- ContentsContents
- Article InfoArticle Info
- CiteCite
- MetricsMetrics
- CommentComment
- RelatedRelated
- FigsFigs
- TabsTabs
- MapMap
- TaxaTaxa
- RefsRefs
- CitedCited
- NanopubsNanopubs
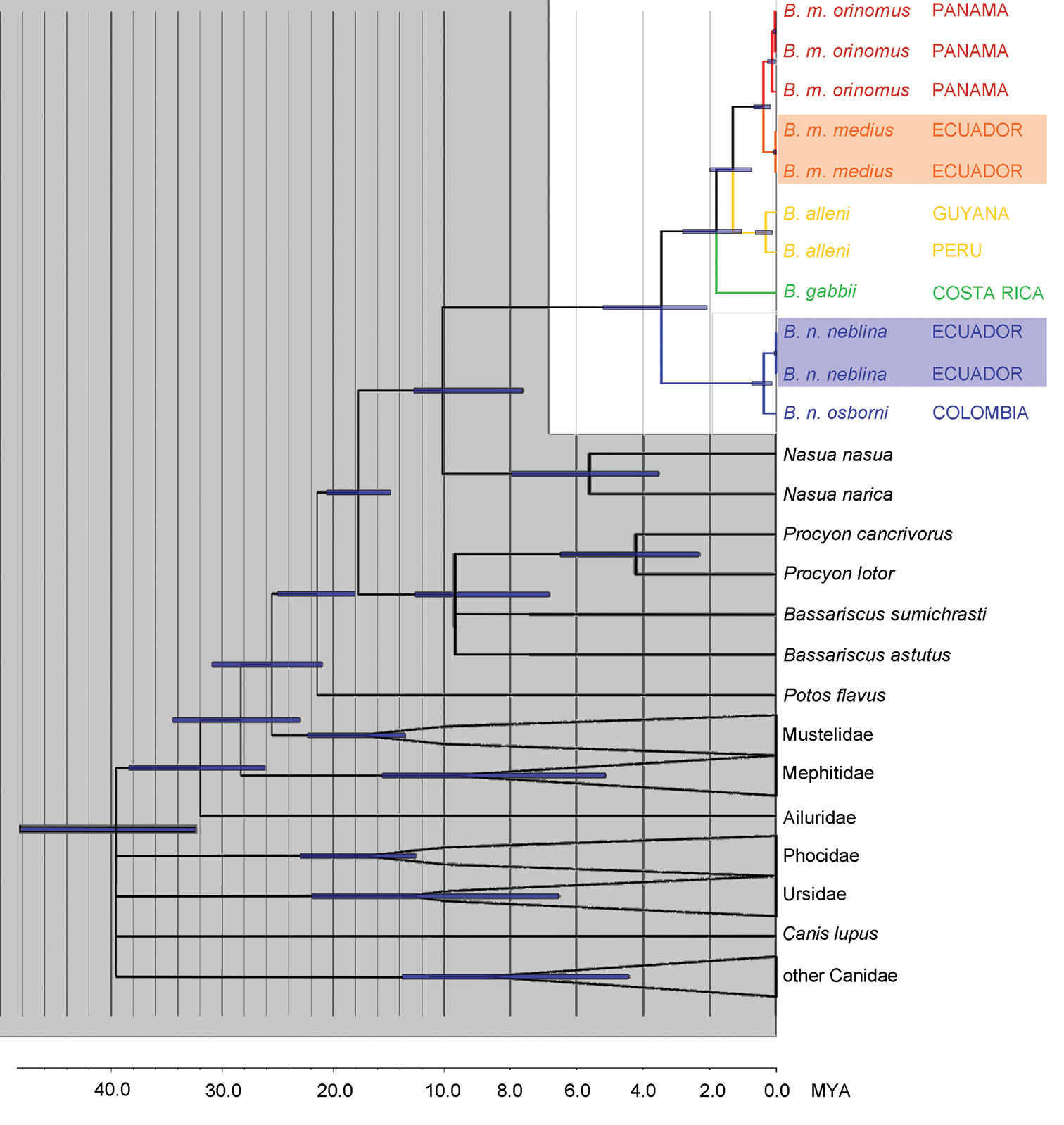
Phylogeny of the genus Bassaricyon. Phylogeny generated from the concatenated CHRNA1 and cytochrome b sequences. All analyses consistently recovered the same relationships with high support. Divergence dating was generated in BEAST; bars show the 95% confidence interval at each node. Branches without support are collapsed and outgroup clades have been collapsed, leaving monophyletic groupings with 100% support. Data for CHRNA1 are missing for Bassaricyon gabbii, for which DNA was extracted from a museum skull. All nodes in Bassaricyon have 1.00 Bayesian posterior probability, except the split between Bassaricyon gabbii and Bassaricyon alleni/Bassaricyon medius (0.97 Bayesian posterior probability). Non-focal and outgroup taxa are shaded in gray, Bassaricyon species and subspecies are color coded, samples of Bassaricyon medius medius and Bassaricyon neblina neblina that were collected within 5 km of each other in Ecuador are shaded.
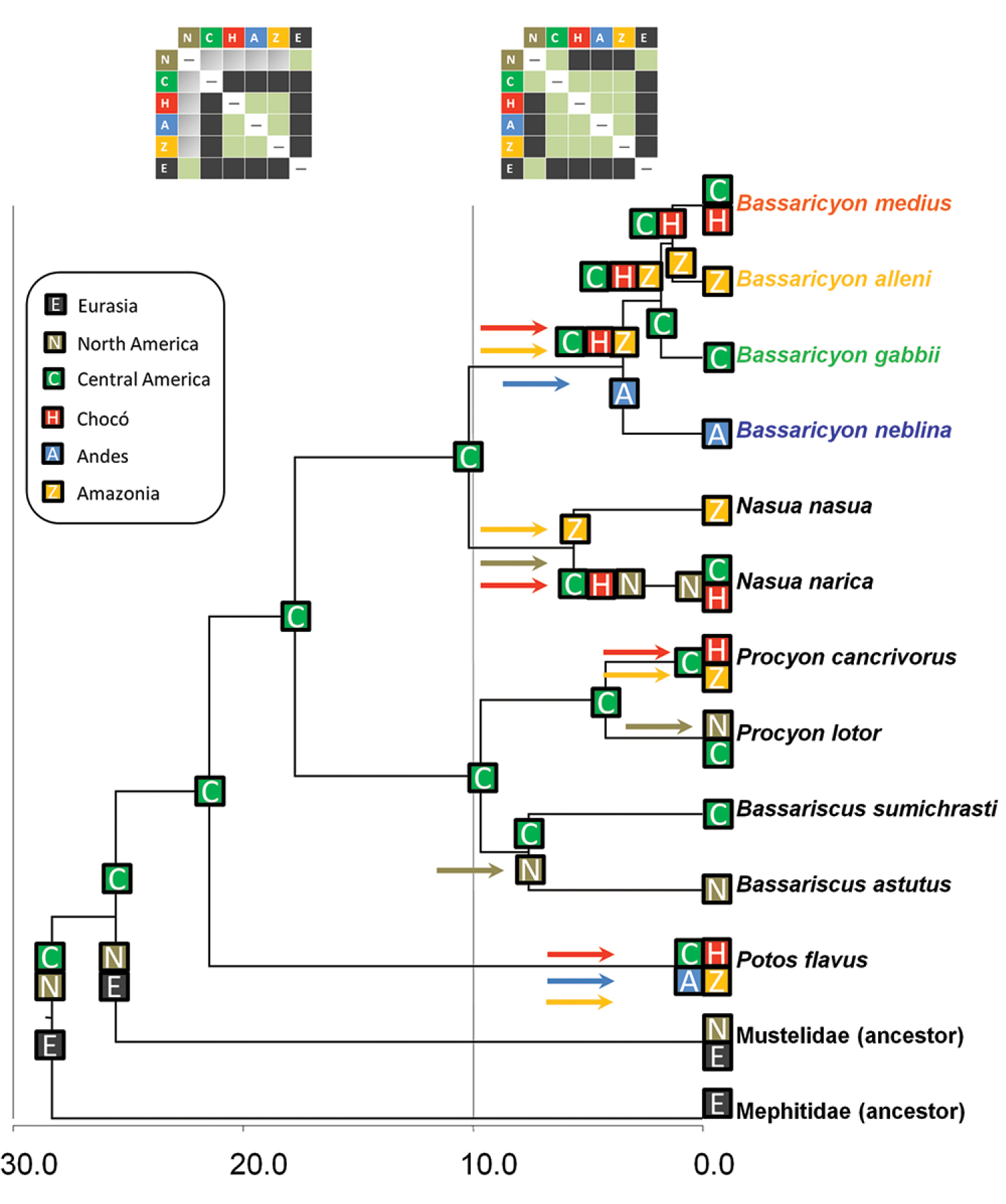
Historical biogeography of procyonids. Reconstructed under the DEC model implemented in Lagrange. See legend for geographical areas used in the analysis. Colored squares at the tip of the branches reflect the distribution of taxa, and previously inferred distributions of the ancestors of mustelids and mephitids. Colored squares at the nodes represent the geographic ranges with the highest probabilites in the DEC model inherited by each descendant branch. Colored arrows reflect dispersal events between ancestral and derived areas, with colors matching with recipient areas. Upper boxes: different dispersal constraints at time intervals 0–10 mya and 10–30 mya, the former to simulate the effect of the land bridge formation between Central and South America, the latter restricted dispersal due to the absence of the land bridge; the cells in green indicate no restriction to dispersal, cells in gray indicate a reduction by half in dispersal capability, and cells in black do not allow dispersal. Timescale in millions of years before present (mya).

Illustrations of the species of Bassaricyon. From top to bottom, Bassaricyon neblina sp. n. (Bassaricyon neblina ruber subsp. n. of the western slopes of the Western Andes of Colombia), Bassaricyon medius (Bassaricyon medius orinomus of eastern Panama), Bassaricyon alleni (Peru), and Bassaricyon gabbii (Costa Rica, showing relative tail length longer than average). Artwork by Nancy Halliday.
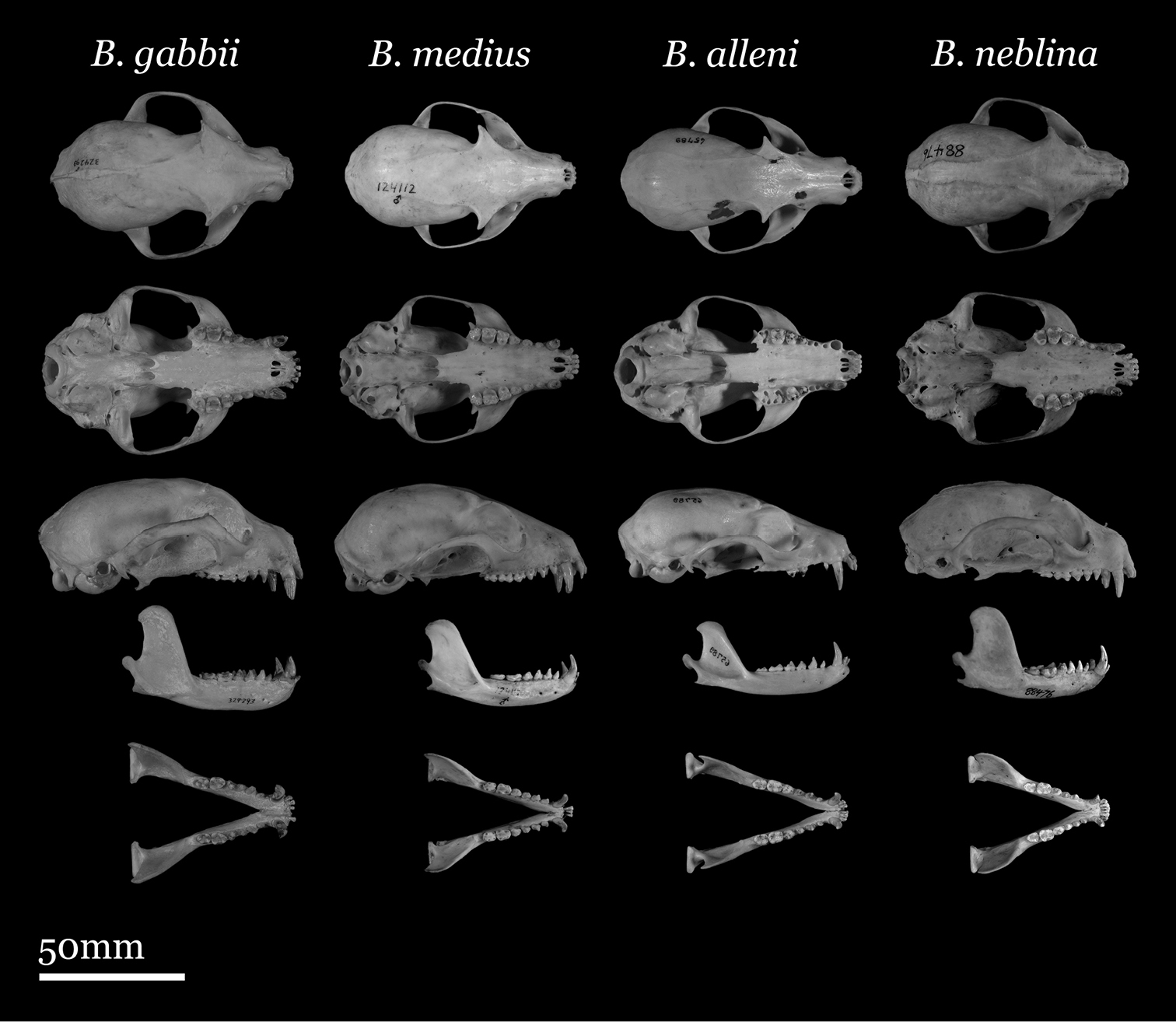
Skulls of adult male Bassaricyon. From left to right: Bassaricyon gabbii (USNM 324293, Cerro Punta, 1700 m, Chiriqui Mountains, Panama); Bassaricyon medius medius (MVZ 124112, Dagua, 1800 m, Colombia); Bassaricyon alleni (FMNH 65789, Chanchamayo, 1200 m, Junin, Peru); Bassaricyon neblina osborni (FMNH 88476, Munchique, 2000 m, Cauca, Colombia). Scale bar = 50 mm.
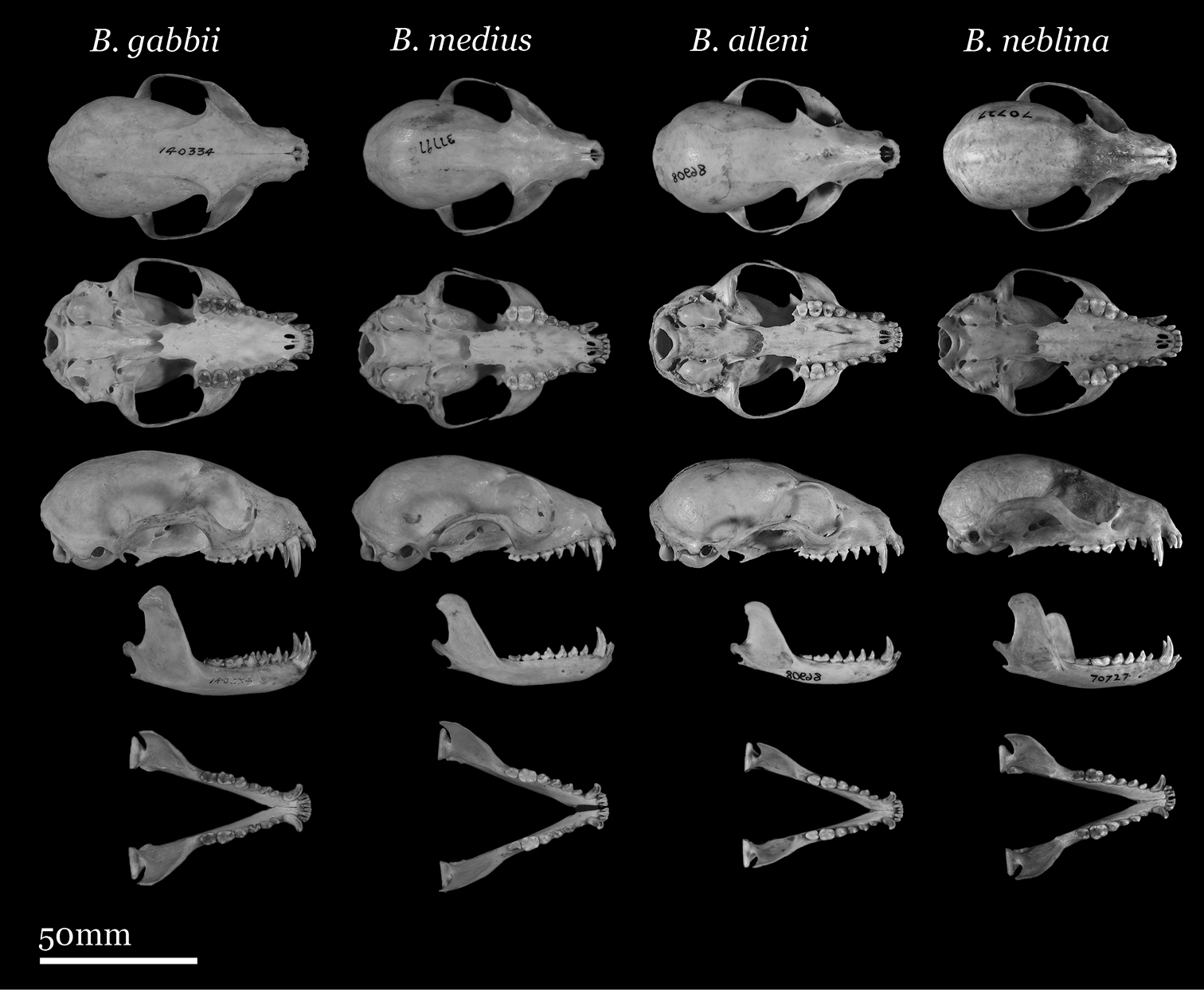
Skulls of adult female Bassaricyon. From left to right: Bassaricyon gabbii (AMNH 140334, Lajas Villa, Costa Rica); Bassaricyon medius orinomus (AMNH 37797, Puerta Valdivia, Antioquia District, Colombia); Bassaricyon alleni (FMNH 86908, Santa Rita, Rio Nanay, Maynas, Loreto Region, Peru); Bassaricyon neblina hershkovitzi (FMNH 70727, San Antonio, Agustin, Huila District, Colombia). Scale bar = 50 mm.
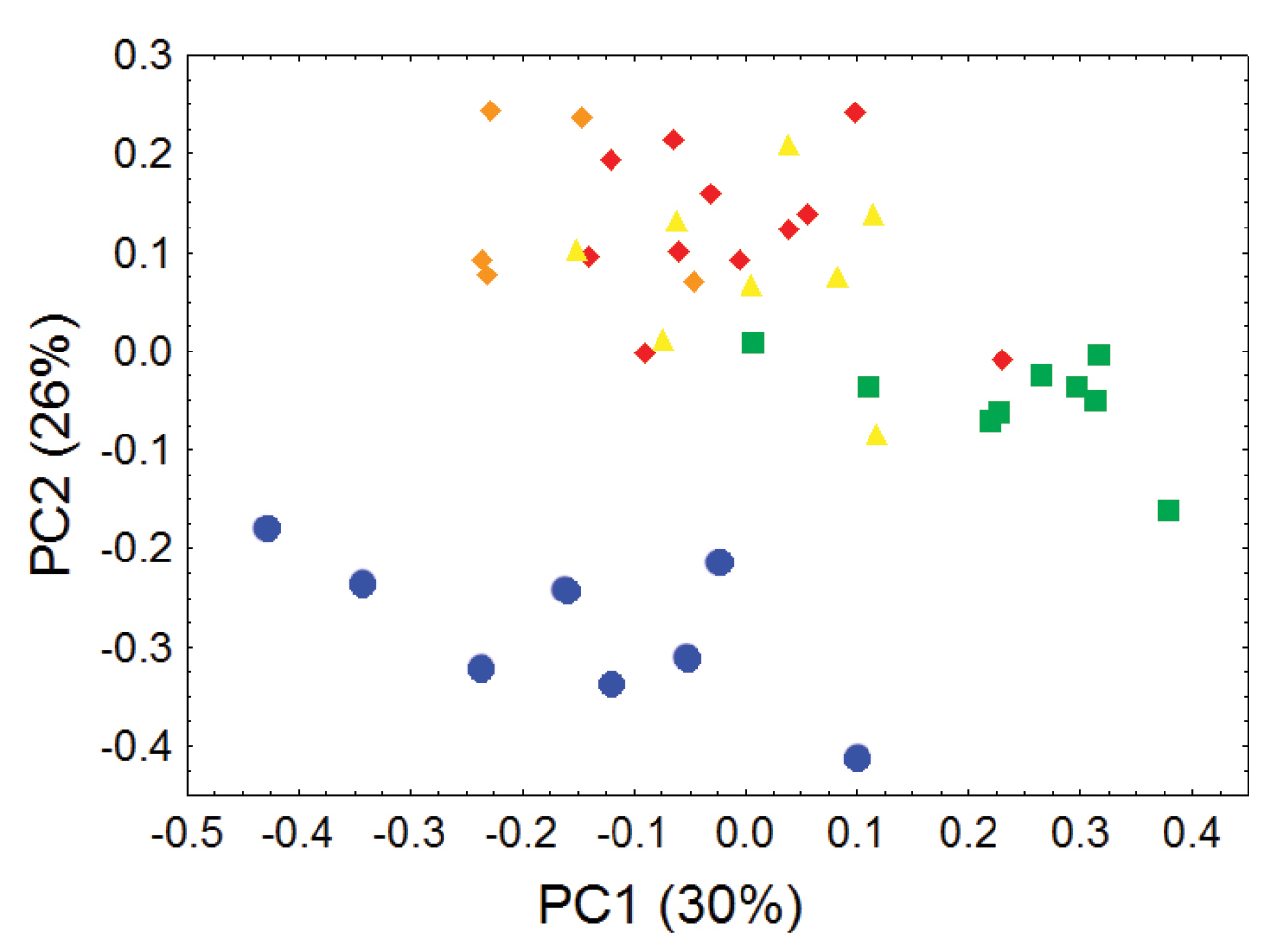
Morphometric distinction between Olinguitos and other Bassaricyon, males. Morphometric dispersion (first two components of a principal component analysis) of 41 adult male Bassaricyon skulls based on 21 craniodental measurements (see Appendix 1, Table A1). The most notable morphometric distinction is between the Olinguito (blue circles) and all other Bassaricyon taxa. The plot also demonstrates substantial morphometric variability across geographic populations of the Olinguito, which we characterize with the description of four subspecies across different Andean regions. Symbols: blue circles (Bassaricyon neblina), green squares (Bassaricyon gabbii), yellow triangles (Bassaricyon alleni), orange diamonds (Bassaricyon medius medius), red diamonds (Bassaricyon medius orinomus).
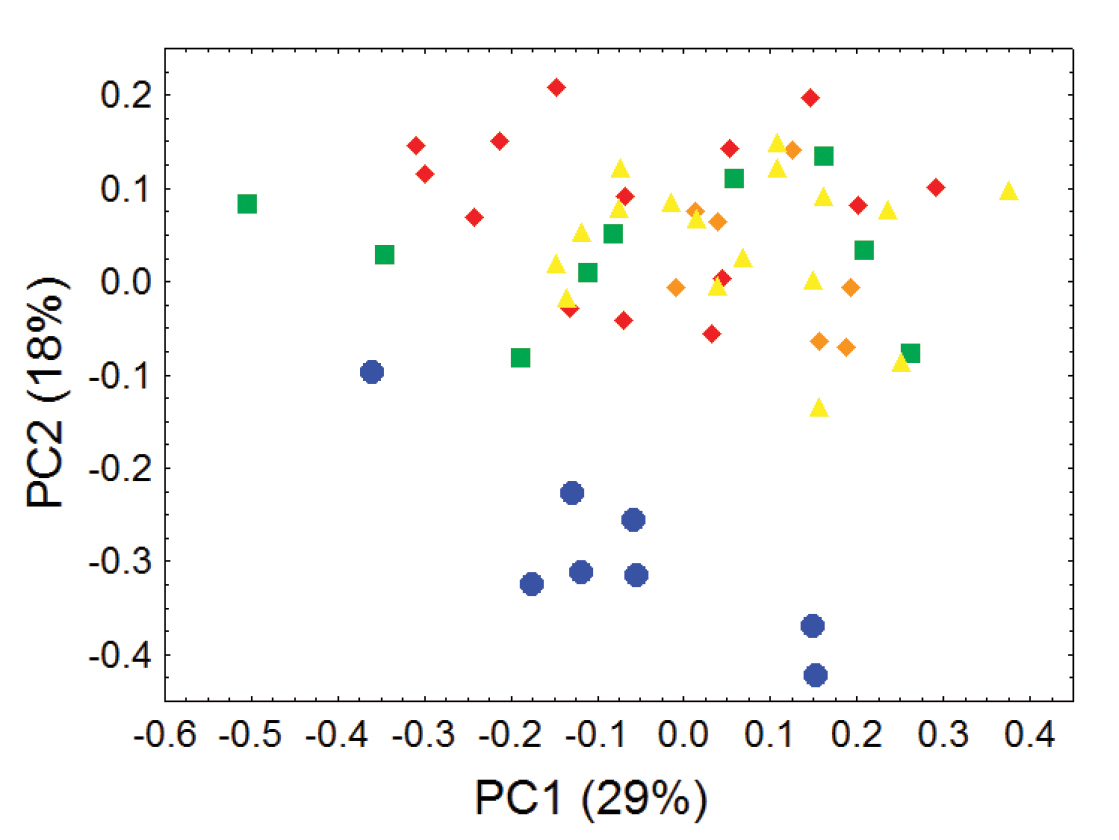
Morphometric distinction between Olinguitos and other Bassaricyon, females. Morphometric dispersion (first two components of a principal component analysis) of 55 adult female Bassaricyon skulls based on 24 craniodental measurements (see Appendix 1, Table A2). The most notable morphometric distinction is between the Olinguito (blue circles) and all other Bassaricyon taxa. The plot also demonstrates substantial morphometric variability across geographic populations of the Olinguito, which we characterize with the description of four subspecies across different Andean regions. Symbols: blue circles (Bassaricyon neblina), green squares (Bassaricyon gabbii), yellow triangles (Bassaricyon alleni), orange diamonds (Bassaricyon medius medius), red diamonds (Bassaricyon medius orinomus).
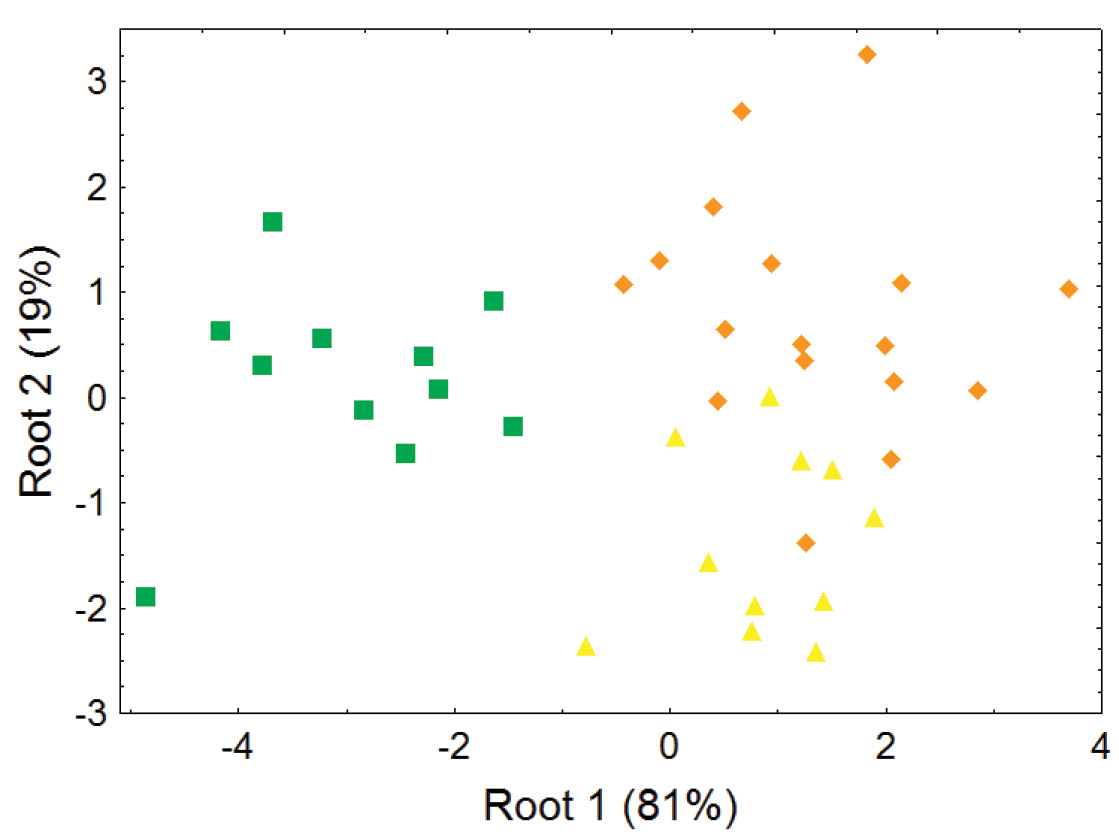
Morphometric distinction between species of Bassaricyon, excluding the Olinguito, adult males. Morphometric dispersion (first two variates of a discriminant function analysis) of 39 adult male Bassaricyon skulls based on 8 craniodental measurements (see Appendix 1, Table A3). Symbols: green squares (Bassaricyon gabbii), yellow triangles (Bassaricyon alleni), orange diamonds (Bassaricyon medius).
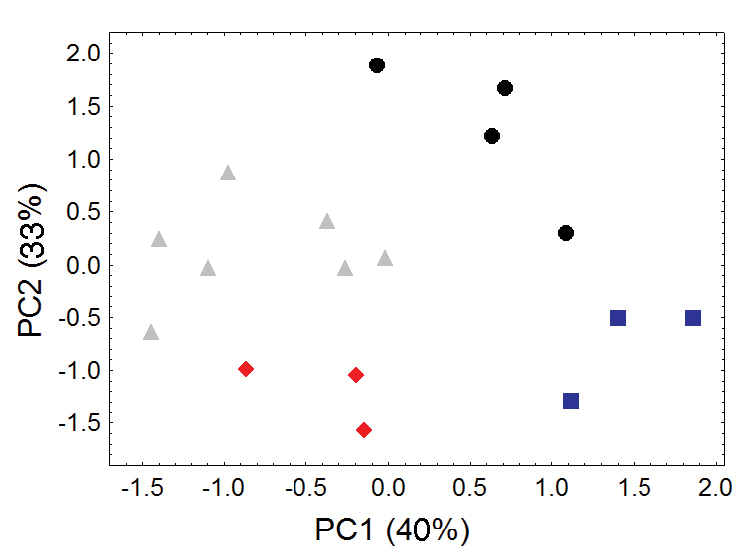
Morphometric distinction between Olinguito subspecies. Both sexes combined. Morphometric dispersion (first two components of a principal component analysis) of 17 adultskulls based on 13 cranial measurements (see Appendix 1, Table A4). (Dental measurements also discretely partition these subspecies in a separate principal component analysis, not shown.) Black dots = Bassaricyon neblina neblina; gray triangles = Bassaricyon neblina osborni; red diamonds = Bassaricyon neblina ruber; blue squares = Bassaricyon neblina hershkovitzi.
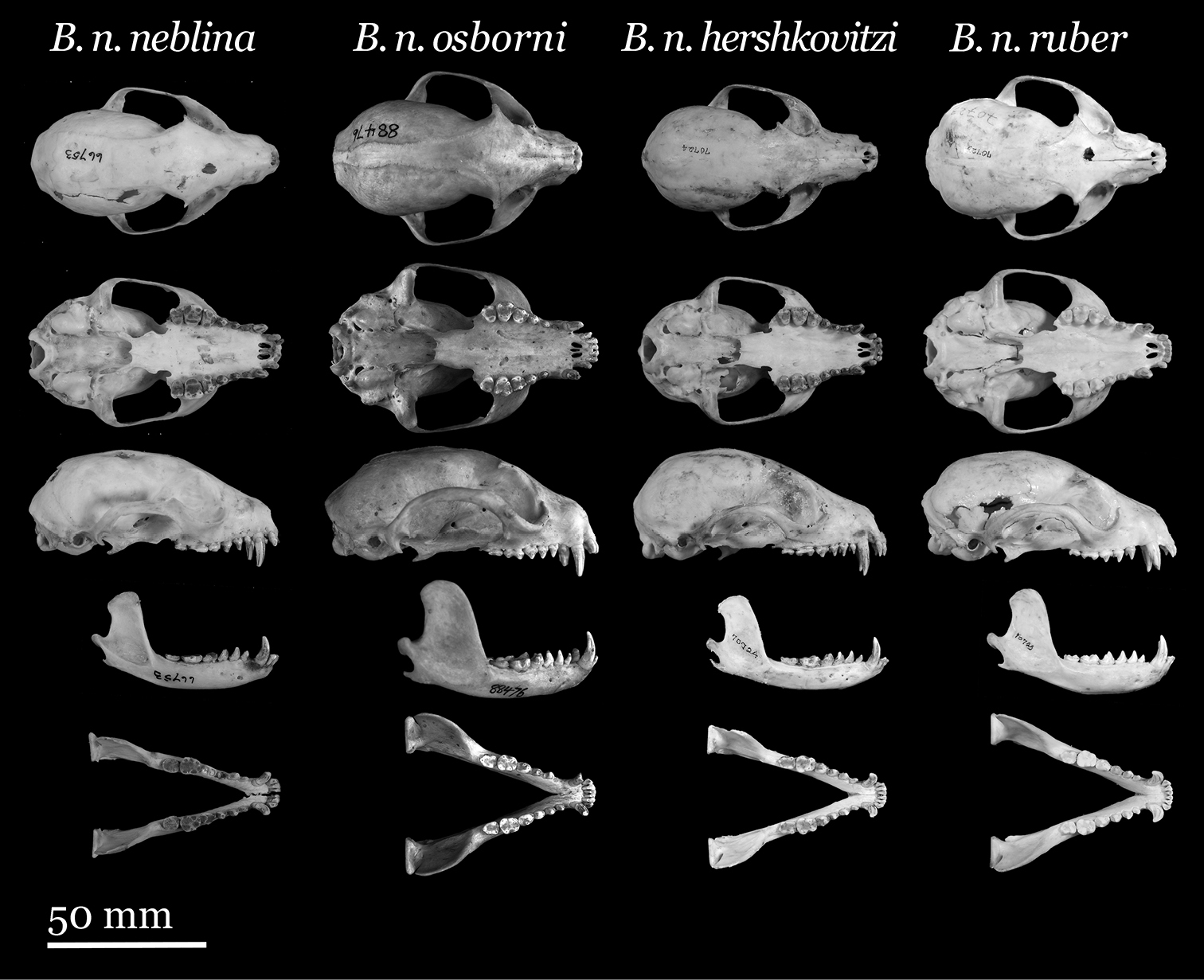
Skulls of Olinguito subspecies. From left to right: Bassaricyon neblina neblina (AMNH 66753, holotype, old adult female, Las Maquinas, Ecuador); Bassaricyon neblina osborni (FMNH 88476, holotype, adult male, Munchique, 2000 m, Cauca Department, Colombia); Bassaricyon neblina hershkovitzi (FMNH 70724, paratype, adult male, San Antonio, Agustin, Huila District, Colombia); Bassaricyon neblina ruber (FMNH 70723, paratype, adult male, Guapantal, 2200 m, Urrao, Antioquia Department, Colombia). Scale bar = 50 mm.
Bioclimatic distribution models and localities for Bassaricyon species. Models from MAXENT using all vouchered occurrence records, 19 bioclimatic variables, and one potential habitat variable.
Predicted distribution for Bassaricyon species based on bioclimatic models. To create these binary maps we used the average minimum training presence for 10 test models as our cutoff. In addition, we excluded areas of high probability that were outside of the known range of the species if they were separated by unsuitable habitat.

The Olinguito, Bassaricyon neblina neblina, in life, in the wild. Taken at Tandayapa Bird Lodge, Ecuador (for mammalogical background of Tandayapa, see Lee et al. 2006). Photograph by Mark Gurney.

Olinguito skins from different regions of the Colombian Andes. Left, Bassaricyon neblina ruber, of the western slopes of the Western Andes of Colombia (FMNH 70722, adult male); Middle, Bassaricyon neblina hershkovitzi, of the eastern slopes of the Central Andes of Colombia (FMNH 70727, adult female); Right, Bassaricyon neblina osborni, of the eastern slopes of the Western Andes and eastern slopes of the Central Andes of Colombia (FMNH 90052, adult female).

The Olinguito, Bassaricyon neblina osborni, in life. Photograph taken in captivity, at the Louisville Zoo (see Poglayen-Neuwall 1976). This animal, named “Ringerl”, was received as an adult in 1967 from the mountains of Colombia near Cali, and exhibited in various zoos, including the National Zoo in Washington, D.C. (see text). Photographs by I. Poglayen-Neuwall, previously unpublished (additional photographs published by Poglayen-Neuwall 1976).
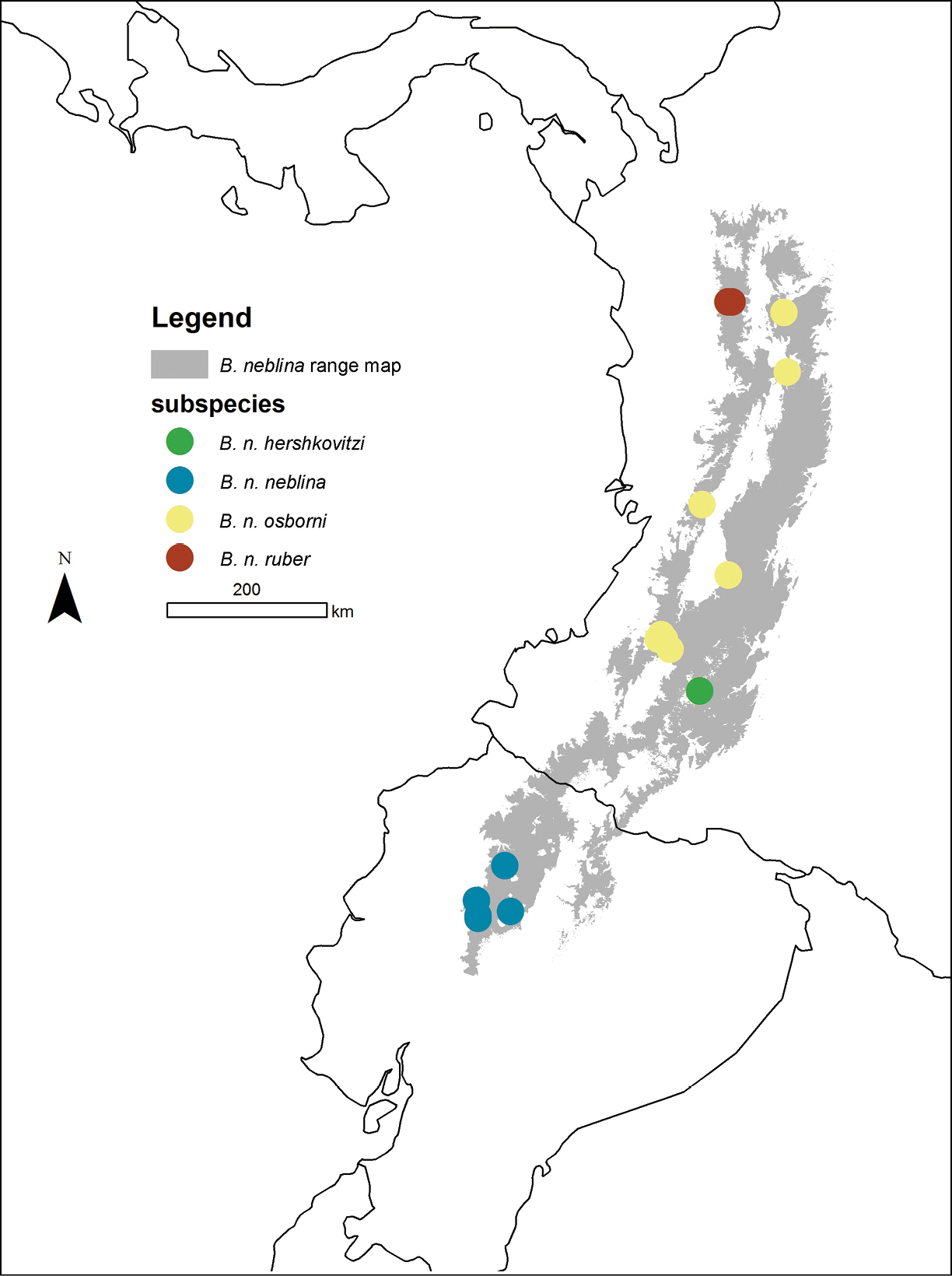
Distributions (localities) of the four Olinguito subspecies in the Andes of Colombia and Ecuador.

Area of sympatric occurrence between Bassaricyon species in western Ecuador. Farmland cutting into cloud forest habitat at Las Pampas, approximately 1800 m, on the western slopes of the Western Andes, Ecuador, along the boundary of Otonga, a protected forest reserve. It is at this elevational and environmental boundary that Bassaricyon medius medius (lower elevations, including more anthropogenically disturbed habitats) and Bassaricyon neblina neblina (higher elevations, less disturbed forests) co-occur in regional sympatry on the western slopes of the Andes.

Type series of an Olinguito subspecies, Bassaricyon neblina hershkovitzi, in the field. Two Olinguito specimens (FMNH 70726, paratype of hershkovitzi, and FMNH 70727, holotype of hershkovitzi, along with a Long-tailed weasel, Mustela frenata, FMNH 70998) brought in by a local hunter, 6 September 1951, at San Antonio, San Agustín, Huila District, Colombia. Photo by P. Hershkovitz, courtesy of the Field Museum of Natural History.
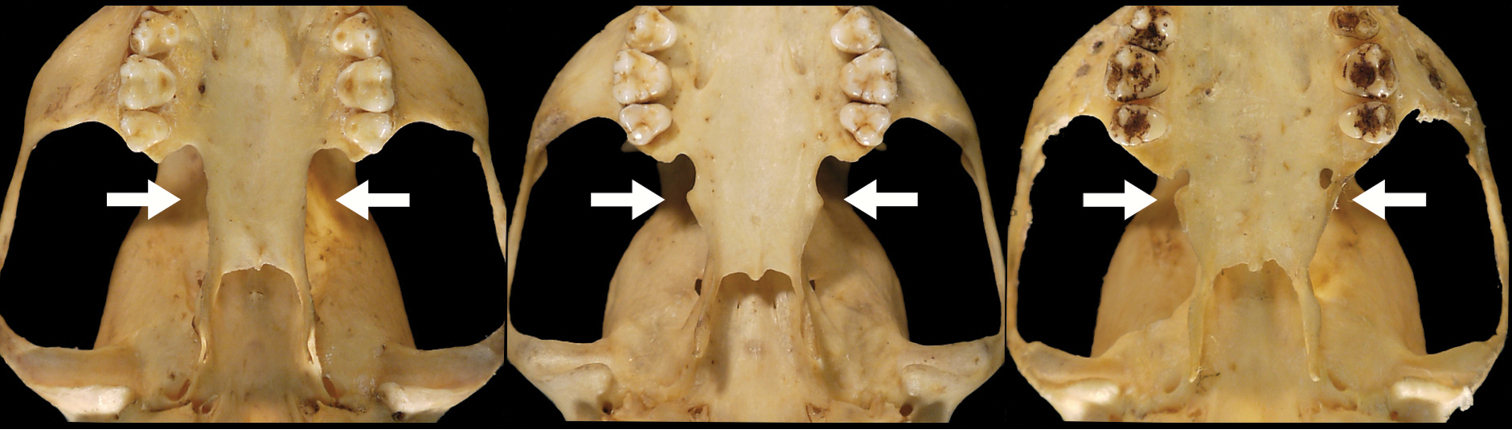
Lateral flare of the postpalatal shelf in Bassaricyon. Lateral extension of the postpalatal shelf (shown by white arrows) is usually absent or little-developed in other Bassaricyon (e.g., left, Bassaricyon alleni, FMNH 41501), but is well-developed in Bassaricyon neblina (e.g., center, Bassaricyon neblina ruber, FMNH 70721, and right, Bassaricyon neblina hershkovitzi, FMNH 70726).

Northern Olingo, Bassaricyon gabbii, in life, in the wild. Photographed at Monteverde, Costa Rica by Greg Basco (left) and Samantha Burke (right).

Eastern Lowland Olingo, Bassaricyon alleni, in life, in the wild. Top, photographed at night (accentuating the dark tones in the pelage) at La Esperanza (Distrito de Yambrasbamba, Provincia de Bongará, Departamento Amazonas), 2000 m, northern Peru; Middle, color camera trap photo in forest canopy, from confluence of the Camisea and Urubamba Rivers (11°42'S, 72°48'W). Peru; Bottom, infrared camera trap photo in forest canopy (same locality as middle photo), showing an olingo carrying a baby in its mouth. Top photograph by César M. Aguilar; middle and bottom camera trap photos courtesy of Smithsonian Conservation Biology Institute.

Western Lowland Olingo, Bassaricyon medius medius, in life. A wild animal photographed under studio conditions at Las Pampas, adjacent to Otonga Reserve, Ecuador. Photographs courtesy of P. Asimbaya and L. Velásquez.

Western Lowland Olingo, Bassaricyon medius orinomus, in life. Wild animals captured, radio-collared, released, and studied by Roland Kays in Limbo Plot, Pipeline Road, Gamboa, Panama (Kays 2000). Photographs courtesy of M. Guerra and R. Kays.
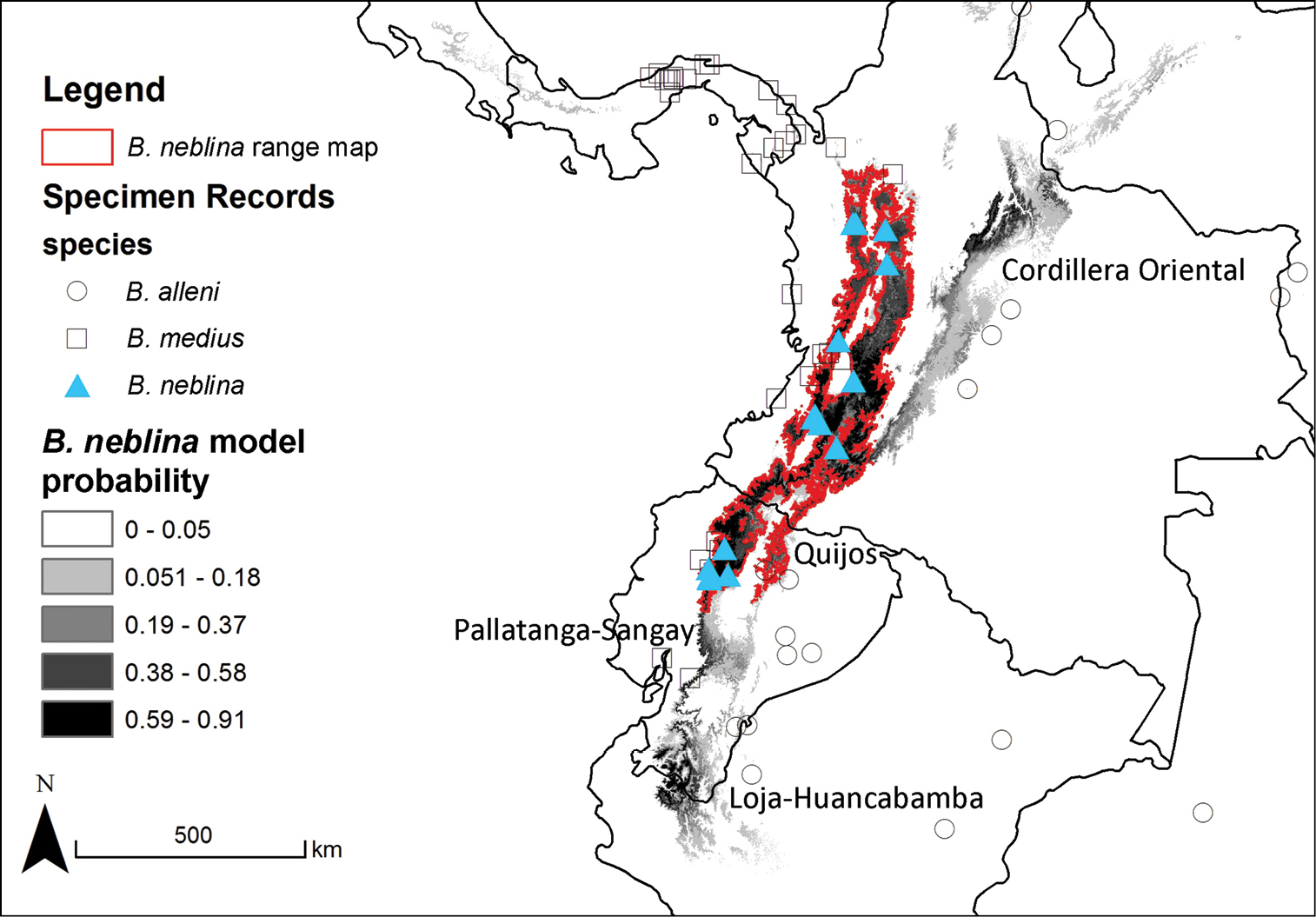
Selected priority areas to search for Olinguitos. Areas mentioned in the text with appropriate cloud forest habitats. A) Cordillera Oriental, the eastern branch of the Andes in Colombia. B) Quijos, a county on the eastern side of the Andes in Ecuador. C) Pallatanga-Sangay in central Ecuador. D) The Loja-Huancabamba region of the Andes in southern Ecuador and northern Peru.