinae of the world (Coleoptera, Chrysomelidae). Polskie Pismo Entomologiczne 53: 433–518. https://scholar.google.at/scholar?q=Borowiec+L+(1984)+Zoogeographical+study+on+Donaciinae&hl=de&as_sdt=0&as_vis=1&oi=scholart
- Bousquet Y (2016) Litteratura Coleopterologica (1758–1900): A guide to selected books related to the taxonomy of Coleoptera with publication dates and notes. ZooKeys 583: 1–776. https://doi.org/10.3897/zookeys.583.7084
- Bousquet Y (2018) The dating of the fourth volume of Guillaume-Antoine Olivier’s “Entomologie, ou histoire naturelle des insectes”. ZooKeys 734: 137–148. https://doi.org/10.3897/zookeys.734.22901
- Böving AG (1910) Natural history of the larvae of Donaciinae. Mitteilungen aus dem biologischen Süßwasserlaboratorium Frederiksdal bei Lyngby (Dänemark). Nr. VII. Sonderabdruck aus Internationale Revue der gesamten Hydrobiologie und Hydrographie 1910: 1–108.
- Brelih S, Drovenik B, Pirnat A (2006) Material for the Beetle Fauna (Coleoptera) of Slovenia. 2nd contribution: Polyphaga: Chrysomeloidea (= Phytophaga): Cerambycidae. Scopolia, Journal of the Slovenian Museum of Natural History, Ljubljana 58, 442 pp. https://www.zin.ru/Animalia/Coleoptera/rus/lazpdf90.htm
- Chen SH (1941) Notes on Donaciinae beetles. Sinensia 12: 1–17.
- Cho HW, An SL (2020) An annotated checklist of Leaf Beetles (Coleoptera: Chrysomelidae) of Korea, with comments and new records. Far Eastern Entomologist = Dal’nevostochnyi Entomolog 404: 1–36. https://doi.org/10.25221/fee.404.1
- Chȗjȏ M (1959) Contribution to the Fauna of Chrysomelidae (Coleoptera) in Japan (III): Memories of the Faculty of Liberal Arts and Education. Kagawa University 81(2): 1–16.
- Clavareau CH (1913) Pars 53: Chrysomelidae: Megascelinae, Megalopodinae, Clytrinae, Cryptocephalinae. Chlamydinae, Lamprosominae. In: Schenkling S (Ed.) Coleopterorum Catalogus. W. Junk, Berlin, 278 pp. https://www.biodiversitylibrary.org/item/39830#page/11/mode/1up
- Csiki E (1953) Über neue und bekannte Coleopteren aus Ungarn und den angrenzenden Ländern. Annales Historico-Naturales Musei Nationalis Hungarici, Series Nova 3 [1952]: 115–135. http://publication.nhmus.hu/annales/cikkreszletes.php?idhoz=639
- Duvivier A (1885) Quatre Phytophages nouveaux. Bulletin ou Comptes-Rendus des Scéances de la Société Entomologique de Belgique 1885: cxvi–cxix. https://www.biodiversitylibrary.org/item/303035#page/120/mode/1up
- Ekiz AN, Sen I, Aslan EG, Gök A (2013) Checklist of leaf beetles (Coleoptera: Chrysomelidae) of Turkey, excluding Bruchinae. Journal of Natural History 47(33–34): 2213–2287. https://doi.org/10.1080/00222933.2012.763069
- Ekiz AN, Geiser E, Gök A, Kaya ÖD (2020) Donaciinae (Coleoptera: Chrysomelidae) of Turkey: species list and new records. Aquatic Insects 41(3): 1–13. https://doi.org/10.1080/01650424.2020.1739312
- Fabricius JC (1775) Systema entomologiae sistens insectorum classes, ordines, genera, species, adiectis synonymis, locis, descriptionibus, observationibus. Libraria Kortii, Flensburgi et Lipsiae, [xxxii +] 832 pp. https://www.biodiversitylibrary.org/page/25551417#page/213/mode/1up
- Fabricius JC (1792) Entomologia systematica emendata et aucta, secundum classes, ordines, genera, species, adjectis, synonimis, locis, observationibus, descriptionibus. Tomus I. Pars II. C. G. Proft, Hafniae, [xx +] 538 pp. https://www.biodiversitylibrary.org/item/219086#page/472/mode/1up
- Faldermann F (1837) Fauna entomologica transcaucasica. Coleoptera II. Nouvelles Mémoires de la Société des Naturalistes de Moscou 5: 1–433[, 15 pls]. [separate issue: Fauna entomologica Trans-Caucasica. Coleoptera. Pars 2. l’imprimerie d’Auguste Semen, Imprimeur de l’Academie Impériale Medico-Chirurgicale, Moscow, 433 pp, pls 1–15].
- Gavrilović BD, Ćurčić SB (2011) Diversity of species of the family Chrysomelidae (Insecta, Coleoptera) in Serbia, with an overview of previous researches. Acta Zoologica Bulgarica 63(3): 231–244. https://scholar.google.at/scholar?q=Gavrilovi%C4%87+BD,+%C4%86ur%C4%8Di%C4%87+SB+(2011)+Diversity+of+species+of+the+family+Chrysomelidae&hl=de&as_sdt=0&as_vis=1&oi=scholart
- Geiser E (2015) World checklist of freshwater Coleoptera: Chrysomelidae – Donaciinae species. Freshwater Animal Diversity Assessment (FADA). World Wide Web electronic publication. http://fada.biodiversity.be/group/show/65
- Geiser E (in press) Donaciinae. In: Bezděk J, Sekerka L (Eds) Catalogue of Palaearctic Coleoptera. Volume 6/2. Revised and updated Second Edition. Chrysomeloidea II (Megalopodidae, Orsodacnidae, Chrysomelidae). Brill, Leiden.
- Geiser E, Bezděk J (in press) Donaciinae. In: Bezděk J, Sekerka L (Eds) Catalogue of Palaearctic Coleoptera. Volume 6/2. Revised and updated 2nd Edn. Chrysomeloidea II (Megalopodidae, Orsodacnidae, Chrysomelidae). Brill, Leiden.
- Geiser E, Geiser R (2023) Original descriptions of Palaearctic species of the genus Plateumaris C. G. Thomson, 1859 (Coleoptera, Chrysomelidae, Donaciinae) and their translations. In: Chaboo CS, Schmitt M (Eds) Research on Chrysomelidae 9. ZooKeys 1177: 235–258. https://doi.org/10.3897/zookeys.1177.103212
- Geoffroy EL (1762) Histoire abrégée des insectes qui se trouvent aux environs de Paris, dans laquelle ces animaux sont rangés suivant un ordre méthodique. Tome première. Paris: Durand, [[4] + xxxviii +] 523 [+ [1]] pp.[, 10 pls.] [Re-issued in identical edition in 1764]. [Work rejected for nomenclatorial purposes (ICZN, Opinion 228), but some names subsequently made available (ICZN, 1994)] https://www.biodiversitylibrary.org/item/51065#page/9/mode/1up
- Goecke H (1937) Bemerkungen zu Plateumaris weisei Duviv. (Col. Chrys.). 7. Beitrag zur Kenntnis der Donaciinen. Festschrift zum 60. Geburtstage von Prof. Dr. Embrik Strand (Riga) 2: 371–372. https://dspace.lu.lv/dspace/handle/7/61203
- Goecke H (1942a) Donacia imitatrix Apfelbeck. Kleine Mitteilung 1397. Entomologische Blätter 38: 220.
- Goecke H (1942b) Plateumaris intermedia Apfelbeck. Kleine Mitteilung 1396. Entomologische Blätter 38: 220.
- Goecke H (1943) Monographie der Schilfkäfer. II. Die fossilen Funde und ihre Bestimmung. 13. Beitrag zur Kenntnis der Donaciinen. Nova Acta Leopoldina N.F. 12: 339–380.
- Goecke H (1957a) Plateumaris roscida. Kleine Mitt. 1576. Entomologische Blätter 53: 124.
- Goecke H (1957b) Plateumaris sericea. Kleine Mitt. 1577. Entomologische Blätter 53: 124.
- Goecke H (1960) Monographie der Schilfkäfer. III. Die Gattungen und Arten der Donaciinae (Col. Chrys.) und ihre Verbreitung. 18. Beitrag zur Kenntnis der Donaciinae. Entomologische Blätter 56(1): 1–19.
- Gortani M (1906) Saggio sulla distribuzione geografica die Coleotteri in Friuli. Tipografia G.B. Doretti, Udine, 93 pp.
- Gressitt JL, Kimoto S (1961) The Chrysomelidae (Coleoptera) of China and Korea, Part 1. Pacific Insects Monograph 1 A: 1–299.
- Gruev B, Tomov V (1984) Coleoptera, Chrysomelidae: Chast 1. Orsodacninae, Zeugophorinae, Donaciinae, Criocerinae, Clytrinae, Cryptocephalinae, Lamprosomatinae, Eumolpinae. Band 13 von Fauna Bulgaria - Izd-vo na Bŭlgarskata akademiia na naukite, 220 pp.
- Gus’kova EV, Efimov DA, Atuchin AA (2018) Leaf beetles (Coleoptera: Chrysomelidae) of the Kuznetsk-Salair Mountain Area (Russia, Siberia). Part One: Subfamilies Donaciinae, Criocerinae, Cassidinae and Chrysomelinae. Entomologist’s Gazette 69(2): 123–145. https://doi.org/10.31184/G00138894.692.1664
- Hayashi M (2001) Taxonomic notes on Plateumaris amurensis Weise and Plateumaris weisei (Duvivier) (Coleoptera: Chrysomelidae: Donaciinae). Entomological Review of Japan 56(1): 1–8.
- Hayashi M (2002) Host Plants of Plateumaris constricticollis babai Chûjô (Coleoptera: Chrysomelidae: Donaciinae). Entomological Review, Japan 57(2): 203–204.
- Hayashi M (2020) Revision of extant and fossil Donaciinae (Coleoptera: Chrysomelidae) of Japan. Japanese Journal of Systematic Entomology, Supplementary Series (2): 61–160.
- Hayashi M, Cho H-W (2017) Occurrence of Plateumaris weisei (Duvivier) (Coleoptera, Chrysomelidae, Donaciinae) in South Korea. Elytra, Tokyo. New Series 7(1): 233–234.
- Hayashi M, Shiyake S (2004) A check-list of the Japanese Member of Donaciinae (Coleoptera: Chrysomelidae). Entomological Review of Japan 59(1): 113–125.
- Hayashi M, Shiyake S (2011) Past Distribution of Plateumaris weisei (Duvivier) (Coleoptera, Chrysomelidae, Donaciinae) at the Last Glacial Maximum in Shiga Prefecture, West Honshu, Japan. Elytra, Tokyo, New Series 1(2): 315–320.
- Hayashi M, Sota T (2014) Quaternary Donaciine beetles (Coleoptera, Chrysomelidae) in Japan: Colonization and divergence patterns inferred from fossil and molecular data. Quaternary International 341: 255–266. https://doi.org/10.1016/j.quaint.2013.08.022
- Hayashi M, Tominaga O (2005) Records of Donaciinae from Primorsky Province in 2004, with notes on the distribution of Plateumaris shirahatai Kimoto (Coleoptera: Chrysomelidae). Entomological Review of Japan 60(1): 1–7.
- Hendrich L, Moriniére J, Haszprunar G, Hebert PD, Hausmann A, Köhler F, Balke M (2015) A comprehensive DNA barcode database for Central European beetles with a focus on Germany: Adding more than 3500 identified species to BOLD. Molecular Ecology Resources 15(4): 795–818. https://doi.org/10.1111/1755-0998.12354
- Herbst JFW (1784) Kritisches Verzeichniss meiner Insectensammlung. Fortsetzung. Curculio-Dytiscus. Archiv der Insectengeschichte, herausgegeben von J. C. Fuessly 5(1): 69–128[, pls 24–28]. http://www.animalbase.uni-goettingen.de/zooweb/servlet/AnimalBase/home/reference?id=2667
- Hoppe DH (1795) Enumeratio insectorum elytratorum circa Erlangam indigenarum observationibus iconibusqve illustrata. Dissertatio inauguralis quam summi numinis auspicio ex decreto gratiosae facultatis medicae pro summis in medicina honoribus legitime obtinendis publico eruditorum examini submittit David Henricus Hoppe. Hilpertianis, Erlangae, 70 pp[, 1 pl. DP: [30] April 1795 (Bousquet 2016)].
- Iablokoff-Khnzorian SM (1966) Considérations sur l’édéage des chrysomelides et son importance phylogénique. Entomologiste 22: 115–137. http://lentomologiste.fr/tome-22-1966/
- Jacobsohn [= Jacobson] GG (1892) Analytische Übersicht der bekannten Donacia- und Plateumaris-Arten der Alten Welt. Horae Societatis Entomologicae Rossicae 26: 412–437. https://www.biodiversitylibrary.org/item/46110#page/976/mode/1up
- Jacobsohn [= Jacobson] GG (1894) Adnotationes de Chrysomelidis nonnullis novis vel parum cognitis. Horae Societatis Entomologicae Rossicae 28: 242–246. https://www.biodiversitylibrary.org/item/81150#page/892/mode/1up
- Jacoby M (1885) Descriptions of the phytophagous Coleoptera of Japan, obtained by Mr. George Lewis during his second journey, from February 1880 – September 1881. Part I. Proceedings of the scientific meetings of the Zoological society of London 1885: 190–211[, pl. XI]. https://doi.org/10.1111/j.1096-3642.1885.tb02894.x
- Jolivet P (1970) Coleopterorum Catalogus supplementa. Pars. 51 fasc. 2 (ed. secunda), Donaciinae. In: Steel WO (Ed.) Coleopterorum Catalogus, Supplementa; W. Junk, The Hague 51(2), 1–71.
- Jolivet P, Petitpierre E, Hsiao TH (1988) Biology of Chrysomelidae. Series Entomologica Vol. 42. Kluwer Academic Publishers, Dordrecht-Boston-London, 615 pp. https://doi.org/10.1007/978-94-009-3105-3
- Kaszab Z (1962) Levélbogarak, Chrysomelidae. Magyarorzág Állatvilága, Fauna Hungariae 63. Akadémiai Kiadó, Budapest, 416 pp.
- Kimoto S (1963) Descriptions of new species of Chrysomelidae from Japan, with notes on some known species. Fragmenta Coleopterologica 3–4: 13–18.
- Kimoto S (1971) [new taxa]. In: Kimoto S, Hiura I (1971) List of the Chrysomelidae specimen preserved in the Osaka Museum of Natural History, III. (Insecta: Coleoptera). Bulletin of the Osaka Museum of Natural History 25: 1–26. https://doi.org/10.5962/bhl.part.19678
- Kimoto S (1981) New or little known Japanese Donaciinae (Coleoptera: Chrysomelidae). Bulletin of the Osaka Museum of Natural History 34: 23–26.
- Kippenberg H (1994) Chrysomelidae (without Alticinae). In: Lohse GA, Lucht W (Eds) Lohse-Lucht: Die Käfer Mitteleuropas (3rd Supplement), Volume 14: 17–92.
- Kirby W (1837) Part the fourth and last. The insects. In: Richardson J (Ed.) Fauna Boreali-Americana; or the zoology of the northern parts of British America: containing descriptions of the objects of natural history collected on the late Northern Land Expedition, under command of captain Sir John Franklin, R. N. J. Fletcher, Norwich, [xxxix +] 325 [+ 1] pp[, 8 pls]. https://www.biodiversitylibrary.org/item/129782#page/11/mode/1up
- Kleinschmidt B, Kölsch G (2011) Adopting Bacteria in Order to Adapt to Water—How Reed Beetles Colonized the Wetlands (Coleoptera, Chrysomelidae, Donaciinae). Insects 2(4): 540–554. https://doi.org/10.3390/insects2040540
- Kolossow J (1929) Was ist Donacia asiatica Faldermann? Entomologische Blätter 25: 189–190.
- Kolossow J (1930) Annotationes de quibusdam Donaciini. Coleopterologisches Zentralbl. Bd. 5. Organ für systematische Coleropterologie der paläarktischen Zone. Herausgegeben von Hans Wagner 5(1): 28–29.
- Kölsch G, Pedersen BV (2008) Molecular phylogeny of reed beetles (Col., Chrysomelidae, Donaciinae): The signature of ecological specialization and geographical isolation. Molecular Phylogenetics and Evolution 48(3): 936–952. https://doi.org/10.1016/j.ympev.2008.05.035
- Krause R (1982) Zur Faunistik, Ökologie und Phänologie der Blattkäfer der Sächsischen Schweiz (Insecta, Coleoptera, Chrysomelidae). Faunistische Abhandlungen 9: 1–55.
- Kunze G (1818) Beiträge zur Monographie der Rohrkäfer. Neue Schriften der Naturforschenden Gesellschaft zu Halle 2(4): 1–56.
- Latreille PA (1796) Précis des caractères génériques des Insectes, disposés dans un ordre naturel I–XIII, Paris et Brive, 201 pp. https://www.biodiversitylibrary.org/item/119655#page/7/mode/1up
- Latreille PA (1802) Histoire naturelle, générale et particulière, des crustacés et des insectes. Ouvrage faisant suite à l’histoire naturelle générale et particulière, composée par Leclerc de Buffon, et rédigée par C. S. Sonnini, membre de plusieurs sociétés savantes. Tome troisième. Familles naturelles des genres. Paris: F. Dufart, [xii + 13–]467 pp. [+ 1 p. errata] https://www.biodiversitylibrary.org/item/80064#page/5/mode/1up
- Lays P (2002) Notes on the Donaciines (Coleoptera Chrysomelidae Donaciinae). Bulletin de la Société Royale Belge d’Entomologie 138: 143–150.
- LeGrand G (1861a) Liste des coléoptères du département de l’Aube. Mémoires de la Société Académique d’Agriculture, des Sciences, Arts et Belles-Lettres du Département de l’Aube, Deuxième Série 12: 181–274. [DP: 13 July 1861 (Bibl. France 1861, 50(28): 338).]
- LeGrand G (1861b) Liste des coléoptères du département de l’Aube. Mémoires de la Société Académique d’Agriculture, des Sciences. Arts et Belles-Lettres du Département de l’Aube 12(2): 1–98.
- Linnaeus C (1758) Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymies, locis. Tomus I. Editio decima, reformata. Impensis Direct. Laurentii Salvii, Holmiae, [iv +] 824 [+ 1] pp. https://www.biodiversitylibrary.org/item/10277#page/419/mode/1up
- Linnaeus C (1760) Fauna suecica sistens animalia Sueciae Regni: Mammalia, Aves, Amphibia, Pisces, Insecta, Vermes. Distributa per classes et ordines, genera et species. 2nd ed. Laur. Salvii, Stockholmiae, 45 + 578 pp. https://www.biodiversitylibrary.org/item/100333#page/254/mode/1up
- Löbl I, Smetana A [Eds] (2010) Catalogue of Palaearctic Coleoptera. Vol. 6. Apollo Books, Stenstrup, 924 pp. https://doi.org/10.1163/9789004260917_004
- Löbl I, Smetana A [Eds] (2013) Catalogue of Palaearctic Coleoptera, vol. 8. Brill, Leiden, Boston, 700 pp. https://doi.org/10.1163/9789004259164_010
- Lopatin IK (1984) Leaf Beetles (Chrysomelidae) of Central Asia and Kazakhstan. Amerind Publ. Co. Pvt. Ltd., New Delhi, [xxxiv +] 416 pp. [Translation from Zhuki-Listoedy (Chrysomelidae) Srednei Azii i Kazakhstana. Nauka Publ., Leningrad]
- Lopatin IK, Kulenova KZ (1986) Zhuky-Listoedy (Coleoptera, Chrysomelidae) Kazakhstana: Opredelitel [Leaf Beetles (Coleoptera, Chrysomelidae) of Kazakhstan: Identification Key]. Izdatel’stvo “Nauka” Kazakhskoi SSR, Alma-Ata, 199 pp. [In Russian]
- Mann JS, Crowson RA (1983) Observations on the internal anatomy and classification of Donaciinae (Col. Chrysomelidae). Entomologist’s Monthly Magazine 119: 17–27.
- Marx EJF (1957) A review of the subgenus Donacia in the Western hemisphere (Coleoptera, Donaciidae). Bulletin of the American Museum of Natural History, New York 112(3): 191–278[, figs 1–54]. https://bugguide.net/node/view/1121849
- Medvedev LN (1973) Novye zhuki-listoedy (Coleoptera, Chrysomelidae) palearktiki. Entomologicheskoe Obozrenie 52: 876–885. [New leaf beetles (Coleoptera, Chrysomelidae) from Palaearctic]
- Medvedev LN (1978) Taksonomicheskie zametki o zhukakh-listoedakh (Coleoptera, Chrysomelidae, Sakhalina i Kuril’skih ostrovov. Trudy Biologo-Pochvennogo Instituta (N.S. ) 50(153): 82–86. [In Russian] [= Taxonomic notes on leaf beetles (Coleoptera, Chrysomelidae) of Sakhalin and the Kuril Islands. – New data on insects of Sakhalin and the Kuril Islands. Proceedings of the Biological and Soil Institute New series 50(153): 82–86.]
- Medvedev LN (1992) Fam. Chrysomelidae. In: Opredelitel’ nasekomyh Dalnego Vostoka SSSR. V3. Part 2. Nauka, St. Petersburg, 533–602. [In Russian]
- Medvedev LN, Muravitsky OS (2014) [Медведев ЛН, Муравицкий ОС (2014) Описания неизвестных ранее личинок листоедов (Coleoptera, Chrysomelidae) из Европейской части России и с Украины // Энтомологическое обозрение.] Opisanya neizvestnikh ranee lichinok listoedov (Coleoptera, Chrysomelidae) iz Evropeiskoy chasti Rossii i s Ukrainoi. Entomologicheskoe obozrenie 93(1): 131–144. [In Russian] [= Descriptions of previously unknown leaf beetle larvae (Coleoptera, Chrysomelidae) from the European part of Russia and Ukraine. Entomological Review 93(1): 131–144.]
- Menzies IS, Cox ML (1996) Notes on the natural history, distribution and identification of British reed beetles. British Journal of Entomology and Natural History (Supplement 9): 137–162. https://www.biodiversitylibrary.org/page/29883043#page/419/mode/1up
- Mohr KH (1966a) 88. Familie: Chrysomelidae: 95–280. In: Freude H, Harde KW, Lohse GA (Eds) Die Käfer Mitteleuropas. Vol. 9. Goecke & Evers, Krefeld, 299 pp.
- Mohr KH (1966b) Ergebnisse der Albanien-Expedition 1961 des Deutschen Entomologischen Institutes, 47. Beitrag, Coleoptera: Chrysomelidae II. Beiträge zur Entomologie 16: 347–380. https://www.zobodat.at/publikation_articles.php?id=341187
- Monrós F (1959) Los Generos de Chrysomelidae (Coleoptera). Opera Lilloana III: 5–337[, 3 pls].
- Nakane T (1963) New or little-known Coleoptera from Japan and its adjacent regions. XVI. Fragmenta Coleopterologica 4/5: 18–20.
- Nakane T (1999) Notes on the specific name of “Plateumaris sericea” in Japan (Coleoptera, Chrysomelidae). Nature and Insects (Tokyo) 34(8): 45–46.
- Narita Y (2003) Descriptions of Donaciine larvae (Coleoptera, Chrysomelidae) from Japan. Elytra, Tokyo 31(1): 1–30. http://coleoptera.sakura.ne.jp/Elytra/31(1)001NaritaY
- Olivier AG (1791) Encyclopédie métodique, ou par ordre de matières; par une société de gens de lettres, de savans et d’artistes; précédée d’un vocabulaire universel, servant de table pour tout l’ouvrage, ornée des portraits de Mm. Diderot et d’Alembert, premiers éditeurs de l’Encyclopédie. Histoire Naturelle. Insectes. Tome sixième. Pars I. Panckoucke, Paris, 704 pp. https://www.biodiversitylibrary.org/item/156051#page/302/mode/1up
- Olivier AG (1800) [second part, first part started in 1795]: Entomologie, ou histoire naturelle des insectes. Avec leur caractéres génériques et spécifiques, leur description, leur synonymie, et leur figure enluminée. Coléoptères. Tome quatrième. de Lanneau, Paris, 519 pp. [+72 pls.] [note: each genus is separately paginated] https://www.biodiversitylibrary.org/item/125991#page/401/mode/1up
- Pallas PS (1773) Reise durch verschiedene Provinzen des Russischen Reichs. Zweyter Theil. Zweytes Buch vom Jahr 1771. St. Petersburg: Kayserliche Akademie der Wissenschaften, 371–744. http://resolver.sub.uni-goettingen.de/purl?PPN330045997
- Panzer GWF (1795) Entomologia Germanica exhibens insecta per Germaniam indigena secundum classes, ordines, genera, species adiectis synonymis, locis, observationibus. I. Eleuterata. Norimbergae: Felssecker, [8] + 12 + 370 + 2 pp. [12 pls.] [DP: 14 February 1795 (Bousquet 2016).] https://www.biodiversitylibrary.org/item/44240#page/282/mode/1up
- Paykull G (1799) Fauna Svecica; Insecta. (Coleoptera.). Tomus II. Joh. F. Edman, Upsaliae, 234 pp.
- Perris E (1864) Description de quelques espèces nouvelles de coléoptères et notes diverses. Annales de la Société Entomologique de France 4(4): 275–310. https://www.biodiversitylibrary.org/page/8254869#page/281/mode/1up
- Reis F, Kirsch R, Pauchet Y, Bauer E, Bilz LC, Fukumori K, Fukatsu T, Kölsch G, Kalthneoth M (2020) Bacterial symbionts support larval sap feeding and adult folivory in (semi-)aquatic reed beetles. Nature Communications 11(1): 2964. https://doi.org/10.1038/s41467-020-16687-7
- Reitter E (1920) Bestimmungstabellen der europäischen Coleopteren. Heft 88: Chrysomelidae, 1.Teil: Tribus Donaciinae. Wiener Entomologische Zeitung 38(1–3): 21–43. https://www.biodiversitylibrary.org/page/12110882#page/279/mode/1up
- Rheinheimer J, Hassler M (2018) Die Blattkäfer Baden-Württembergs. Kleinsteuber Books, Karlsruhe, 928 pp.
- Samin N (2018) Nineteen new records of species of the family Chrysomelidae (Coleoptera: Chrysomeloidea) for the fauna of Iran. Acta Zoologica Bulgarica 70(4): 459–464.
- Schenkling S (1917) Erklärung der wissenschaftlichen Käfernamen aus Reitter’s Fauna Germanica. K.G. Lutz Verlag, Stuttgart, 79 pp. https://www.biodiversitylibrary.org/item/37302#page/6/mode/1up
- Schilling PS (1838) [new taxon]. In: Schummel TE (Ed.) Bericht über die Arbeiten der entomologischen Section im Jahre 1837. Uebersicht der Arbeiten und Veränderungen der Schlesischen Gesellschaft für Vaterländische Kultur (Breslau) [1837], 99–112. [31 December 1838] [Volume title page]
- Schilsky J (1908) Neue märkische Käfer und Varietäten aus der Gegend von Luckenwalde. Deutsche Entomologische Zeitschrift 52: 599–604. https://www.biodiversitylibrary.org/item/105322#page/651/mode/1up
- Schrank de Paula F (1781) Enumeratio insectorum Austriae indigenorum. E. Klett et Franck, Augustae Vindelicorum, [24 +] 548 [+ 4] pp. https://www.zobodat.at/publikation_volumes.php?id=34990
- Scopoli JA (1772) Observationes zoologicae. Annus V. Historico-Naturalis. Christ. Gottlob Hilscheri, Lipsiae, 70–128. https://www.biodiversitylibrary.org/item/137649#page/74/mode/1up
- Semenov AP (1895) Coleoptera asiatica nova. V. Horae Societatis Entomologicae Rossicae 29[1894–1895]: 251–270. https://www.biodiversitylibrary.org/item/45835#page/311/mode/1up
- Shavrov WB (1948) Novye formy i mestonakhozhdeniya donaciy (Coleoptera, Chrysomelidae, Subfam. Donaciinae) fauny SSSR. II. Byulleten’ Moskovskogo Obshchestva Ispytateley Prirody, Otdel biologicheskiy (N. S. ) 53(1): 49–52.
- Silfverberg H (2010) Donaciinae. In Löbl I, Smetana A (Eds) Catalogue of Palaearctic Coleoptera, vol. 6. Apollo Books, Stenstrup, 354–368.
- Solsky SM (1871) Coleopteres de la Siberie orientale. Horae Societatis Entomologicae Rossiae 8[1871–1872]: 232–277. https://www.biodiversitylibrary.org/item/185648#page/278/mode/1up
- Sota T, Hayashi M (2007) Comparative historical biogeography of Plateumaris leaf beetles (Coleoptera: Chrysomelidae) in Japan: interplay between fossil and molecular data. Journal of Biogeography 34(6): 977–993. https://doi.org/10.1111/j.1365-2699.2006.01672.x
- Sota T, Hayashi M, Yagi T (2007) Geographic variation in body and ovipositor sizes in the leaf beetle Plateumaris constricticollis (Coleoptera: Chrysomelidae) and its association with climatic conditions and host plants. European Journal of Entomology 104(2): 165–172. https://doi.org/10.14411/eje.2007.025
- Sota T, Bocak L, Hayashi M (2008) Molecular phylogeny and historical biogeography of the Holarctic wetland leaf beetle of the genus Plateumaris. Molecular Phylogenetics and Evolution 46(1): 183–192. https://doi.org/10.1016/j.ympev.2007.10.016
- Standfuss K, Kerzhner IM (2004) Publikationsdaten der “Trudy Russkogo Entomologicheskogo Obshchestva” und “Horae Societatis Entomologicae Rossicae”, 1861–1932. Entomofauna 25: 237–248. https://www.zobodat.at/publikation_articles.php?id=42970
- Steinhausen W (1994) Chrysomeloidea. In: Klausnitzer B (Ed.) Die Larven der Käfer Mitteleuropas. 2. Band Myxophaga, Phytophaga Teil 1. Goecke und Evers, Krefeld.
- Suffrian E (1846) Bemerkungen zu den in “Lacordaire’s Monographie des coléoptères subpentamères de la famille des Phytophages” vorkommenden deutschen Arten. Entomologische Zeitung (Stettin) 7: 80–92, 152–160. https://www.biodiversitylibrary.org/page/36267045#page/520/mode/1up
- Thomson CG (1859) Skandinaviens Coleoptera, synoptiskt bearbetade. Tom. I. Lundbergska Boktryckeriet, Lund, [2 +] 290 pp. https://www.biodiversitylibrary.org/item/235561#page/168/mode/1up
- Thomson CG (1866) Skandinaviens Coleoptera, synoptiskt bearbetade. Tom. VIII. Lundbergska Boktryckeriet, Lund, 409 [+ lxxv] pp. https://www.biodiversitylibrary.org/item/235559#page/7/mode/1up
- Tominaga O, Katsura K (1984) Studies of the Japanese Donaciinae (Coleoptera: Chrysomelidae). 2. Notes on geographical diversity of Plateumaris contricticollis, with description of an allied new species. Bulletin of the Osaka Museum of Natural History 37: 25–40. https://archive.org/details/bulletin-osaka-museum-natural-history-37-025-027
- Walter T (1990) Naturmonographie Ruggeller Riet. Naturkundliche Forschung im Fürstentum Liechtenstein 12, 40 pp.
- Warchałowski A (2003) Chrysomelidae. The leaf-beetles of Europe and the Mediterranean area. Natura Optima Dux Foundation, Warszawa, 600 pp.
- Warchałowski A (2010) The Palaearctic Chrysomelidae – Identification Keys. Natura Optima Dux Foundation, Warszawa, 1212 pp.
- Weise J (1893) Chrysomelidae. Lieferung 6. In: Naturgeschichte der Insecten Deutschland. Erste Abtheilung Coleoptera. Sechster Band [1893]. Nicolaische Verlags-Buchhandlung R. Stricker, Berlin, [xiv +] 1161 pp.[, 1 pl.] https://www.biodiversitylibrary.org/item/105313#page/69/mode/1up
- Weise J (1898) Ueber neue und bekannte Chrysomeliden. Archiv für Naturgeschichte 64(1): 177–224. https://www.zobodat.at/publikation_articles.php?id=231328
- Weise J (1900) Beschreibungen von Chrysomeliden und synonymische Bemerkungen. Archiv für Naturgeschichte 66(1): 267–296. https://www.zobodat.at/publikation_articles.php?id=231929
- Weise J (1912) Beitrag zur Kenntnis der Chrysomeliden. Archiv für Naturgeschichte 78(A2): 76–98. https://www.zobodat.at/publikation_articles.php?id=235190
- Westhoff F (1882) Die Käfer Westfalens. Verhandlungen des Naturhistorisches Vereins der Preussischen Reinlande und Westfalens und des Reg.-Bezirks Osnabrück (Bonn) 38 [1881–1882], Supplement: [i–xxviii] + 141–323 + [1] pp.
- Winkler A (1930) Catalogus Coleopterorum regionis palaearcticae, pars 11: 1265–1392. [Wien.]
- Zaitzev FA (1930) K rasprostraneniyu na Kavkaze vidov triby Donaciini (Coleoptera, Chrysomelidae). Bulletin du Musée de GeoPedriliargie 5: 105–114.
- Zschach JJ (1788) Museum N. G. Leskeanum. Pars entomologica ad systema entomologiae Cl. Fabricii ordinata. Lipsiae: Müller, 136 pp[, 3 pls.] https://www.biodiversitylibrary.org/item/256042#page/33/mode/1up