Monograph |
|
Corresponding author: Chris A. Hamilton ( hamiltonlab@uidaho.edu ) Academic editor: Jeremy Miller
© 2016 Chris A. Hamilton, Brent E. Hendrixson, Jason E. Bond.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Hamilton CA, Hendrixson BE, Bond JE (2016) Taxonomic revision of the tarantula genus Aphonopelma Pocock, 1901 (Araneae, Mygalomorphae, Theraphosidae) within the United States. ZooKeys 560: 1-340. https://doi.org/10.3897/zookeys.560.6264
|
Abstract
This systematic study documents the taxonomy, diversity, and distribution of the tarantula spider genus Aphonopelma Pocock, 1901 within the United States. By employing phylogenomic, morphological, and geospatial data, we evaluated all 55 nominal species in the United States to examine the evolutionary history of Aphonopelma and the group’s taxonomy by implementing an integrative approach to species delimitation. Based on our analyses, we now recognize only 29 distinct species in the United States. We propose 33 new synonymies (A. apacheum, A. minchi, A. rothi, A. schmidti, A. stahnkei = A. chalcodes; A. arnoldi = A. armada; A. behlei, A. vogelae = A. marxi; A. breenei = A. anax; A. chambersi, A. clarum, A. cryptethum, A. sandersoni, A. sullivani = A. eutylenum; A. clarki, A. coloradanum, A. echinum, A. gurleyi, A. harlingenum, A. odelli, A. waconum, A. wichitanum = A. hentzi; A. heterops = A. moderatum; A. jungi, A. punzoi = A. vorhiesi; A. brunnius, A. chamberlini, A. iviei, A. lithodomum, A. smithi, A. zionis = A. iodius; A. phanum, A. reversum = A. steindachneri), 14 new species (A. atomicum sp. n., A. catalina sp. n., A. chiricahua sp. n., A. icenoglei sp. n., A. johnnycashi sp. n., A. madera sp. n., A. mareki sp. n., A. moellendorfi sp. n., A. parvum sp. n., A. peloncillo sp. n., A. prenticei sp. n., A. saguaro sp. n., A. superstitionense sp. n., and A. xwalxwal sp. n.), and seven nomina dubia (A. baergi, A. cratium, A. hollyi, A. mordax, A. radinum, A. rusticum, A. texense). Our proposed species tree based on Anchored Enrichment data delimits five major lineages: a monotypic group confined to California, a western group, an eastern group, a group primarily distributed in high-elevation areas, and a group that comprises several miniaturized species. Multiple species are distributed throughout two biodiversity hotspots in the United States (i.e., California Floristic Province and Madrean Pine-Oak Woodlands). Keys are provided for identification of both males and females. By conducting the most comprehensive sampling of a single theraphosid genus to date, this research significantly broadens the scope of prior molecular and morphological investigations, finally bringing a modern understanding of species delimitation in this dynamic and charismatic group of spiders.
Keywords
Biodiversity, New species, Conservation, Molecular systematics, DNA taxonomy, DNA barcoding, Spider taxonomy
Introduction
The family Theraphosidae (tarantulas, baboon spiders, earth tigers) is the most diverse lineage (
Morphology-based phylogenies of mygalomorph spiders have revealed widespread patterns of homoplasy among traditional taxonomic characters (
Distributed across two major biogeographic realms (Nearctic and Neotropical), Aphonopelma species are distributed across the southern third of the United States, ranging west of the Mississippi River to California and south through Mexico and Central America (Fig.

Breadth of diversity of Aphonopelma species habitat types across the United States. A grassland prairie, Otero Co., Colorado B high-elevation pine/conifer in Coconino Co., Arizona C mid-elevation oak woodland throughout the “sky islands” of southeastern Arizona (e.g. Madera Canyon in the Santa Rita Mountains) D grass/oak foothills of the Sierra Nevada Mountains, Mariposa Co., California EZion National Park, Utah F Chihuahuan Desert below the Chiricahua Mountains in Cochise Co., Arizona G Sonoran Desert in Pinal Co., Arizona H Mojave Desert in San Bernardino Co., California I Tamaulipan thornscrub in Starr Co., Texas.

Representation of Aphonopelma burrows in different habitats across the United States. A–C a typical “scrape” (burrow under rock) of A. hentzi in rocky habitat across their distribution D–E a turreted mound around the burrow of A. icenoglei (also A. atomicum, A. mojave, and A. prenticei) F the distinct crescent mound burrow of A. paloma G–I typical free-standing burrows of A. chalcodes, A. eutylenum, A. iodius, or A. johnnycashi in desert, grassland, or rocky habitats.
The taxonomy of Aphonopelma is beset with poorly delimited species boundaries and very few specimens can be confidently identified using published keys (e.g.,
Aphonopelma has a complicated nomenclatural history.
The discovery of species provides the crucial first step in the ongoing pursuit to understand the evolutionary patterns and processes shaping the biodiversity landscape. With over 250 years of taxonomic work behind us, ~1.2 million species have been described - with an estimated 7–10 million species on Earth still remaining to be described (
Abbreviations
Institutional
AUMNH Auburn UniversityMuseum of Natural History; Auburn, Alabama
BMNH The Natural History Museum, London; London, England
Quantitative morphological landmarks (Fig. 3 )
Cl length of the carapace
Cw width of the carapace
LBl labial length
LBw labial width
F1 femur I length (retrolateral aspect)
F1w femur I width
P1 patella I length
T1 tibia I length
M1 metatarsus I length
A1 tarsus I length
F3 femur III length (prolateral aspect)
F3w femur III width
P3 patella III length
T3 tibia III length
M3 metatarsus III length
A3 tarsus III length
F4 femur IV length (prolateral aspect)
F4w femur IV width
P4 patella IV length
T4 tibia IV length
M4 metatarsus IV length
A4 tarsus IV length
PTl palpal tibia length (retrolateral aspect)
PTw palpal tibia width
SC3 ratio of the extent of metatarsus III scopulation (length of scopulation/ventral length of metatarsus III)
SC4 ratio of the extent of metatarsus IV scopulation (length of scopulation/ventral length of metatarsus IV)

Diagrammatic representation of informative quantitative measurements. A–B carapace, labium length and width C retrolateral palpal tibia length and width D retrolateral lengths of leg 1 femur, patella, tibia, metatarsus, tarsus, and width of femur E prolateral lengths of leg 3 femur, patella, tibia, metatarsus, tarsus, and width of femur F prolateral lengths of leg 4 femur, patella, tibia, metatarsus, tarsus, and width of femur G ventral length of scopulation (line), ventral length of metatarsus III (dashed line) H ventral length of scopulation (line), ventral length of metatarsus IV (dashed line).
Methods
Measurement, characterization, and illustration of morphological features
All material was preserved in 80% ethanol and assigned a unique alphanumeric voucher number (APH_#### or AUMS_####) added to each vial and can be used to cross-reference all images, measurements, and locality data. Quantitative measurements are reported in millimeters and were made with a Leica M165C stereomicroscope using the Leica Application Suite software and a digital camera, or from a Mitutoyo ABSOLUTE Digimatic handheld digital caliper. Appendage measurements were based on left appendages (unless otherwise stated); palpal tibia & leg I - retrolateral, legs III & IV - prolateral, extent of metatarsal scopulation - ventral. Lengths of leg articles were taken from the mid-proximal point of articulation to the mid-distal point of the article (sensu
Quantitative measurements are based on a minimum of five individuals of each sex, when a sufficient number of specimens were available, that represent the geographic, molecular (based on the CO1 data), and morphological breadth of each species across its distribution (i.e., every attempt was made to select specimens that represented the range of sizes available across the distribution). Material Examined sections were generated using the MATex Python script used in
Evaluation of quantitative morphological characters for species diagnoses
Morphometrics that were determined to have non-overlapping ranges were used as features for morphological diagnoses of species. The approach applied here is not without certain problems. For example, the inclusion of additional specimens, or in particular the case of species represented by only one or a few specimens (i.e., A. chiricahua or A. saguaro females), additional collecting could add specimens whose features expand the range of some characters and negate some of the measurements used to diagnose species. Building on the importance of certain morphological features found in
To evaluate morphological variation, we examined the morphospace occupied by each species by plotting measurement ratios in boxplots (e.g., Fig.

Examples of boxplots from male quantitative measurements used in species diagnosis (x-axis represents species). A, B, D are ratios: A carapace length/metatarsus III length B palpal tibia length/metatarsus III length D extent of scopulation on metatarsus IV C is carapace length (a proxy for body size), clearly showing the size differences between the miniature species and all other species (y-axis in mm). Additional boxplots can be viewed in Suppl. material

Taxon sampling, molecular techniques, and phylogenetic analyses
Through extensive fieldwork and a citizen-based science program (in association with the American Tarantula Society, see http://www.atshq.org/articles/found.html), we acquired nearly 1,800 specimens of Aphonopelma and closely related sister taxa, for DNA. Specimens were opportunistically collected throughout the southwestern United States, with every attempt made to gather topotypic material from (or near) the type localities of all nominal species of Aphonopelma in the United States. Of these, A. phasmus Chamberlin, 1940 (type locality Phantom Ranch, Grand Canyon) was the only species for which we were unable to obtain fresh material. Legs were removed from all freshly collected material and preserved in ≥95% ethanol or RNAlaterTM (Qiagen, Valencia, CA, USA) and stored at -80oC. Specimens are deposited at the AUMNH, with select duplicate specimens of novel species deposited at the
Phylogenetic analyses of Aphonopelma relationships were conducted using molecular datasets (mitochondrial and nuclear) employing likelihood optimality criteria. A new mitochondrial dataset drawn from the taxa in
The analyses described above and recent smaller scale molecular studies (
The use of nuclear loci generally has led to an enhanced understanding of the Tree of Life, but unfortunately, many non-model invertebrates (e.g., spiders) lack adequately developed loci for targeted sequencing. Until very recently, few genetic markers were available for inferring spider phylogenies (i.e., only 13 independent markers though 2013, see
Anchored Enrichment data were collected through the Center for Anchored Phylogenomics at Florida State University (http://www.anchoredphylogeny.com) following the general methods of
Utilizing the bioinformatics pipeline described in
Locality data, georeferencing, generation of niche-based distribution models, and conservation status
For all newly collected samples, latitude and longitude were recorded in the field using a Global Positioning System (GPS) receiver (WGS84 datum) in Decimal Degrees (DD). For previously collected museum specimens, locality data were manually georeferenced using Google Earth (Google, Mountain View, CA) or Topo North America (DeLorme, Yarmouth, ME) in Decimal Degrees (DD). All georeferenced and field recorded locality data (latitude, longitude, elevation) were crosschecked by hand in Google Earth, Topo North America, or ArcGIS (ESRI, Redlands, CA) prior to generating distribution maps, niche-based distribution models, and database entry. Distribution maps were constructed in ArcGIS. Because older locality labels often lack sufficient locality information, many georeferenced values are imprecise and should be interpreted with caution. As well, because these older labels are often faded or handwritten, there may be slight discrepancies in the spelling of localities, collectors, etc. Data for labels that document only county and/or town information were georeferenced to the approximate geographic center of the given locality. Precision for each georeferenced point is annotated as a superscript in the material examined section for each species using the confidence value scheme employed by
As an approach to facilitate species discovery and delimitation, niche-based distribution models (DMs) were constructed for species for which sufficient locality data were available (i.e., at least 10 different localities separated by at least 1 km). Niche-based DMs provide estimates for the probability of finding a species at a location on the landscape given the set of correlate ecological and climatic parameters used to construct the model. Locality coordinates for each specimen were imported into ArcMap (ESRI, Redlands, CA) and converted into shapefiles. Following the procedures outlined in

A hypothesized conservation status for each species has been included with each respective description. The designations provided herein are not based on any formal calculations and therefore should not be viewed as formal status declarations. These designations are based upon our own extensive fieldwork and the observations of fellow arachnologists (e.g., Tom Prentice and Wendell Icenogle). As such, our estimation of the conservation status for each species is likely conservative.
Species delimitation and conceptualization
The species concept used throughout this taxonomic revision utilizes the ideas of de Queiroz’s Unified Species Concept (2005). Where possible, we employ a combination of molecular phylogenetic, morphological, behavioral, and biogeographic evidence to identify independently evolving lineages. If not possible, an alternative species concept is noted. Initial species hypotheses were based on the recognized morphological species hypotheses (
Data resources
All data (molecular, morphological, geographic, and images) used to establish these species hypotheses have been deposited in the Dryad Data Repository (https://doi.org/10.5061/dryad.k6c82). All locality data underpinning the analysis reported in this paper are deposited at GBIF, the Global Biodiversity Information Facility, http://ipt.pensoft.net/resource?r=aphonopelma. All morphological images have been deposited in Morphbank, and can be viewed by referencing the "APH-S" Specimen External Identifier. Additional specimen data, species plates, and morphological data can be found in the Suppl. materials.
Results and discussion
Summary of taxonomic diversity
Prior to our work leading up to this taxonomic revision (
Aphonopelma phylogeny
Following the integrative approach for delimiting species within Aphonopelma outlined in

Maximum Likelihood inferred CO1 gene tree phylogeny for the 1,032 Aphonopelma specimen dataset. Species delimitations followed the integrative methodological approach outlined in
To further assess species-level diversity and evaluate the associated hypothesized species boundaries delimited by the CO1 data, we sequenced multiple specimens per putative species using Anchored Enrichment. A dataset of 455 loci (229,854 basepairs) across 80 OTUs produced a highly resolved species-level phylogeny of all the major species groups and regional clades within Aphonopelma (Fig.

Species tree of all United States Aphonopelma, inferred from the 455 loci Anchored Enrichment dataset. Species delimitations correspond to our final integrative approach outlined herein. Black circles denote 100% bootstrap support; black squares denote bootstrap support between 99–80%; white squares denote bootstrap support less than 80%. Node support values = based on the RAxML bootstrap support from all trees and all loci. All genealogically exclusive species are identified with a grey bar; A. iodius, a paraphyletic species as presently defined, is identified by the black boxes. All major species groups are identified by colored boxes.
While our methodological approach using CO1 identified the broad effectiveness of this 900 basepair fragment of mtDNA to identify known species and help illuminate species boundaries across the most comprehensive molecular sampling of a single spider genus to date, the AE nuDNA data robustly highlight where deep mitochondrial divergences and introgression have obscured our understanding of the true evolutionary history of these lineages. It is important to point out that a number of the putative mitochondrial species corresponded to lineages that were considered cryptic species in
Steindachneri species group
The Steindachneri species group presently includes a single species (A. steindachneri) from California. The CO1 and AE datasets both support the “basal” position of A. steindachneri as the sister taxon to all other species of Aphonopelma in the United States (Figs
Western species group diversity (Iodius species group)
The western group of species (note: the western designation is only applied here in an informal sense because other lineages are distributed west of the Cochise Filter Barrier) is comprised entirely of the Iodius species group (A. chalcodes, A. eutylenum, A. iodius, and A. johnnycashi sp. n.) (Fig.
Unfortunately, the mitochondrial data are complex. Results from the CO1 analysis confounds our understanding of species boundaries and relationships in Aphonopelma, likely due to mitochondrial introgression or deep haplotype conservation (e.g., one mtDNA lineage of A. chalcodes is sister to A. vorhiesi, a species that belongs to the Marxi species group, while another mtDNA lineage of A. chalcodes is sister to other A. iodius lineages). Alternatively, the AE nuclear data provide support for these species (A. chalcodes, A. vorhiesi) as independently evolving and monophyletic lineages. In previous analyses (
Eastern species group diversity (Hentzi and Moderatum species groups)
The eastern group of species is designated as such because the six species are largely distributed east of the Cochise Filter Barrier. This monophyletic lineage is strongly supported and includes the Hentzi species group (A. anax, A. armada, and A. hentzi) and Moderatum species group (A. gabeli, A. moderatum, and A. moellendorfi sp. n.) (Fig.
When reviewing both the putative mitochondrial and cryptic species identified in
Sky islands diversity (Marxi species group)
The Marxi species group includes A. catalina sp. n., A. chiricahua sp. n., A. madera sp. n., A. marxi, A. peloncillo sp. n., and A. vorhiesi (Fig.
Despite our extensive collecting throughout this region, there likely still remains undescribed diversity in this species group (Figs
Miniaturization and diversification in Aphonopelma (Paloma species group)
The Paloma species group includes a dozen miniaturized species primarily located in the Mojave and Sonoran deserts: A. atomicum sp. n., A. icenoglei sp. n., A. joshua, A. mareki sp. n., A. mojave, A. paloma, A. parvum sp. n., A. phasmus, A. prenticei sp. n., A. saguaro sp. n., A. superstitionense sp. n., and A. xwalxwal sp. n. (Fig.
The CO1 data can be used to confidently identify the species in this group (probably due to the relatively high mtDNA divergence between species), except A. mareki and A. superstitionense due to putative mitochondrial introgression. Unfortunately, interspecific relationships are not well supported and the placement of A. paloma and A. xwalxwal is problematic (i.e., not consistently recovered in the same place in the phylogeny). When the AE data are employed, the Paloma species group is strongly supported and the placement of A. paloma and A. xwalxwal is confidently resolved. Most relationships within and between the remaining species are also strongly supported and highly resolved (Fig.
Body size is one of the most important determinants of an organism’s ecological role (
Species considered nomina dubia
The species Aphonopelma cratium Chamberlin, 1940 is based on a male holotype and female allotype (both
The description of Eurypelma mordax Ausserer, 1871 was based on a single adult male but the holotype was likely destroyed during World War II (see
Two different type localities (Ft. Yuma and Williams, Arizona) were listed in the description of Eurypelma rusticum Simon, 1891 but it is unclear which specimen was actually used for the description (
The enigmatic species Delopelma radinum (Chamberlin & Ivie, 1939) was described on the basis of a single adult male collected from Manhattan Beach, California in November 1937. To our knowledge, no specimens comparable to the badly fragmented holotype (
Taxonomy
Family Theraphosidae Thorell, 1869
Subfamily Theraphosinae Thorell, 1870
Aphonopelma
Rhechostica Simon, 1892: 162 (type species by original designation Homoeomma texense Simon, 1891). Suppressed as a senior synonym of Aphonopelma by ICZN Opinion 1637.
Dugesiella Pocock, 1901: 551 (type species by original designation Dugesiella crinita Pocock, 1901). First synonymized with Rhechostica by
Aphonopelma Pocock, 1901: 553 (type species by original designation Eurypelma seemanni Pickard-Cambridge, 1897). First synonymized with Rhechostica by
Delopelma Petrunkevitch, 1939: 567 (type species by original designation Eurypelma marxi Simon, 1891). First synonymized with Rhechostica by
Gosipelma Chamberlin, 1940: 4 (type species by original designation Gosipelma angusi Chamberlin, 1940). Originally described as a subgenus of Aphonopelma, but never elevated to full generic status. First synonymized with Rhechostica by
Chaunopelma Chamberlin, 1940: 30 (type species by original designation Delopelma radinum Chamberlin & Ivie, 1939). First synonymized with Rhechostica by
Apachepelma Smith, 1994: 45 (type species by original designation Aphonopelma paloma Prentice, 1992). First synonymized with Aphonopelma by
Type species
Eurypelma seemanni F.O. Pickard-Cambridge, 1897; female holotype from Puerto Culebra, Pacific coast, W of Liberia, Guanacaste province, Costa Rica, coll. Dr. Seeman; deposited in BMNH. [examined]
Eurypelma seemanni F. O. Pickard-Cambridge, 1897: 26.
Aphonopelma seemanni Pocock, 1901: 553.
Aphonopelma seemanni Valerio, 1980: 274.
Rhechostica seemanni Raven, 1985: 149.
Aphonopelma seemanni Smith, 1995: 141.
Diagnosis
From
“The genus Aphonopelma is distinguished from all other theraphosid genera by the following combination of characters: (1) no known external stridulation organs; (2) hair-like or spiniform plumose setae on the prolateral surface of the trochanter and femur of leg I and on the retrolateral surface of the coxa and trochanter of the pedipalp; (3) type I urticating setae only; (4) corresponding segments of all legs approximately the same width in females (femur III in males sometimes laterally swollen); (5) scopula of tarsus IV usually entire, if divided then only partially and narrowly by line of setae; (6) setae on the prolateral surface of coxa I hair-like and not basally swollen, spiniform and basally swollen, or distinctly stout and thorn-like; (7) metatarsus I flexing against the lower process of the tibial spur, with either the apex of the spur contacting the ventral surface of the metatarsus or the outer edge of the spur in the apical half contacting the prolateral surface of the metatarsus; (8) and the lower process of the tibial spur curving prolaterodistally and widening apically, usually equipped with at least one apical or preapical megaspine, and the upper shorter process less stout basally, relatively uniform in diameter throughout its length, and equipped on its inner surface with at least one stout, basally articulated megaspine.”
Future nomenclatural status of Aphonopelma
Due to our inability to conduct fieldwork in Mexico and Central America, a comprehensive taxonomic revision of Aphonopelma (and other closely related genera) is not feasible at this time. However, we do anticipate future genus-level nomenclatural changes for those species found in the United States. As presently defined, we have reason to believe that Aphonopelma is not monophyletic. Our choice of outgroups places the Aphonopelma species from the United States closer to the genus Sericopelma than to the Central American species of Aphonopelma (Fig.
Aphonopelma
within the United States
Aphonopelma anax (Chamberlin, 1940)
Aphonopelma armada (Chamberlin, 1940)
Aphonopelma atomicum Hamilton, sp. n.
Aphonopelma catalina Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma chalcodes Chamberlin, 1940
Aphonopelma chiricahua Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma eutylenum Chamberlin, 1940
Aphonopelma gabeli Smith, 1995
Aphonopelma hentzi (Girard, 1852)
Aphonopelma icenoglei Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma iodius (Chamberlin & Ivie, 1939)
Aphonopelma johnnycashi Hamilton, sp. n.
Aphonopelma joshua Prentice, 1997
Aphonopelma madera Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma mareki Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma marxi (Simon, 1891)
Aphonopelma moderatum (Chamberlin & Ivie, 1939)
Aphonopelma moellendorfi Hamilton, sp. n.
Aphonopelma mojave Prentice, 1997
Aphonopelma paloma Prentice, 1993
Aphonopelma parvum Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma peloncillo Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma phasmus Chamberlin, 1940
Aphonopelma prenticei Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma saguaro Hamilton, sp. n.
Aphonopelma steindachneri (Ausserer, 1875)
Aphonopelma superstitionense Hamilton, Hendrixson & Bond, sp. n.
Aphonopelma vorhiesi (Chamberlin & Ivie, 1939)
Aphonopelma xwalxwal Hamilton, sp. n.
Key to the male Aphonopelma of the United States
| Arizona | A1 | |
| California | B1 | |
| Colorado, Kansas, Oklahoma, Missouri, Arkansas, and Louisiana | C1 | |
| Nevada and Utah | D1 | |
| New Mexico | E1 | |
| Texas | F1 |
A1. Key to the male Aphonopelma of Arizona
| 1 | Stout setae on the prolateral surface of coxa I; distribution restricted to southern Greenlee County | Aphonopelma hentzi |
| – | No stout setae on the prolateral surface of coxa I; widespread distribution | 2 |
| 2 | Small species (Cl ≤ 9 mm); generally found in desert, grassland, and/or chaparral habitats | 3 |
| – | Small to medium-sized species (Cl 6–12 mm); generally distributed throughout mid- to high-elevation forested habitats; possessing a black carapace – never blonde, tan, or brown | 5 1 |
| – | Medium to large-sized species (Cl ≥ 9 mm) found in a variety of habitats (but seldom found in higher-elevation forested habitats); carapace blonde, tan, brown, or black | 6 |
| 3 | Possessing a swollen or slightly swollen femur III | 4 1 |
| – | Lacks a swollen femur III; distributed east of Tucson in Cochise, Graham, and Greenlee Counties | Aphonopelma parvum |
| 4 | Distributed across the Mojave Desert and adjacent sections of the Sonoran Desert in western Arizona | Aphonopelma prenticei |
| – | Distributed across lower-elevation sections of the Sonoran Desert in southern Arizona | Aphonopelma paloma |
| – | Distributed across mid- to high-elevation chaparral and forested habitats north of the Phoenix Metropolitan Area | Aphonopelma mareki |
| – | Distributed at the bottom of Grand Canyon in the vicinity of Phantom Ranch | Aphonopelma phasmus |
| – | Distributed across the southern foothills and canyons of the Santa Catalina and Rincon Mountains east of Tucson | Aphonopelma saguaro |
| – | Distributed east of the Phoenix Metropolitan Area in the foothills and canyons of the Superstition Mountains | Aphonopelma superstitionense |
| 5 | Distributed north and east of the Phoenix Metropolitan Area across the Colorado Plateau and on isolated mountains (e.g., Four Peaks, Mount Ord) | Aphonopelma marxi |
| – | Distributed across the Santa Catalina Mountains | Aphonopelma catalina |
| – | Distributed across the Huachuca, Pajarito, Patagonia, and Santa Rita Mountains | Aphonopelma madera |
| – | Distributed across the Chiricahua Mountains | Aphonopelma chiricahua |
| 6 | Carapace blonde, tan, or brown | 7 |
| – | Carapace black | 8 |
| 7 | Distributed across the Colorado Plateau north of the Colorado River | Aphonopelma iodius |
| – | Widespread throughout Arizona but never north of the Colorado River | Aphonopelma chalcodes |
| 8 | PT1/M1 ≤ 0.68 | Aphonopelma gabeli |
| – | PT1/M1 ≥ 0.74 | Aphonopelma peloncillo and Aphonopelma vorhiesi2 |
B1. Key to the male Aphonopelma of California
| 1 | Metatarsus IV scopulation ≥ 60% | 2 |
| – | Metatarsus IV scopulation ≤ 60% | 4 |
| 2 | Possessing a black carapace; distributed across the plains and foothills of the western Sierra Nevada Mountains | Aphonopelma johnnycashi |
| – | Possessing a tan or brown carapace | 3 1 |
| 3 | Distributed west of the Mojave Desert, from the western part of the Transverse Range and down the southern California coast, along the Peninsular Ranges | Aphonopelma eutylenum |
| – | Distributed from the Bay Area south along the Coast Ranges, west of the Central Valley, across the Transverse Range and into the Mojave Desert | Aphonopelma iodius |
| 4 | Possessing stout setae on the medial surface of the sternum | 5 |
| – | Lacks stout setae on the medial surface of the sternum | 6 |
| 5 | F4/T4 ≤ 1.05; distributed across the mountains and foothills west of the Coachella Valley and south to the Borrego Springs area; fall breeding period | Aphonopelma xwalxwal |
| – | F4/T4 ≥1.07; distributed in and around Joshua Tree National Park; summer breeding period | Aphonopelma joshua |
| 6 | Medium to large-sized species (Cl ≥ 9 mm); distributed across southwestern California but not within the Mojave Desert | Aphonopelma steindachneri |
| – | Small species (Cl ≤ 9mm); distributed across the Mojave Desert | 7 |
| 7 | Femur III swollen or slightly swollen | 8 |
| – | Femur III not swollen | 9 1 |
| 8 | A1/F3 ≥ 0.58; distributed across the Panamint Range and eastern San Bernardino County | Aphonopelma prenticei |
| – | A1/F3 ≤0.56; narrowly distributed across the Amargosa Range in southeastern Inyo County | Aphonopelma atomicum |
| 9 | Distributed across the northwestern Mojave Desert in eastern Kern and northwestern San Bernardino Counties | Aphonopelma mojave |
| – | Distributed throughout the southern Mojave Desert along the foothills on the northern side of the San Gabriel and San Bernardino Mountains, east to the Coxcomb Mountains and south into Joshua Tree National Park, north to Kramer Junction and Barstow | Aphonopelma icenoglei |
C1. Key to the male Aphonopelma of Colorado, Kansas, Oklahoma, Missouri, Arkansas, and Louisiana
| 1 | Carapace tan, brown, or copper; possessing stout setae on the prolateral surface of coxa I; distributed east of the Rocky Mountains across Colorado, Kansas, Oklahoma, Missouri, Arkansas, and Louisiana | Aphonopelma hentzi |
| – | Carapace black; lacks stout setae on the prolateral surface of coxa I; distributed west of the Rocky Mountains across southwestern Colorado | Aphonopelma marxi |
D1. Key to the male Aphonopelma of Nevada and Utah
| 1 | Metatarsus IV scopulation ≥ 60%; widespread distribution | Aphonopelma iodius |
| – | Metatarsus IV scopulation ≤ 60% | 2 |
| 2 | Distributed across the Colorado Plateau in southeastern Utah | Aphonopelma marxi |
| – | Distributed across the Mojave Desert in southwestern Utah and/or southern Nevada | 3 1 |
| 3 | A1/F3 ≥ 0.58; widespread across the northeastern Mojave Desert | Aphonopelma prenticei |
| – | A1/F3 ≤ 0.58; distribution restricted to the Amargosa Valley and Nevada Test Site near Mercury, NV | Aphonopelma atomicum |
E1. Key to the male Aphonopelma of New Mexico
| 1 | Medium to large-sized species (Cl ≥ 9 mm) | 2 |
| – | Small species (Cl ≤ 9 mm) distributed across the extreme southwestern part of the state (Hidalgo County) | Aphonopelma parvum |
| 2 | Possessing stout setae on the prolateral surface of coxa I; possessing a tan, brown, or copper carapace | Aphonopelma hentzi |
| – | Lacks stout setae on the prolateral surface of coxa I; possessing a black carapace | 3 |
| 3 | Distributed across the northern half of the state and western part of the Rockies, inhabiting high-elevation pine forest and sagebrush steppe | Aphonopelma marxi |
| – | Distributed across the southern half of the state | 4 |
| 4 | PT1/M1 ≤ 0.68 | Aphonopelma gabeli |
| – | PT1/M1 ≥ 0.74 | 5 3 |
| 5 | Distribution restricted to the southeastern Peloncillo Mountains; males active mostly during mid-summer | Aphonopelma peloncillo |
| – | Distribution widespread across southern New Mexico; males active mostly during late summer and early fall | Aphonopelma vorhiesi |
F1. Key to the male Aphonopelma of Texas
| 1 | Possessing a short and stout embolus; distributed across South Texas | Aphonopelma anax |
| – | Possessing a long, thin, and tapering embolus; widespread | 2 |
| 2 | Possessing a tan, brown, or copper carapace | 3 |
| – | Possessing a black carapace | 4 |
| 3 | Metatarsus III scopulation ≤ 63%; stout setae on the prolateral surface of coxa I present along dorsal, posterior, and ventral margins, but lacking from the center and anterior margin | Aphonopelma armada |
| – | Metatarsus III scopulation ≥ 69%; stout setae abundant across the prolateral surface of coxa I | Aphonopelma hentzi |
| 4 | M1/M4 ≤ 0.74 | Aphonopelma gabeli |
| – | M1/M4 ≥ 0.75 | Aphonopelma moderatum and Aphonopelma moellendorfi4 |
1 Species that key here are morphologically indistinguishable for the most part but can be identified based on their localities and molecular data.
2 Mature males of these two species are morphologically indistinguishable and cannot be identified using morphological criteria when they co-occur in southeastern Cochise County, but can be differentiated using molecular data. Aphonopelma vorhiesi is likely the correct determination if the specimen originates from Graham, Pima, Pinal, Santa Cruz, or western Cochise Counties.
3 Species that key here are morphologically indistinguishable for the most part but can be identified based on their localities, the timing of their breeding periods, and molecular data.
4 Mature males of these two species are morphologically indistinguishable and cannot be identified using morphological criteria when they co-occur, but can be differentiated using molecular data. Aphonopelma moderatum reaches its northernmost distribution in Val Verde County but is largely distributed to the south along the Rio Grande Valley. Aphonopelma moellendorfi reaches its easternmost distribution in Val Verde County with other specimens having been located in extreme West Texas. When these two species come into syntopy (southeastern Val Verde County), males can easily be distinguished prior to their ultimate molt due to their phenotypic differences: A. moderatum possess distinct alternating black and tan/orange bands on its legs whereas A. moellendorfi is more uniformly brown.
Key to the female Aphonopelma of the United States
| Arizona | A2 | |
| California | B2 | |
| Colorado, Kansas, Oklahoma, Missouri, Arkansas, and Louisiana | C2 | |
| Nevada and Utah | D2 | |
| New Mexico | E2 | |
| Texas | F2 |
A2. Key to the female Aphonopelma of Arizona 1
| 1 | Possessing stout setae on the prolateral surface of coxa I; distribution restricted to southern Greenlee County | Aphonopelma hentzi |
| – | Lacks stout setae on the prolateral surface of coxa I; widespread | 2 |
| 2 | Small species (Cl ≤ 9 mm); generally found in desert, grassland, and/or chaparral habitats | 3 2 |
| – | Small to large-sized species (Cl 7.5–16.5 mm); generally distributed throughout mid- to high-elevation forested habitats; possessing black or gray carapace – never blonde, tan, or brown | 4 2 |
| – | Medium to large-sized species (Cl ≥ 9 mm) found in a variety of habitats (but seldom found in higher-elevation forested habitats); carapace blonde, tan, brown, black, or gray | 6 |
| 3 | Distributed east of Tucson in Cochise, Graham, and Greenlee Counties | Aphonopelma parvum |
| – | Distributed across the Mojave Desert and adjacent sections of the Sonoran Desert in western Arizona | Aphonopelma prenticei |
| – | Distributed across lower-elevation sections of the Sonoran Desert in southern Arizona | Aphonopelma paloma |
| – | Distributed across mid- to high-elevation chaparral and forested habitats north of the Phoenix Metropolitan Area | Aphonopelma mareki |
| – | Distributed across the southern foothills and canyons of the Santa Catalina and Rincon Mountains east of Tucson | Aphonopelma saguaro |
| – | Distributed east of the Phoenix Metropolitan Area in the foothills and canyons of the Superstition Mountains | Aphonopelma superstitionense |
| 4 | T1/T4 ≥ 1.06; distributed north and east of the Phoenix Metropolitan Area across the Colorado Plateau and on isolated mountains (e.g., Four Peaks, Mount Ord) | Aphonopelma marxi |
| – | T1/T4 ≤1.02; distributed across the Madrean Sky Islands in southeastern Arizona | 5 |
| 5 | Distributed across the Santa Catalina Mountains | Aphonopelma catalina |
| – | Distributed across the Huachuca, Pajarito, Patagonia, and Santa Rita Mountains | Aphonopelma madera |
| – | Distributed across the Chiricahua Mountains | Aphonopelma chiricahua |
| 6 | Metatarsus IV scopulation ≥ 55% | 7 |
| – | Metatarsus IV scopulation ≤ 55% | 8 |
| 7 | Distributed across the Colorado Plateau north of the Colorado River | Aphonopelma iodius |
| – | Widespread throughout Arizona, but never north of the Colorado River | Aphonopelma chalcodes |
| 8 | Possessing spermathecae with capitate bulbs | Aphonopelma peloncillo and Aphonopelma vorhiesi3 |
| – | Possessing short, wide, and slightly rounded spermathecae without capitate bulbs | Aphonopelma gabeli |
B2. Key to the female Aphonopelma of California 4
| 1 | Metatarsus IV scopulation ≥ 55% | 2 2 |
| – | Metatarsus IV scopulation ≤ 55% | 3 |
| 2 | Distributed across the plains and foothills of the western Sierra Nevada Mountains | Aphonopelma johnnycashi |
| – | Distributed west of the Mojave Desert, from the western part of the Transverse Range and down the southern California coast, along the Peninsular Ranges | Aphonopelma eutylenum |
| – | Distributed from the Bay Area south along the Coast Ranges, west of the Central Valley, across the Transverse Range and into the Mojave Desert | Aphonopelma iodius |
| 3 | Large species (Cl ≥ 9 mm); distributed across southwestern California but not within the Mojave Desert | Aphonopelma steindachneri |
| – | Small species (Cl ≤ 9 mm); distributed across the Mojave and portions of the Sonoran (Colorado) Deserts | 4 2 |
| 4 | Distributed across the northwestern Mojave Desert in eastern Kern and northwestern San Bernardino Counties | Aphonopelma mojave |
| – | Distributed in and around Joshua Tree National Park; F1/M4 ≤ 1.04; L1/Cl ≤ 2.87 | Aphonopelma joshua 5 |
| – | Distributed across the Panamint Range and eastern San Bernardino County | Aphonopelma prenticei |
| – | Distributed across the Amargosa Range in southeastern Inyo County | Aphonopelma atomicum |
| – | Distributed throughout the southern Mojave Desert along the foothills on the northern side of the San Gabriel and San Bernardino Mountains, east to the Coxcomb Mountains and south into Joshua Tree National Park, north to Kramer Junction and Barstow; F1/M4 ≥ 1.07; L1/Cl ≥ 2.92 | Aphonopelma icenoglei 5 |
C2. Key to the female Aphonopelma of Colorado, Kansas, Oklahoma, Missouri, Arkansas, and Louisiana
| 1 | Carapace tan or brown; possessing stout setae on the prolateral surface of coxa I; distributed east of the Rocky Mountains across Colorado, Kansas, Oklahoma, Missouri, Arkansas, and Louisiana | Aphonopelma hentzi |
| – | Carapace black; lacks stout setae on the prolateral surface of coxa I; distributed west of the Rocky Mountains across southwestern Colorado | Aphonopelma marxi |
D2. Key to the female Aphonopelma of Nevada and Utah
| 1 | Metatarsus IV scopulation ≥ 60%; widespread distribution | Aphonopelma iodius |
| – | Metatarsus IV scopulation ≤ 50% | 2 |
| 2 | Distributed across the Colorado Plateau in southeastern Utah | Aphonopelma marxi |
| – | Distributed across the Mojave Desert in southwestern Utah and/or southern Nevada | 3 2 |
| 3 | Cl/M4 ≥ 1.31; widespread across the northeastern Mojave Desert | Aphonopelma prenticei |
| – | Cl/M4 ≤ 1.28; distribution restricted to the Amargosa Valley and Nevada Test Site near Mercury, NV | Aphonopelma atomicum |
E2. Key to the female Aphonopelma of New Mexico
| 1 | Possessing spermathecae with capitate bulbs | 2 |
| – | Possessing short, wide, and slightly rounded spermathecae without capitate bulbs; anterior margin of carapace broad, associated with robust chelicerae | Aphonopelma gabeli |
| 2 | Possessing stout setae on the prolateral surface of coxa I; possessing a tan or brown carapace | Aphonopelma hentzi |
| – | Lacks stout setae on the prolateral surface of coxa I; possessing a black or gray carapace | 3 |
| 3 | Distributed across the northern half of the state and western part of the Rockies, inhabiting high-elevation pine forest and sagebrush steppe | Aphonopelma marxi |
| – | Distributed across the southern half of the state | 4 |
| 4 | Medium to large-sized species (Cl ≥ 10 mm) | 5 2 |
| – | Small species (Cl ≤ 9 mm); distributed in the extreme southwestern part of the state (Hidalgo County) | Aphonopelma parvum |
| 5 | Distribution restricted to the southeastern Peloncillo Mountains | Aphonopelma peloncillo |
| – | Distribution widespread across southern New Mexico | Aphonopelma vorhiesi |
F2. Key to the female Aphonopelma of Texas 6
| 1 | Possessing spermathecae with capitate bulbs | 2 |
| – | Possessing short, wide, and slightly rounded spermathecae without capitate bulbs | 4 |
| 2 | Possessing stout setae on the prolateral surface of coxa I; all leg segments uniformly colored brown or black | 3 |
| – | Lacks stout setae on the prolateral surface of coxa I; legs distinctly colored with the femora and tibiae generally orange or tan and the patellae, metatarsi, and tarsi dark brown to black (if the leg segments are uniformly colored, they will be orange or tan, never brown or black) | Aphonopelma moderatum |
| 3 | Stout setae on the prolateral surface of coxa I present along dorsal, posterior, and ventral margins, but lacking from the center and anterior margin; metatarsi I, II, and III distinctly flared; species with overall shiny, lustrous appearance | Aphonopelma armada |
| – | Stout setae abundant across the prolateral surface of coxa I; metatarsi not distinctly flared; species with overall hirsute appearance | Aphonopelma hentzi |
| 4 | Distributed across South Texas; possessing a tan or brown carapace with a large robust appearance | Aphonopelma anax |
| – | Distributed across the Chihuahuan Desert and southwestern High Plains in West Texas; possessing a brownish-gray carapace; anterior margin of carapace broad, associated with robust chelicerae | Aphonopelma gabeli |
1 Adult females of Aphonopelma phasmus remain unknown.
2 Species that key here are morphologically indistinguishable for the most part but can be identified based on their localities and molecular data.
3 Mature females of these two species can be morphologically indistinguishable when they co-occur in southeastern Cochise County. Aphonopelma vorhiesi is likely the correct determination if the specimen originates from Graham, Pima, Pinal, Santa Cruz, or western Cochise Counties.
4 Adult females of Aphonopelma xwalxwal remain unknown.
5Aphonopelma joshua and A. icenoglei are syntopic at various locations in and around Joshua Tree National Park. Females of A. joshua generally can be diagnosed by possessing tarsi IV divided by setae (
6 Adult females of Aphonopelma moellendorfi remain unknown.
Aphonopelma anax
Dugesiella anax Chamberlin, 1940: 34; male holotype and female allotype from Kingsville, Kleberg Co., Texas, 27.515869 -97.8561095, elev. 58ft., no collecting date, coll. Prof. J.C. Cross; 3 female paratypes from Harlingen, Cameron Co., Texas, 26.190631 -97.6961035, elev. 40ft., 1939, coll. Bryce Brown; deposited in
Rhechostica anax Raven, 1985: 149.
Aphonopelma anax Smith, 1995: 71.
Aphonopelma breenei Smith, 1995: 78; female holotype from Harlingen, Cameron Co., Texas, 26.190631 -97.6961035, elev. 40ft., 1939, coll. Bryce Brown; deposited in
Diagnosis
Aphonopelma anax (Fig.


Aphonopelma anax (Chamberlin, 1940). A–I male specimen, APH_0924 A dorsal view of carapace, scale bar = 5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 4mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 6mm.



Descriptions
Male and female originally described by
Redescription of male exemplar
(APH_0924; Fig.
Variation (11).Cl 14.371–21.97 (17.885±0.7), Cw 13.32–19.84 (16.557±0.62), LBl 1.96–2.96 (2.341±0.09), LBw 2.16–3.02 (2.711±0.08), F1 14.1–19.3 (16.839±0.55), F1w 3.52–5.1 (4.392±0.15), P1 6.12–8.1 (7.105±0.22), T1 11.32–15.71 (13.314±0.39), M1 9.69–13.43 (11.878±0.4), A1 6.9–9.6 (8.499±0.28), L1 length 48.47–65.97 (57.635±1.73), F3 11.39–16.14 (13.868±0.48), F3w 3.41–5.03 (4.132±0.16), P3 4.86–7.65 (6.035±0.27), T3 7.91–12.16 (10.428±0.43), M3 9.89–14.89 (12.572±0.49), A3 6.93–9.45 (8.291±0.27), L3 length 41.32–59.2 (51.194±1.82), F4 13.52–19.31 (16.56±0.58), F4w 3.22–4.83 (4.094±0.17), P4 5.5–8.16 (6.647±0.26), T4 11.15–15.56 (13.676±0.41), M4 13.66–19.69 (17.13±0.58), A4 8.1–11.15 (9.504±0.3), L4 length 52.79–72.08 (64.171±1.92), PTl 7.187–10.136 (8.709±0.27), PTw 2.246–3.32 (2.882±0.09), SC3 ratio 0.607–0.805 (0.713±0.02), SC4 ratio 0.351–0.524 (0.44±0.02), Coxa 1 setae = thick tapered, F3 condition = normal/slightly swollen.
Redescription of female exemplar
(APH_0857; Figs
Variation (13).Cl 16.06–23.8 (20.27±0.79), Cw 14.9–21.73 (17.951±0.62), LBl 2.45–3.79 (2.878±0.1), LBw 2.71–4.3 (3.478±0.15), F1 13.12–18.14 (15.181±0.45), F1w 3.87–5.8 (4.945±0.18), P1 5.61–8.59 (7.196±0.26), T1 9.76–13.56 (11.616±0.35), M1 7.0–10.98 (8.911±0.31), A1 5.73–8.34 (7.138±0.19), L1 length 42.75–59.3 (50.042±1.47), F3 10.94–14.63 (12.375±0.37), F3w 3.36–5.13 (4.268±0.17), P35.1–7.72 (6.179±0.23), T3 7.45–10.56 (8.816±0.28), M3 8.27–12.05 (9.918±0.34), A3 6.52–8.84 (7.398±0.18), L3 length 38.67–53.7 (44.687±1.26), F4 13.27–18.66 (15.654±0.49), F4w 3.61–5.31 (4.514±0.17), P4 5.34–8.4 (6.55±0.28), T4 9.72–13.98 (12.038±0.35), M4 10.37–16.15 (13.621±0.46), A4 7.01–9.83 (8.373±0.25), L4 length 45.86–66.63 (55.723±1.74), SC3 ratio 0.644–0.763 (0.706±0.01), SC4 ratio 0.368–0.474 (0.433±0.01), Coxa 1 setae = thick tapered. Spermathecae variation can be seen in Figures
Material examined
United States: Texas: Bexar: Hollywood Park, 220 Mecca, 29.59413 -98.479462, 934ft., [APH_0033, 2/6/2006, 1♂, Connor Shannon, Ryan Tubbesing, AUMNH]; Cameron: 2100 W. San Marcelo Blvd #158, Brownsville, 25.95835 -97.5004892, 21ft., [APH_0523, 20/5/2009, 1♂, Lilia Perez, AUMNH]; Brownsville, 25.901747 -97.4974845, 26ft., [APH_2045, 4/1963, 1♀, 1♂, Ted Beimler,
Distribution and natural history
In the United States, A. anax is widely distributed throughout South Texas (Fig.

Conservation status
Aphonopelma anax is very common throughout its distribution. Extensive fieldwork near Edinburg and McAllen (Hidalgo County) suggests that some local populations of A. anax have probably been extirpated due to extensive agriculture in the Lower Rio Grande Valley, but overall the species is fairly abundant throughout South Texas. These spiders appear to thrive in a variety of anthropogenic settings including golf courses, residential lawns, city parks, roadside picnic areas, and mowed highway shoulders. The status of A. anax is likely secure.
Remarks
Aphonopelma anax is one of the largest and most robust Aphonopelma within the United States. This species exhibits size variation within males and females across their distribution, with northern populations generally smaller and the southern populations representing the largest tarantulas in the United States. Other important ratios that distinguish males: A. anax possess a smaller M1/F4 (≤0.75; 0.69-0.75) than A. moderatum (≥0.80; 0.80-0.88) and A. moellendorfi (≥0.81; 0.81-0.88). No other ratios distinguish female A. anax from their syntopic or closely related phylogenetic species. For both males and females, certain morphometrics have potential to be useful though due to the amounts of variation, small number of specimens, and the small differences between species none are claimed to be significant at this time (see Suppl. material
It is important to note the tremendous amount of variation that can be observed in the shape of the spermathecae from numerous populations of A. anax (Figs
Mitochondrial DNA (CO1) identifies A. anax as a polyphyletic group with respect to A. armada and A. hentzi (Fig.
Aphonopelma armada
Dugesiella armada Chamberlin, 1940: 32; female holotype from Austin, Travis Co., Texas, 30.267153 -97.7430616, elev. 461ft., ix.1909, coll. A. Petrunkevitch; deposited in
Rhechostica armada Raven, 1985: 149.
Aphonopelma armada Smith, 1995: 71.
Aphonopelma arnoldi Smith, 1995: 74; male holotype from Hwy 82 near Crosbyton, Crosby Co., Texas, 33.660017 -101.2946445, elev. 3063ft., 17.vi.1963, coll. P. Keathley; deposited in Oklahoma State University collection. [not examined] syn. n.
Diagnosis
Aphonopelma armada (Fig.


Aphonopelma armada (Chamberlin, 1940). A–I male specimen, APH_0950 A dorsal view of carapace, scale bar = 6.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 3mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 2mm.

Description
Female originally described by
Description of male exemplar
(APH_0950; Fig.
Variation (5).Cl 13.19–16.38 (14.614±0.61), Cw 11.37–13.88 (12.912±0.48), LBl 1.79–2.09 (1.956±0.06), LBw 2.04–2.51 (2.28±0.09), F1 13.7–15.67 (14.656±0.4), F1w 3.27–3.97 (3.688±0.13), P1 4.61–6.91 (6.056±0.38), T1 11.1–12.48 (11.724±0.23), M1 9.28–11.36 (10.386±0.39), A1 7.13–8.51 (7.76±0.25), L1 length 46.14–53.64 (50.582±1.41), F3 10.81–12.46 (11.742±0.31), F3w 3.18–3.97 (3.546±0.14), P3 4.99–5.48 (5.236±0.08), T3 8.26–9.35 (8.806±0.22), M3 10.78–12.26 (11.414±0.29), A3 6.68–7.72 (7.25±0.21), L3 length 41.97–46.59 (44.448±1.02), F4 12.85–14.12 (13.4±0.24), F4w 3.17–3.66 (3.474±0.09), P4 4.73–5.96 (5.476±0.22), T4 10.95–13.16 (11.896±0.36), M4 14.18–15.5 (14.95±0.23), A4 7.37–8.77 (8.15±0.24), L4 length 51.3–56.35 (53.872±1.03), PTl 7.053–8.554 (8.061±0.28), PTw 2.372–2.756 (2.578±0.07), SC3 ratio 0.483–0.634 (0.566±0.03), SC4 ratio 0.292–0.371 (0.328±0.01), Coxa I setae = very thick tapered & stout, F3 condition = normal.
Redescription of female exemplar
(APH_0848; Figs

Variation (6).Cl 11.96–16.76 (14.852±0.78), Cw 9.64–14.64 (12.702±0.78), LBl 1.7–2.4 (2.107±0.11), LBw 1.93–2.97 (2.45±0.14), F1 9.847–13.69 (11.846±0.56), F1w 3.31–4.45 (3.908±0.19), P1 4.427–6.51 (5.481±0.33), T1 7.642–10.33 (9.015±0.4), M1 5.741–7.44 (6.824±0.27), A1 4.659–6.84 (5.905±0.3), L1 length 32.316–44.74 (39.071±1.79), F3 7.03–11.18 (9.385±0.63), F3w 2.62–3.72 (3.192±0.19), P3 3.31–5.63 (4.588±0.33), T3 5.51–7.33 (6.422±0.27), M3 5.66–8.73 (7.21±0.41), A3 4.81–6.5 (5.732±0.26), L3 length 26.32–39.37 (33.337±1.84), F4 9.11–12.55 (11.117±0.56), F4w 2.72–4.03 (3.41±0.21), P4 3.85–5.82 (4.963±0.3), T4 7.67–10.35 (9.172±0.43), M4 7.81–11.74 (10.153±0.58), A4 5.58–7.59 (6.528±0.32), L4 length 34.02–47.93 (41.933±2.09), SC3 ratio 0.566–0.737 (0.629±0.02), SC4 ratio 0.282–0.429 (0.356±0.03), Coxa 1 setae = very thick tapered & stout. Spermathecae variation can be seen in Figure
Material examined
United States: Texas: Andrews: SW4001 and SW7000, 32.131369 -102.6216891, 3152ft., [APH_1049, 6/7/2010, 1♀, Skyler Stevens, AUMNH]; SW4001, 32.113981 -102.6158141, 3140ft., [APH_1052, 6/7/2010, 1♂, Skyler Stevens, AUMNH]; Briscoe: Caprock Canyons State Park, Honey Flat camping area (site 4), 34.419514 -101.0560811, 2611ft., [APH_0551-0554, 7/6/2009, 4 juv, Brent E. Hendrixson, Courtney Dugas, Sloan Click, AUMNH]; Burleson: Caldwell, 30.49425 -96.69215, 373ft., [APH_0944, 7/2008, 1♂, Dave Moellendorf, AUMNH]; Coleman: O.H. Ivie reservoir, Coleman Lake at Hords Creek, 31.842967 -99.56961, 1936ft., [APH_0840, 9/2008, 1♀, Chris A. Hamilton, AUMNH]; O.H. Ivie reservoir, 31.57695 -99.660651, 1571ft., [APH_0950, 9/2008, 1♂, Chris A. Hamilton, AUMNH]; Crosby: 0.2 miles N US-82 on FM 2591 (E of Crosbyton), 33.669122 -101.1756561, 2802ft., [APH_0547-0550, 7/6/2009, 4 juv, Brent E. Hendrixson, Courtney Dugas, Sloan Click, AUMNH]; DeWitt: Westhoff, Meyer Rd off Hwy 240 and County Rd 142, 29.136983 -97.4970331, 342ft., [APH_0922-0923, 9/2008, 2♂, Dan Lewis, AUMNH]; Ector: Cowden H Ranch, 32.076633 -102.7899835, 3320ft., [APH_0855, 2006, 1♀, Dave Moellendorf, AUMNH]; [APH_0965-0967, 2006, 3♂, Dave Moellendorf, AUMNH]; [APH_0977, 2006, 1♀, Dave Moellendorf, AUMNH]; Fayette: La Grange, 29.914433 -96.8663171, 321ft., [APH_0807, 5/2008, 1♀, Chris A. Hamilton, AUMNH]; Glasscock: E of Midland, off Hwy 137, 31.94904 -101.723462, 2569ft., [APH_1468, 19/6/2012, 1♂, Darryl Burton, AUMNH]; Hall: Hulver Cemetery, 5.9 miles W US-287 on Hwy-86, 34.522756 -100.5343822, 1955ft., [APH_1460, 27/5/2012, 1♀, Brent E. Hendrixson, AUMNH]; Howard: Big Spring, at miniature golf course, 32.200567 -101.477151, 2701ft., [APH_0841-0844, 9/2008, 4♀, Chris A. Hamilton, AUMNH]; [APH_0945, 9/2008, 1♀, Chris A. Hamilton, AUMNH]; Kimble: Texas Tech Field Station, Junction, 30.472222 -99.7808331, 1718ft., [APH_1067, 24/6/2010, 1♂, Skyler Stevens, AUMNH]; [APH_1068, 16/6/2010, 1♀, Bryce Hubbell, AUMNH]; [APH_1069, 17/6/2010, 1♀, Travis Fisher, AUMNH]; Kinney: 0.59 miles E US-277 on FM-693, 29.17044 -100.6732591, 972ft., [APH_1164, 17/3/2010, 4 juv, Brent E. Hendrixson, Gerri Wilson, Thomas Martin, AUMNH]; Midland: Midland, 31.924217 -102.05831, 2784ft., [APH_0953, 9/2008, 1♀, Chris A. Hamilton, AUMNH]; W County Rd 54, 32.027933 -102.2068531, 2883ft., [APH_1056-1059, 5/7/2010, 4♂, Skyler Stevens, AUMNH]; CR60, 32.010556 -102.22751, 2888ft., [APH_1061-1062, 2/7/2010, 2♂, Skyler Stevens, AUMNH]; near Midland, along Hwy-158, 31.999899 -102.1802161, 2862ft., [APH_1172-1173, 21/7/2010, 1♀, 1♂, Brent E. Hendrixson, Brendon Barnes, Nate Davis, AUMNH]; near Midland, along CR-60, 32.003836 -102.2561841, 2890ft., [APH_1174, 21/7/2010, 1♂, Brent E. Hendrixson, Brendon Barnes, Nate Davis, AUMNH]; 0.5 miles S CR 118W on Hwy-349, 31.58789 -101.97042, 2793ft., [APH_1462, 13/6/2012, 1♂, Darryl Burton, AUMNH]; 0.3 miles S FM-1787 on FM-1492, 31.66265 -102.206892, 2876ft., [APH_1466, 17/6/2012, 1♂, Darryl Burton, AUMNH]; Reagan: Hwy 137, 31.343611 -101.4958331, 2617ft., [APH_1064-1065, 30/6/2010, 2♂, Skyler Stevens, AUMNH]; off Hwy 67, 31.23071389 -101.72644442, 2715ft., [APH_1385, 19/9/2011, 1 juv, Darryl Burton, AUMNH]; Scurry: Snyder, 32.682133 -100.9254831, 2353ft., [APH_0845-0849, 9/2008, 5♀, Chris A. Hamilton, AUMNH]; Tom Green: San Angelo, approx. 1 miles N of W Hwy-67, 31.46431 -100.495114, 1922ft., [APH_0010, 23/6/2005, 1♂, Kati and Martha Mayfield, AUMNH]; Upton: oil fields W of Hwy 329, 31.28048056 -102.08022, 2605ft., [APH_1374, 17/6/2011, 1♀, Darryl Burton, AUMNH]; [APH_1375, 16/6/2011, 1 juv, Darryl Burton, AUMNH]; [APH_1377, 19/7/2011, 1 juv, Darryl Burton, AUMNH]; [APH_1379, 6/9/2011, 1 juv, Darryl Burton, AUMNH]; [APH_1381, 10/9/2011, 1♀, Darryl Burton, AUMNH]; FM 1492, 31.56408333 -102.17425562, 2855ft., [APH_1384, 11/9/2011, 1♀, Darryl Burton, AUMNH]; [APH_1387, 31/8/2011, 1 juv, Darryl Burton, AUMNH]; [APH_1388, 11/9/2011, 1 juv, Darryl Burton, AUMNH]; oil fields E of Hwy 329, 31.35340556 -102.08773332, 2695ft., [APH_1389, 30/8/2011, 1♀, Darryl Burton, AUMNH]; FM 1492, 31.56408333 -102.17425562, 2855ft., [APH_1391, 23/8/2011, 1♀, Darryl Burton, AUMNH]; Val Verde: Del Rio, near River Rd, 29.35553 -100.9722975, 885ft., [APH_0593-0594, Spring 2009, 2 juv, unknown, AUMNH]; 2.2 miles off Hwy 90 on spur 406, N of Del Rio, 29.574484, -101.0418981, 1195ft., [APH_3124, 15/6/2014, 1♀, Dave Moellendorf, AUMNH].
Distribution and natural history
Aphonopelma armada has a wide distribution across Texas and can be found inhabiting these Level III Ecoregions: Chihuahuan Deserts, High Plains, Southwestern Tablelands, Central Great Plains, Edwards Plateau, Southern Texas Plains, Texas Blackland Prairies, and East Central Texas Plains (Fig.

Conservation status
Aphonopelma armada is very common throughout its distribution in South and West Texas. The species is likely secure.
Remarks
The type locality for A. armada is vague (Austin, Texas) and despite much fieldwork in the region, we have never been able to find specimens referable to A. armada in or around the Austin city limits (this area is dominated by A. hentzi). We have, however, found the species in counties to the south and east of Austin and note that it has a crescent-like distribution around Austin and becomes more common to the west (Fig.
We did not examine the holotype of A. arnoldi but we did have the opportunity to study freshly collected topotypic material of the species from Crosbyton, Texas. Our morphological and molecular analyses fail to recognize this species as a separate, independently evolving lineage. As a consequence, we consider A. arnoldi a junior synonym of A. armada.
Aphonopelma atomicum , sp. n.
Aphonopelma mojave Prentice, 1997 (in part): 161.
Types
Male holotype (APH_2727-2) collected at the Nevada Testing Site, Nye Co., Nevada, 37.025579 -116.0238656, elev. 3990ft., viii.1961, coll. Gertsch; deposited in
Etymology
The specific epithet is a neuter noun meaning “of or relating to atoms”. The name references the Nevada Test Site, constructed by the Atomic Energy Commission for the testing of nuclear devices, where this species was originally collected. The name is in homage to the famous sci-fi B movies of the 1950’s, of which Tarantula (1955) was the most entertaining, and slightly ironic given that this species is one of the smallest tarantulas in the United States.
Diagnosis
Aphonopelma atomicum (Fig.

Description of male holotype
(AUMS_2727-2; Fig.

Aphonopelma atomicum sp. n. A–I male holotype, APH_2727-2 A dorsal view of carapace, scale bar = 2mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 3mm E ventral view of metatarsus IV, scale bar = 3mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2.5mm.
Variation (5).Cl 6.523–7.917 (7.157±0.23), Cw 6.382–6.798 (6.64±0.09), LBl 0.832–0.988 (0.927±0.03), LBw 1.098–1.394 (1.231±0.06), F1 7.434–8.185 (7.761±0.12), F1w 1.565–1.914 (1.783±0.08), P1 2.859–3.481 (3.12±0.11), T1 6.618–7.496 (7.053±0.16), M1 5.767–6.402 (6.083±0.12), A1 3.768–4.149 (3.976±0.08), L1 length 26.653–28.926 (27.993±0.5), F3 6.665–7.388 (6.962±0.12), F3w 1.958–2.454 (2.223±0.09), P3 2.355–2.84 (2.562±0.08), T3 5.294–5.812 (5.58±0.1), M3 6.438–7.349 (6.88±0.15), A3 4.204–4.298 (4.252±0.02), L3 length 25.04–27.374 (26.236±0.39), F4 7.773–8.406 (8.073±0.11), F4w 1.596–2.045 (1.816±0.08), P4 2.621–2.9 (2.786±0.06), T4 6.615–7.533 (7.11±0.17), M4 8.197–9.072 (8.573±0.15), A4 4.358–4.937 (4.64±0.1), L4 length 29.958–32.832 (31.181±0.54), PTl 4.486–4.966 (4.689±0.08), PTw 1.433–1.736 (1.564±0.06), SC3 ratio 0.552–0.725 (0.626±0.03), SC4 ratio 0.264–0.409 (0.356±0.03), Coxa I setae = very thin tapered, F3 condition = slightly swollen/swollen.
Description of female paratype
(AUMS_3267-2; Figs


Variation (3).Cl 7.081–7.978 (7.45±0.27), Cw 5.951–7.105 (6.623±0.35), LBl 0.922–1.218 (1.066±0.09), LBw 1.105–1.621 (1.381±0.15), F1 5.656–7.25 (6.465±0.46), F1w 1.663–2.032 (1.862±0.11), P1 2.912–3.253 (3.026±0.11), T1 5.232–5.714 (5.394±0.16), M1 3.461–4.532 (3.982±0.31), A1 2.787–3.179 (3.023±0.12), L1 length 20.052–23.928 (21.889±1.12), F3 5.11–5.984 (5.544±0.25), F3w 1.493–1.907 (1.73±0.12), P3 2.046–2.81 (2.425±0.22), T3 3.603–4.223 (4.005±0.2), M3 3.981–4.61 (4.318±0.18), A3 3.305–3.641 (3.455±0.1), L3 length 18.045–21.268 (19.746±0.93), F4 6.234–7.413 (6.848±0.34), F4w 1.473–1.956 (1.732±0.14), P4 2.541–2.92 (2.738±0.11), T4 5.234–6.039 (5.663±0.23), M4 5.687–6.245 (5.975±0.16), A4 3.446–3.86 (3.715±0.13), L4 length 23.142–26.477 (24.939±0.97), SC3 ratio 0.683–0.755 (0.726±0.02), SC4 ratio 0.372–0.49 (0.439±0.03), Coxa I setae = thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: California: Inyo: Death Valley National Park, Hwy-178, Salsberry Pass, 35.923118 -116.4317471, 3204ft., [APH_1478, 30/7/2012, 1♀, Brent E. Hendrixson, Brendon Barnes, Austin Deskewies, AUMNH]; Death Valley National Park, ~7.5 miles S Hwy-190 on Dante’s View Rd, 36.267976 -116.6655761, 3242ft., [APH_1479, 30/7/2012, 1 juv, Brent E. Hendrixson, Brendon Barnes, Austin Deskewies, AUMNH]; Nevada: Nye: Rock Valley Wash, off Hwy-95, 36.622963 -116.3094641, 2784ft., [APH_1549, 24/10/2012, 1♀, Brent E. Hendrixson, AUMNH]; Rock Valley, 36.632732 -116.3139335, 2884ft., [AUMS_2637, 20/10/1972, 1♀, E.L. Sleeper, AUMNH]; Mercury, 36.66051 -115.9944755, 3796ft., [APH_2725-1, 6/9/1960, 2♀, 14♂, Gertsch,
Distribution and natural history
Aphonopelma atomicum is only known from a handful of specimens surrounding the Amargosa Desert in southern Nye County (Nevada) and southeastern Inyo County (California), including the Amargosa Range and Nevada Test Site (Fig.

Conservation status
Aphonopelma atomicum has a highly restricted distribution limited to the mountains and foothills surrounding the Amargosa Desert and Death Valley. While this species is not dramatically different from A. prenticei, it is genetically unique and should be considered important. The species is most likely secure.
Remarks
Aphonopelma atomicum is unique because it was quite possibly the first miniature tarantula species collected in the United States (although it was never described and sat on a shelf in the
Aphonopelma catalina , sp. n.
Types
Male holotype (APH_1440) from Coronado National Forest, along Bug Spring Trail, Pima Co., Arizona, 32.34544 -110.716024, elev. 5255ft., 17.xii.2011, coll. Brent E. Hendrixson and Thomas Martin; deposited in AUMNH. Paratype female (APH_1602) from Coronado National Forest, along Bug Spring Trail, Pima Co., Arizona, 32.34544 -110.716024, elev. 5255ft., 9.xi.2012, coll. Brent E. Hendrixson; deposited in AUMNH. Paratype male (APH_1439) from Coronado National Forest, along Bug Spring Trail, Pima Co., Arizona, 32.34544 -110.716024, elev. 5255ft., 17.xii.2011, coll. Brent E. Hendrixson and Thomas Martin; deposited in
Etymology
The specific epithet is a noun in apposition taken from type locality, the Santa Catalina Mountains near Tucson, Arizona, where this new species appears to be endemic.
Diagnosis
Aphonopelma catalina (Fig.

Description of male holotype
(APH_1440; Fig.

Aphonopelma catalina sp. n. A–I male holotype, APH_1440 A dorsal view of carapace, scale bar = 4mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 3mm E ventral view of metatarsus IV, scale bar = 4mm F prolateral view of L pedipalp and palpal tibia, scale bar = 4mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 4.5mm.
Variation (4).Cl 9.57–12.39 (10.595±0.62), Cw 8.88–11.33 (9.708±0.55), LBl 0.967–1.237 (1.114±0.06), LBw 1.102–1.56 (1.354±0.11), F1 11.28–14.64 (12.765±0.7), F1w 2.18–2.83 (2.467±0.14), P1 3.73–5.31 (4.483±0.32), T1 9.89–12.78 (11.132±0.61), M1 6.42–8.91 (7.446±0.54), A1 5.0–5.95 (5.392±0.2), L1 length 36.32–47.59 (41.217±2.34), F3 7.97–10.4 (9.098±0.5), F3w 2.1–2.88 (2.465±0.16), P3 3.04–4.084 (3.551±0.26), T3 5.74–8.63 (7.039±0.6), M3 6.72–9.18 (7.882±0.51), A3 4.82–5.89 (5.37±0.23), L3 length 28.29–37.99 (32.94±2.02), F4 9.75–12.9 (11.077±0.66), F4w 2.05–2.78 (2.357±0.15), P4 3.26–4.54 (3.902±0.26), T4 7.82–10.96 (9.444±0.64), M4 9.26–12.34 (10.762±0.63), A4 5.41–7.11 (6.095±0.4), L4 length 35.5–47.85 (41.279±2.53), PTl 6.587–8.058 (7.173±0.32), PTw 2.328–2.911 (2.572±0.12), SC3 ratio 0.481–0.53 (0.514±0.01), SC4 ratio 0.228–0.359 (0.286±0.03), Coxa I setae = tapered/thin tapered, F3 condition = normal.
Description of female paratype
(APH_1602; Figs


Variation (2).Cl 14.79–16.39 (15.59±0.8), Cw 13.74–15.06 (14.4±0.66), LBl 1.81–2.03 (1.92±0.11), LBw 1.97–2.67 (2.32±0.35), F1 11.87–13.85 (12.86±0.99), F1w 3.84–4.24 (4.04±0.2), P1 5.36–5.58 (5.47±0.11), T1 9.77–11.33 (10.55±0.78), M1 6.92–7.57 (7.245±0.33), A1 6.04–6.05 (6.045±0), L1 length 39.96–44.38 (42.17±2.21), F3 9.74–11.31 (10.525±0.79), F3w 3.32–3.78 (3.55±0.23), P3 4.52–5.33 (4.925±0.41), T3 7.26–8.35 (7.805±0.55), M3 7.29–8.42 (7.855±0.57), A3 6.07–6.77 (6.42±0.35), L3 length 34.88–40.18 (37.53±2.65), F4 12.52–13.95 (13.235±0.72), F4w 3.41–3.86 (3.635±0.23), P4 4.92–5.54 (5.23±0.31), T4 10.0–11.17 (10.585±0.59), M4 10.98–11.74 (11.36±0.38), A4 6.64–7.84 (7.24±0.6), L4 length 45.06–50.24 (47.65±2.59), SC3 ratio 0.629–0.686 (0.657±0.03), SC4 ratio 0.37–0.465 (0.418±0.05), Coxa I setae = medium tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Pima: Coronado National Forest, along Bug Spring Trail, 32.34544 -110.716024, 5255ft., [APH_0454, 28/12/2008, 1♂, Paul E. Marek, Charity Hall, AUMNH]; [APH_1438, 11/12/2011, 1♂, Jillian Cowles, Bill Savary, AUMNH]; [APH_1439-1440, 17/12/2011, 2♂, Brent E. Hendrixson, Thomas Martin, AUMNH &
Distribution and natural history
Aphonopelma catalina is known from only six individuals collected within a few kilometers of each other but this species appears to be a sky island endemic found in the Santa Catalina Mountains in Pima County, Arizona at elevations above 1480 meters in oak-grassland communities (Figs

Conservation status
It is difficult to fully assess the distribution and abundance (and therefore the conservation status) of A. catalina due to a lack of specimens and thorough sampling; however, as previously mentioned, the species appears to be narrowly endemic to the Santa Catalina Mountains, which may put the species at some risk. This mountain range is entirely contained within the Coronado National Forest (Santa Catalina Ranger District) which is afforded some degree of protection; however, increased urbanization of the Tucson Metropolitan Area (one of the most rapidly growing areas in the United States), increased recreation in the mountains, and climate change have impacted these habitats (
Remarks
As noted in
Aphonopelma chalcodes
Aphonopelma chalcodes Chamberlin, 1940: 7; male holotype, male paratype, and two female paratypes from Tucson, Pima Co., Arizona, 32.221743 -110.9264796, elev. 2473ft., 27.vii.1936, coll. Prof. C.T. Vorhies; deposited in
Rhechostica chalcodes Raven, 1985: 149.
Aphonopelma chalcodes Smith, 1995: 82.
Aphonopelma apacheum Chamberlin, 1940: 15; male holotype from Tucson, Pima Co., Arizona, 32.221743 -110.9264796, elev. 2473ft., elev. ft., no collecting date, coll. unknown; deposited in
Rhechostica apacheum Raven, 1985: 149.
Aphonopelma apacheum Smith, 1995: 73. syn. n.
Aphonopelma minchi Smith, 1995: 121; male holotype from Usery Pass Rd., near Usery Mountain Regional Park, Maricopa Co., Arizona, 33.482543 -111.6231784, elev. 2033ft., no collecting date, coll. A. Smith and M. Sullivan; deposited in BMNH. Paratype male from western end of Apache Trail, 33.444378 -111.5104915, elev. 1937ft., no collecting date, coll. A. Smith and M. Sullivan; deposited in BMNH. [examined] syn. n.
Aphonopelma schmidti Smith, 1995: 140; male holotype and female paratype from Mineral Mountain, near Florence Junction on Hwy 60, Pinal Co., Arizona, 33.265044 -111.3484275, elev. 1916ft., 10.viii.1992, coll. A. Smith and M. Sullivan; deposited in BMNH. [examined] syn. n.
Aphonopelma stahnkei
Diagnosis
Aphonopelma chalcodes (Fig.

Redescription of male exemplar
(APH_0954; Fig.

Aphonopelma chalcodes Chamberlin, 1940. A–I male specimen, APH_0954 A dorsal view of carapace, scale bar = 5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 3.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1.5mm I prolateral view of tibia I (mating clasper), scale bar = 4.5mm.
Variation (13).Cl 14.43–21.07 (16.98±0.51), Cw 13.23–18.32 (15.675±0.45), LBl 1.44–2.6 (2.07±0.08), LBw 1.69–2.91 (2.401±0.08), F1 14.56–19.13 (16.687±0.38), F1w 3.65–5.05 (4.119±0.12), P1 6.0–8.11 (6.852±0.16), T1 13.29–15.65 (14.136±0.19), M1 12.08–15.77 (13.313±0.3), A1 7.49–9.02 (8.219±0.14), L1 length 53.99–67.39 (59.208±1.09), F3 12.86–15.58 (14.193±0.25), F3w 3.97–5.5 (4.369±0.13), P3 5.08–6.86 (5.769±0.15), T3 10.26–12.33 (11.244±0.21), M3 12.46–14.84 (13.615±0.24), A3 7.16–9.09 (8.096±0.15), L3 length 48.04–58.45 (52.917±0.92), F4 15.25–18.59 (16.683±0.3), F4w 3.53–5.09 (4.012±0.12), P4 5.09–7.09 (6.138±0.15), T4 12.11–15.53 (13.848±0.28), M4 14.9–20.1 (17.587±0.39), A4 8.03–10.05 (8.951±0.17), L4 length 56.11–70.85 (63.208±1.17), PTl 8.835–11.026 (9.601±0.19), PTw 2.79–3.48 (3.065±0.06), SC3 ratio 0.651–0.86 (0.773±0.02), SC4 ratio 0.428–0.764 (0.647±0.03), Coxa I setae = tapered, F3 condition = normal/slightly swollen.
Redescription of female exemplar
(APH_0887; Figs

Aphonopelma chalcodes Chamberlin, 1940. A–E female specimen, APH_0887 A dorsal view of carapace, scale bar = 8mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 4.5mm D ventral view of metatarsus IV, scale bar = 5mm E prolateral view of L pedipalp and palpal tibia.


Variation (10).Cl 13.49-21.79 (18.548±0.72), Cw 11.72-19.71 (16.589±0.68), LBl 1.90-2.61 (2.375±0.07), LBw 2.21-3.02 (2.773±0.1), F1 11.39-16.37 (14.684±0.42), F1w 3.34–4.76 (4.397±0.14), P1 5.03–7.26 (6.554±0.21), T1 9.03–12.42 (11.224±0.31), M1 6.81–11.81 (9.35±0.41), A1 5.68–7.39 (7.012±0.16), L1 length 37.94–55.02 (48.824±1.42), F3 9.38–13.96 (12.093±0.38), F3w 3.15–4.58 (4.028±0.13), P3 3.92–6.25 (5.575±0.21), T3 6.78–9.67 (8.581±0.25), M3 7.83–11.6 (9.925±0.34), A3 5.89–7.32 (7.035±0.14), L3 length 33.8–48.7 (43.209±1.2), F4 12.15–16.3 (14.699±0.36), F4w 3.11–4.73 (4.134±0.14), P4 4.25–6.98 (6.134±0.24), T4 9.59–12.45 (11.334±0.25), M4 11.16–15.35 (13.498±0.37), A4 6.51–8.28 (7.676±0.15), L4 length 43.66–57.92 (53.341±1.24), SC3 ratio 0.728–0.926 (0.843±0.02), SC4 ratio 0.566–0.812 (0.682±0.03), Coxa I setae = medium tapered. Spermathecae variation can be seen in Figures
Material examined
United States: Arizona: Cochise: 0.6 miles E of Portal, 31.913718 -109.1300894, 4700ft., [AUMS_2676, 28/3/1990, 1♂, T.R. Prentice, AUMNH]; 1.4 miles NW Portal Rd on FR 42B (San Simon Rd); 31.926949 -109.1691181, 5126ft., [APH_1229, 7/8/10, 1♂, Brent E. Hendrixson, Ashley Bailey, Andrea Reed, AUMNH]; 10 miles east of Dos Cabezas, 31.93051 -109.7947535, 4281ft., [APH_2062, 4/8/71, 1♂, A. Jung,
Distribution and natural history
Aphonopelma chalcodes is widely distributed across the southern two-thirds of Arizona south of the Grand Canyon, bound to the west by the Colorado River and barely making its way into southwestern New Mexico (Fig.

Conservation status
Aphonopelma chalcodes is the most widespread and abundant tarantula species in Arizona. The species is secure.
Remarks
Aphonopelma chalcodes is herein considered a member of the problematic Iodius species group. Morphological and molecular data confirm that A. chalcodes is the sister lineage to the remaining species in the group (A. iodius, A. eutylenum, and A. johnnycashi). There are no major morphological features that can be used to distinguish A. chalcodes from these species so we must rely on molecular data and distributional information (A. chalcodes is largely restricted to Arizona south of the Grand Canyon). Other important ratios that distinguish males: A. chalcodes possess a larger M1/A1 (≥1.51; 1.51–1.79) than A. johnnycashi (≤1.43; 1.29–1.43). Other important ratios that distinguish females: A. chalcodes possess a smaller F1/M1 (≤1.68; 1.38–1.68) than A. catalina (≥1.71; 1.71–1.83), A. chiricahua (1.84 ± (only 1 specimen)), A. madera (≥1.73; 1.73; 1.73–2.15), and A. marxi (≥1.77; 1.77–1.88); by possessing a smaller P1/M1 (≤0.75; 0.61–0.75) than A. vorhiesi (≥0.75; 0.75–0.85). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Mitochondrial DNA (CO1) identifies A. chalcodes as a polyphyletic group with some samples more closely related to specimens of A. iodius (Fig.
Aphonopelma chiricahua , sp. n.
Types
Male holotype (APH_3191) collected 1 mile up the road (42 Forest Rd.) from the lookout trail, Cochise Co., Arizona, 31.886417 -109.1733561, elev. 5083ft., 14.xi.2013, coll. Helen Snyder; deposited in AUMNH. Paratype female (APH_2097) from SWRS (Southwest Research Station, 5 miles W of Portal), Cochise Co., Arizona, 31.884056 -109.2082615, elev. 5436ft., 30.xi.1965, coll. Jon Jenson; deposited in
Etymology
The specific epithet is a noun in apposition taken from type locality, the Chiricahua Mountains outside of Portal, Arizona, where this new species appears to be endemic.
Diagnosis
Aphonopelma chiricahua (Fig.

Description of male holotype
(APH_3191; Fig.

Aphonopelma chiricahua sp. n. A–I male holotype, APH_3191 A dorsal view of carapace, scale bar = 5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 3mm E ventral view of metatarsus IV, scale bar = 4mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 5mm.
Variation (7).Cl 6.837–11.42 (8.18±0.62), Cw 6.254–11.22 (8.269±0.86), LBl 0.684–1.368 (0.959±0.11), LBw 0.985–1.765 (1.292±0.11), F1 6.145–12.718 (8.731±0.77), F1w 1.898–3.281 (2.309±0.19), P1 2.859–4.947 (3.517±0.27), T1 5.851–11.372 (7.397±0.7), M1 4.09–7.61 (5.06±0.46), A1 3.572–6.165 (4.542±0.31), L1 length 22.568–42.812 (29.248±2.48), F3 5.591–9.531 (6.823±0.5), F3w 1.688–2.982 (2.147±0.18), P3 2.304–4.112 (2.896±0.23), T3 4.162–7.603 (5.286±0.43), M3 4.379–7.794 (5.317±0.45), A3 3.955–6.838 (5.003±0.35), L3 length 20.391–35.878 (25.325±1.95), F4 6.648–11.414 (8.181±0.62), F4w 1.74–3.205 (2.174±0.2), P4 2.524–4.414 (3.141±0.25), T4 5.784–9.674 (7.104±0.48), M4 5.772–10.277 (7.342±0.56), A4 4.944–7.78 (5.781±0.38), L4 length 25.672–43.559 (31.549±2.26), PTl 4.42–7.341 (5.424±0.36), PTw 1.888–2.82 (2.241±0.12), SC3 ratio 0.48–0.656 (0.556±0.02), SC4 ratio 0.33–0.404 (0.376±0.01), Coxa I setae = thin/very thin tapered, F3 condition = normal.
Description of female paratype
(APH_2097; Fig.

Aphonopelma chiricahua sp. n. A–F female paratype, APH_2097 A dorsal view of carapace, scale bar = 3.5mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 2mm D ventral view of metatarsus IV, scale bar = 2mm E prolateral view of L pedipalp and palpal tibia F cleared spermathecae.
Material examined
United States: Arizona: Cochise: SWRS (5 miles W of Portal), 31.884056 -109.2082615, 5436ft., [APH_2097, 30/11/1965, 1♀, Jon Jenson,
Distribution and natural history
Aphonopelma chiricahua is a sky island endemic restricted to the Chiricahua Mountains in Cochise County, Arizona at elevations ranging from 1550 to 2700 meters in oak woodland, pine-oak woodland, and mixed conifer communities (Fig.

Conservation status
It is difficult to assess the conservation status of Aphonopelma chiricahua due to small sample sizes and the very cryptic nature of these spiders. This species does not occur outside of the Chiricahua Mountains so its narrow distribution is one factor that may threaten its future survival. These mountains have the advantage of being somewhat protected by their remoteness and management by the federal government (Coronado National Forest, Douglas Ranger District, Chiricahua National Monument); however, these habitats have also been subjected to habitat degradation from recent urban growth, human-caused forest fires, off-road driving, poorly managed livestock grazing, invasive species, recreational activities, human immigrants, and illegal drug trafficking (
Remarks
Aphonopelma chiricahua is morphologically very similar to other Madrean sky island endemics in the Marxi species group, although generally smaller than A. catalina, A. madera, and A. vorhiesi. Other important ratios that distinguish males: A. chiricahua possess a larger Cl/M3 (≥1.46; 1.46–1.61) than A. catalina (≤1.42; 1.26–1.42), A. parvum (≤1.39; 1.20–1.39), A. peloncillo (≤1.40; 1.20–1.40), and A. vorhiesi (≤1.43; 1.24–1.43); by possessing a larger L3/Cl (≥2.98; 2.98–3.19) than A. madera (≤2.95; 2.71–2.95). Other important ratios that distinguish females: A. chiricahua possess a smaller M3/P4 (1.02 ± (only 1 specimen)) than A. catalina (≥1.48; 1.48–1.52), A. madera (≥1.39; 1.39–1.48), A. parvum (≥1.32; 1.32–1.54), A. peloncillo (≥1.39; 1.39–1.67), and A. vorhiesi (≥1.27; 1.27–1.64). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
This species was first identified as novel by
Aphonopelma eutylenum
Aphonopelma eutylenum Chamberlin, 1940: 9; male holotype and female allotype from San Diego, San Diego Co., California, 32.715738 -117.1610856, elev. 54ft., 1935, coll. unknown; deposited in
Rhechostica eutylenum Raven, 1985: 149.
Aphonopelma eutylenum Smith, 1995: 99.
Aphonopelma chambersi Smith, 1995: 86; male holotype from Garner Valley, S of Idyllwild-Pine Cove, Riverside Co., California, 33.635459 -116.6439745, elev. 4475ft., no collecting date, coll. Aaron Chambers; deposited in BMNH. [examined] syn. n.
Aphonopelma clarum Chamberlin, 1940: 10; male holotype from mountains near Claremont, Los Angeles Co., California, 34.137812 -117.7181944, elev. 1571ft., no collecting date, coll. R.V. Chamberlin; deposited in
Rhechostica clarum Raven, 1985: 149.
Aphonopelma clarum Smith, 1995: 89. syn. n.
Aphonopelma cryptethus Chamberlin, 1940: 16; male holotype from Los Angeles, Los Angeles Co., California, 34.052234 -118.2436856, elev. 309ft., 9.v.1908, coll. unknown; deposited in
Rhechostica cryptethus Raven, 1985: 149.
Aphonopelma cryptethum Smith, 1995: 95. syn. n.
Aphonopelma sandersoni Smith, 1995: 138; male holotype and female allotype from San Bernardino Mountains., San Bernardino Co., California, 34.180742 -117.1646387, elev. 3137ft., x.1995, coll. R. Douglas; deposited in BMNH. [examined] syn. n.
Aphonopelma sullivani Smith, 1995: 149; male holotype from Coachella Valley, Palm Springs, Riverside Co., California, 33.767209 -116.3598686, elev. 277ft., ix.1991, coll. Michael Sullivan; deposited in BMNH. [examined] syn. n.
Diagnosis
Aphonopelma eutylenum (Fig.

Description
Originally described by
Redescription of male exemplar
(APH_1088; Fig.

Aphonopelma eutylenum Chamberlin, 1940. A–I male specimen, APH_1088 A dorsal view of carapace, scale bar = 6mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 5mm E ventral view of metatarsus IV, scale bar = 4.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 6mm.
Variation (8).Cl 12.01–18.42 (15.497±0.73), Cw 10.92–16.38 (14.213±0.68), LBl 1.53–2.14 (1.846±0.09), LBw 1.57–2.38 (2.113±0.11), F1 12.67–16.86 (15.34±0.5), F1w 2.85–4.51 (3.799±0.2), P1 4.94–7.22 (6.028±0.28), T1 10.52–13.62 (12.456±0.42), M1 10.08–14.57 (12.415±0.59), A1 6.71–9.18 (8.328±0.32), L1 length 45.83–61.45 (55.503±1.95), F3 10.75–14.19 (12.924±0.45), F3w 3.21–4.77 (3.944±0.19), P3 3.98–5.98 (5.186±0.28), T3 8.82–11.26 (10.247±0.36), M3 10.59–14.7 (13.179±0.67), A3 6.75–8.48 (7.771±0.26), L3 length 41.73–54.29 (49.353±2.08), F4 12.93–16.82 (15.241±0.54), F4w 2.85–4.41 (3.656±0.19), P4 4.61–6.71 (5.734±0.26), T4 10.79–14.21 (12.781±0.53), M4 13.84–18.71 (16.776±0.77), A4 7.42–9.47 (8.556±0.31), L4 length 49.65–65.06 (59.139±2.5), PTl 7.375–10.727 (8.984±0.35), PTw 2.319–3.47 (3.082±0.14), SC3 ratio 0.728–0.93 (0.827±0.02), SC4 ratio 0.626–0.772 (0.701±0.02), Coxa I setae = tapered, F3 condition = slightly swollen.
Description of female exemplar
(APH_1031; Figs

Aphonopelma eutylenum Chamberlin, 1940. A–E female specimen, APH_1031. A dorsal view of carapace, scale bar = 8mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 4.5mm D ventral view of metatarsus IV, scale bar = 5mm E prolateral view of L pedipalp and palpal tibia.

Variation (8).Cl 15.23–20.84 (18.248±0.72), Cw 13.33–17.78 (16.206±0.61), LBl 1.95–2.81 (2.353±0.12), LBw 2.28–3.04 (2.675±0.1), F1 12.87–15.78 (14.599±0.5), F1w 3.62–4.94 (4.357±0.2), P1 5.38–7.34 (6.479±0.25), T1 9.82–13.05 (11.243±0.42), M1 8.36–10.75 (9.565±0.33), A1 6.6–7.92 (7.36±0.17), L1 length 43.43–54.02 (49.137±1.72), F3 10.53–13.35 (12.208±0.42), F3w 3.3–4.67 (4.044±0.16), P3 5.12–7.26 (5.776±0.27), T3 8.3–9.93 (8.82±0.2), M3 8.64–11.06 (9.958±0.36), A3 6.49–7.87 (7.084±0.16), L3 length 39.24–48.1 (43.845±1.2), F4 13.21–16.57 (14.909±0.53), F4w 3.42–4.73 (4.106±0.19), P4 5.51–7.40 (6.312±0.32), T4 10.5–12.84 (11.509±0.31), M4 11.84–15.26 (13.32±0.48), A4 6.94–8.82 (7.779±0.26), L4 length 48.62–60.13 (54.192±1.91), SC3 ratio 0.772–0.906 (0.851±0.02), SC4 ratio 0.628–0.747 (0.683±0.02), Coxa I setae = tapered. Spermathecae variation can be seen in Figure
Material examined
United States: California: Imperial: 1.5 miles N Gordons Well off of I-8, 32.7636 -114.9664312, 248ft., [APH_0152, unknown, 1 juv, T.R. Prentice, AUMNH]; E of Brawley, off of Hwy 78, 32.969167 -115.2594445, 3ft., [APH_3116, 10/1992, 1♀, T.R. Prentice, AUMNH]; East Mesa, 33.058096 -115.3241475, 46ft., [AUMS_3332, 30/5/1995, 1♂, unknown, AUMNH]; East Mesa Exit of I-8, 2.1 miles N of I-8, 32.73013 -114.9553514, 158ft., [AUMS_2358, 26/10/1993, 1♂, Greg Ballmer, AUMNH]; East Mesa, 1/2 mile E of Highline Canal and 3/10 mile S of Hwy 78, 32.968133 -115.2907171, 37ft., [APH_3211, 15/11/2013, 1♂, W. Icenogle, AUMNH]; East Mesa, 5.4 miles E of Hwy 115 E of Wander Linder Exit of I-8, 32.788365 -115.2074794, 50ft., [AUMS_3303, 30/5/1995, 1♀, unknown, AUMNH]; East Mesa, 5.4 miles E of Hwy 115 E of Wander Linder Exit, 2.1 miles N, 32.790359 -115.2405694, 63ft., [AUMS_3341, 30/5/1995, 1♂, T.R. Prentice, AUMNH]; Hwy 78, 1.0 miles east of Highline Canal, 32.970575 -115.283744, 30ft., [AUMS_2356,-2357 3/11/1998, 2♂, Greg Ballmer, AUMNH]; Hwy 78, 3.35 miles E of East Highline Canal, N of 78 ~0.4 miles, 32.972797 -115.2426654, 90ft., [AUMS_2674, unknown, 1♀, T.R. Prentice, AUMNH]; off I-8 split, W of Devils Canyon, W of Ocotillo, 32.67371 -116.101471, 2238ft., [APH_1028-1029, 18/5/2010, 1♂, 1♀, Chris A. Hamilton, Xavier Atkinson, AUMNH]; Pinto Wash, 32.708139 -115.7155255, 13ft., [APH_2690, 5/3/1961, 1♂, Laurie Breese,
Distribution and natural history
Aphonopelma eutylenum has a distribution that stretches from the western part of the Transverse Ranges, south down the length of the California coast along the Peninsular Ranges, and west of the Mojave Desert (Fig.

Conservation status
Aphonopelma eutylenum is widely distributed across Southern California and is very common. The species is likely secure although some localized populations in urbanized areas (e.g., Los Angeles and San Diego) are likely threatened by human encroachment and development.
Remarks
Aphonopelma eutylenum can easily be differentiated from A. steindachneri and A. xwalxwal by the extent of scopulation on legs III and IV, and from A. xwalxwal a larger body size. Female and immature male A. eutylenum can be distinguished from A. steindachneri and A. xwalxwal by body color as well. Other important ratios that distinguish males: A. eutylenum possess a smaller T1/F3 (≤1.00; 0.93–1.00) than A. steindachneri (≥1.01; 1.01–1.11) and A. xwalxwal (≥1.10; 1.10–1.17); by possessing a larger T3/A3 (≥1.30; 1.30–1.35) than A. johnnycashi (≤1.27; 1.18–1.27), but smaller than A. xwalxwal (≥1.41; 1.41–1.64). Other important ratios that distinguish females: A. eutylenum possess a larger M1/M4 (≥0.67; 0.67–0.78) than A. steindachneri (≤0.67; 0.62–0.67). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Mitochondrial DNA (CO1) is problematic in the Iodius species group. While this locus identifies A. eutylenum as a monophyletic group, sister relationships are unclear. Nuclear DNA reveals the true evolutionary history of the A. eutylenum lineage and highlights the ineffectiveness of CO1 for accurately delimiting species boundaries within this group.
Aphonopelma gabeli
Aphonopelma gabeli Smith, 1995: 100; male holotype from E of Tucson, Pima Co., Arizona, 31.956396 -110.3398177, elev. 4055ft., no collecting date, coll. Russ Gurley; deposited in BMNH. [examined]
Diagnosis
Aphonopelma gabeli (Fig.

Description
Male originally described by
Redescription of male exemplar
(APH_1054; Fig.

Aphonopelma gabeli Smith, 1995. A–I male specimen, APH_1054 A dorsal view of carapace, scale bar = 5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4.5mm E ventral view of metatarsus IV, scale bar = 4mm F prolateral view of L pedipalp and palpal tibia, scale bar = 5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 5mm.
Variation (6).Cl 15.21–16.79 (15.667±0.24), Cw 13.4–15.08 (14.075±0.25), LBl 1.7–2.02 (1.872±0.06), LBw 1.91–2.27 (2.075±0.05), F1 15.63–17.21 (16.127±0.23), F1w 3.41–3.65 (3.518±0.04), P1 5.53–6.37 (6.022±0.13), T1 12.7–13.55 (13.067±0.12), M1 11.71–12.98 (12.328±0.18), A1 7.64–8.56 (7.953±0.14), L1 length 54.27–57.46 (55.497±0.44), F3 13.4–14.24 (13.665±0.13), F3w 3.36–4.01 (3.723±0.09), P3 4.67–5.53 (5.253±0.13), T3 10.02–10.89 (10.472±0.13), M3 12.86–13.88 (13.28±0.14), A3 7.33–7.91 (7.635±0.1), L3 length 48.39–51.87 (50.305±0.46), F4 14.81–17.47 (15.853±0.38), F4w 3.44–3.81 (3.592±0.06), P4 4.79–6.02 (5.423±0.2), T4 12.64–13.39 (12.985±0.12), M4 16.33–18.26 (17.022±0.28), A4 8.09–9.14 (8.673±0.15), L4 length 57.18–63.65 (59.957±0.88), PTl 7.677–8.48 (8.083±0.11), PTw 2.38–2.59 (2.525±0.03), SC3 ratio 0.591–0.721 (0.663±0.02), SC4 ratio 0.361–0.471 (0.416±0.02), Coxa I setae = tapered, F3 condition = normal.
Description of female exemplar
(APH_0680; Figs


Variation (6).Cl 14.69–18.34 (16.535±0.53), Cw 12.65–15.81 (14.145±0.5), LBl 2.07–2.41 (2.29±0.06), LBw 2.29–2.82 (2.485±0.08), F1 12.13–15.24 (13.288±0.45), F1w 3.67–4.26 (3.917±0.09), P1 5.41–6.26 (5.772±0.15), T1 9.35–12.11 (10.285±0.4), M1 7.41–10.2 (8.33±0.43), A1 5.83–7.57 (6.647±0.24), L1 length 40.18–51.38 (44.322±1.6), F3 9.63–12.72 (10.783±0.44), F3w 3.44–3.76 (3.575±0.05), P3 4.27–6.26 (5.053±0.28), T3 7.08–8.88 (7.773±0.26), M3 7.78–10.71 (8.75±0.41), A3 6.09–7.73 (6.593±0.24), L3 length 35.81–46.30 (38.953±1.56), F4 12.21–14.90 (13.065±0.41), F4w 3.43–3.83 (3.642±0.06), P4 4.67–6.55 (5.553±0.27), T4 9.31–12.03 (10.187±0.4), M4 10.5–13.75 (11.663±0.48), A4 6.71–8.49 (7.168±0.28), L4 length 43.83–55.72 (47.637±1.7), SC3 ratio 0.725–0.805 (0.762±0.01), SC4 ratio 0.397–0.529 (0.471±0.02), Coxa 1 setae = tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Cochise: 0.1 mi. west of Portal, 31.913699 -109.1431844, 4780ft., [APH_2356, 2/7/1961, 1♂, J. Cole,
Distribution and natural history
Aphonopelma gabeli is distributed mostly throughout the Chihuahuan Desert in southeastern Arizona, southern New Mexico, and West Texas; this includes the northern finger-like extensions of the desert along the Rio Grande and Pecos Rivers into Socorro and Chaves Counties, New Mexico, respectively (Fig.

Conservation status
Landscape fragmentation due to oil and natural gas production has raised concern about the conservation status of some species in the Permian Basin of West Texas and southeastern New Mexico (
Remarks
Aphonopelma gabeli females and juvenile males are easily differentiated from A. armada by the flared metatarsal scopulae and prolateral coxa I setae of A. armada; from A. anax by spermathecae and palpal bulbs, as well as prolateral coxa I setae; from A. hentzi by its phenotypic appearance and prolateral coxa I setae; from A. moderatum by their unique phenotypic color and banding; from A. peloncillo by its phenotypic appearance; from A. vorhiesi by their black appearance; and from A. parvum due to the extreme small size of the miniature species. Other important ratios that distinguish males: A. gabeli possess a larger F4L/W (≥4.29; 4.29–4.60) than A. hentzi (≤4.24; 3.62–4.24); by possessing a smaller L1/L4 (≤0.95; 0.91–0.95) than A. moderatum (>0.95; 0.95–0.99) and A. moellendorfi (≥0.96; 0.96–1.00); by possessing a larger L4 scopulation extent (36%–47%) than A. vorhiesi (20%-36%) and smaller than A. chalcodes (66%–76%); by possessing a smaller F1/M3 (≤1.24; 1.18–1.24) than A. anax (≥1.28; 1.28–1.43), A. armada (≥1.26; 1.26–1.33), and A. peloncillo (≥1.33; 1.33–1.49). Other important ratios distinguish females: A. gabeli possess a larger T3/T4 (≥0.73; 0.73–0.79) than A. armada (≤0.73; 0.64–0.73) and A. moderatum (≤0.71; 0.63–0.71); by possessing a larger L3 scopulation extent (72%-80%) than A. peloncillo (58%–68%) and A. vorhiesi (49%–69%); by possessing a larger CL/CW (≥1.14; 1.14–1.20) than A. chalcodes (≤1.14; 1.09–1.14). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Aphonopelma hentzi
Mygale hentzii Girard, 1852: 251; male holotype from southwestern Oklahoma, 34.309094 -98.3964947, elev. 968ft., coll. unknown, 1849-1852.
Eurypelma hentzi Simon, 1891: 322.
Rhechostica hentzi Raven, 1985: 149.
Aphonopelma hentzi Smith, 1995: 107; neotype male and female exemplar from Garfield Co., Oklahoma, 36.436139 -97.8721607, elev. 1264ft., summer 1975, coll. R.L. Lardie; deposited in Oklahoma State University collection. [not examined]
Aphonopelma clarki Smith, 1995: 87; female holotype from Dallas, Dallas Co., Texas, 32.780141 -96.8004517, elev. 421ft., 1968, coll. H.J. Berman; deposited in BMNH. [examined] syn. n.
Dugesiella coloradana Chamberlin, 1940: 35; female holotype from Sugar City, Crowley Co., Colorado, 38.231949 -103.6630016, elev. 4307ft., no collecting date, coll. unknown; deposited in
Rhechostica coloradanum Raven, 1985: 149.
Aphonopelma coloradanum Smith, 1995: 90. syn. n.
Dugesiella echina Chamberlin, 1940: 36; male holotype from Arkansas Valley, Colorado, 38.055115 -103.6217437, elev. 4182ft., 15.xi.1938, coll. unknown; deposited in
Rhechostica echinum Raven, 1985: 149.
Aphonopelma echinum Smith, 1995: 96. syn. n.
Aphonopelma gurleyi Smith, 1995: 104; male holotype from Interstate 35, near Moss Lake, Sherman, Grayson Co., Texas, 33.781417 -97.2216335, elev. 796ft., no collection date, coll. Russ Gurley; deposited in BMNH. [examined] syn. n.
Dugesiella harlingena Chamberlin, 1940: 37; female holotype from Harlingen, Cameron Co., Texas, 26.190631 -97.6961036, elev. 41ft., 1939, coll. Bryce Brown; deposited in
Rhechostica harlingenum Raven, 1985: 149.
Aphonopelma harlingenum Smith, 1995: 106. syn. n.
Aphonopelma odelli Smith, 1995: 126; female holotype from Beavers Bend State Resort Park McCurtain Co., Oklahoma, 34.13104 -94.690045, elev. 670ft., 13.vi.1979, coll. D.C. Arnold; deposited in Oklahoma State University collection. [not examined] syn. n.
Dugesiella wacona Chamberlin, 1940: 38; male holotype from Waco, McClennan Co., Texas, 31.549333 -97.1466706, elev. 498ft., 5.vii.1931, coll. unknown; deposited in
Rhechostica waconum Raven, 1985: 149.
Aphonopelma waconum Smith, 1995: 156. syn. n.
Dugesiella wichitana Chamberlin, 1940: 35; male holotype from Wichita Mtns. National Wildlife Refuge, Comanche Co., Oklahoma, 34.772106 -98.6013086, elev. 1495ft., 5.vii.1928, coll. N.M. Newport; deposited in
Rhechostica wichitanum Raven, 1985: 149.
Aphonopelma wichitanum Smith, 1995: 157. syn. n.
Diagnosis
Aphonopelma hentzi (Fig.

Description
Male and female described by
Redescription of male exemplar
(APH_0921; Fig.

Aphonopelma hentzi (Girard, 1854). A–I male specimen, APH_0921 A dorsal view of carapace, scale bar = 6mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 4.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 5mm.
Variation (14).Cl 13.58–18.04 (16.008±0.38), Cw 12.45–17.28 (14.924±0.41), LBl 1.78–2.46 (2.09±0.07), LBw 1.92–2.79 (2.371±0.09), F1 13.1–17.31 (15.509±0.36), F1w 3.19–4.50 (3.991±0.1), P1 5.36–7.63 (6.494±0.18), T1 10.71–14.18 (12.986±0.28), M1 9.15–13.03 (11.118±0.29), A1 6.03–9.13 (7.836±0.26), L1 length 44.35–60.93 (53.943±1.31), F3 10.56–16.83 (13.026±0.42), F3w 3.23–4.71 (3.999±0.11), P3 4.16–7.57 (5.675±0.23), T3 7.89–12.61 (10.126±0.33), M3 9.69–13.93 (11.859±0.33), A3 5.92–8.88 (7.786±0.21), L3 length 38.72–59.23 (48.471±1.42), F4 12.88–17.50 (15.175±0.37), F4w 3.11–4.69 (3.924±0.12), P4 4.36–7.13 (6.046±0.2), T4 10.75–15.16 (12.809±0.32), M4 12.91–17.57 (15.406±0.33), A4 6.64–9.95 (8.803±0.26), L4 length 47.54–66.69 (58.239±1.42), PTl 7.762–10.81 (9.067±0.23), PTw 2.722–3.61 (3.003±0.07), SC3 ratio 0.691–0.866 (0.769±0.01), SC4 ratio 0.324–0.968 (0.579±0.05), Coxa 1 setae = stout/thick tapered, F3 condition = normal/slightly swollen.
Redescription of female exemplar
(APH_0812; Figs



Variation (15).Cl 12.65–22.36 (17.892±0.68), Cw 11.57–19.87 (16.328±0.61), LBl 1.91–2.86 (2.399±0.08), LBw 2.13–3.42 (2.773±0.1), F1 10.41–17.13 (13.72±0.48), F1w 3.16–5.56 (4.413±0.17), P1 4.55–8.09 (6.4±0.25), T1 7.79–13.28 (10.669±0.32), M1 6.06–10.74 (7.945±0.3), A1 5.17–8.86 (6.671±0.23), L1 length 34.72–57.70 (45.404±1.51), F3 8.32–14.09 (11.281±0.43), F3w 2.79–4.99 (3.804±0.16), P3 3.94–8.06 (5.64±0.31), T3 6.11–10.56 (8.164±0.32), M3 6.46–11.65 (8.915±0.38), A3 5.46–9.03 (6.866±0.26), L3 length 30.84–53.39 (40.818±1.7), F4 10.43–17.15 (13.795±0.44), F4w 3.04–5.16 (4.072±0.15), P4 4.58–7.81 (5.906±0.23), T4 8.66–13.41 (10.851±0.31), M4 8.95–15.84 (11.924±0.47), A4 6.04–9.79 (7.672±0.27), L4 length 39.16–64.0 (50.132±1.68), SC3 ratio 0.671–0.953 (0.805±0.02), SC4 ratio 0.424–0.717 (0.547±0.02), Coxa 1 setae = stout/thick tapered. Spermathecae variation can be seen in Figures
Material examined
United States: Arkansas: Carroll: near Eureka Springs, 36.404273 -93.7356465, 1201ft., [APH_2600, unknown, 1♂, H.A. Clay,
Distribution and natural history
Aphonopelma hentzi is the most widely distributed species in the United States. Its distribution is bound to the east by the Mississippi River but these tarantulas are found in all or parts of Missouri, Arkansas, Louisiana, Kansas, Oklahoma, Texas, Colorado, New Mexico and Arizona (Fig.

Aphonopelma hentzi (Girard, 1854). A distribution of known specimens B predicted distribution; warmer colors (red, orange, yellow) represent areas of high probability of occurrence, cooler colors (blue shades) represent areas of low probability of occurrence. Of note, the lone south Texas outlier specimen in A is the harlingenum holotype.
Aphonopelma hentzi exhibits significant variation in burrowing behavior across its distribution. This species is known to inhabit freestanding burrows or scrapes (burrows under rocks, wood, etc.; see Fig.
Conservation status
Aphonopelma hentzi would be considered rare in Arizona due to its very limited distribution in the state but this species is probably the most abundant tarantula in the United States. These spiders thrive in many different habitats and are not uncommon in major metropolitan areas. More than 40 individuals were collected by one of the authors (BEH) from a single residential lawn in suburban Fort Worth, Texas (specimens not reported). Specimens have also been collected from well-maintained grounds at a cemetery just outside Downtown Dallas, Texas near a busy freeway. This species is secure.
Remarks
Other important ratios that distinguish males: A. hentzi possess a larger L3 scopulation extent (69%-86%) than A. armada (48%-63%); by possessing a larger F4/M4 (≥0.92; 0.92–1.03) than A. armada (≤0.92; 0.86–0.92); by possessing a larger PTl/T4 (>0.65; 0.65–0.77) than A. gabeli (≤0.64; 0.58–0.64). Other important ratios distinguish females: A. hentzi possess a larger L4 scopulation extent (42%-72%) than A. armada (28%–43%, with slight overlap); by possessing a smaller L1/L3 (≤1.14; 1.07–1.14) than A. moderatum (>1.14; 1.14–1.24, with slight overlap). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Of particular note,
Mitochondrial DNA (CO1) identifies A. hentzi as a polyphyletic group with respect to A. moderatum, A. anax, and A. armada (Fig.
Aphonopelma icenoglei , sp. n.
Aphonopelma mojave Prentice, 1997 (in part): 161.
Types
Male holotype (APH_2396) collected 7.5 miles north of Pipes Canyon Rd., San Bernardino Co., California, 34.303363 -116.4404925, elev. 3133ft., 14.iv.1991, coll. T.R. Prentice; deposited in
Etymology
The specific epithet is a patronym in recognition of Wendell Icenogle, an arachnologist and prolific collector of North American mygalomorph spiders. This work benefitted substantially from his help collecting specimens and his wealth of knowledge concerning tarantulas in the United States.
Diagnosis
Aphonopelma icenoglei (Fig.

Description of male holotype
(APH_2396; Fig.

Aphonopelma icenoglei sp. n. A–I male holotype, APH_2396 A dorsal view of carapace, scale bar = 2.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2.5mm E ventral view of metatarsus IV, scale bar = 2.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2.5mm.
Variation (5).Cl 7.326–8.228 (7.831±0.15), Cw 6.609–7.307 (6.976±0.11), LBl 1.031–1.069 (1.046±0.01), LBw 1.092–1.475 (1.271±0.08), F1 7.605–8.996 (8.292±0.24), F1w 1.703–1.938 (1.821±0.05), P1 2.909–3.301 (3.067±0.07), T1 6.679–7.727 (7.204±0.21), M1 5.848–7.032 (6.459±0.22), A1 4.012–4.818 (4.338±0.14), L1 length 27.396–31.441 (29.36±0.78), F3 6.881–7.558 (7.203±0.14), F3w 1.763–2.145 (1.963±0.08), P3 2.477–2.771 (2.579±0.05), T3 5.625–6.187 (5.88±0.12), M3 6.742–7.609 (6.986±0.16), A3 4.448–4.952 (4.569±0.1), L3 length 26.376–28.76 (27.216±0.48), F4 7.919–9.094 (8.46±0.22), F4w 1.691–1.902 (1.836±0.04), P4 2.549–2.869 (2.77±0.06), T4 6.914–7.825 (7.283±0.16), M4 8.368–9.267 (8.657±0.17), A4 4.702–5.595 (5.036±0.16), L4 length 30.775–33.887 (32.205±0.65), PTl 4.758–5.115 (4.953±0.07), PTw 1.548–1.77 (1.684±0.04), SC3 ratio 0.626–0.788 (0.704±0.03), SC4 ratio 0.312–0.456 (0.376±0.02), Coxa I setae = very thin tapered, F3 condition = normal.
Description of female paratype
(APH_1562; Figs


Variation (5).Cl 7.424–8.188 (7.89±0.13), Cw 6.522–7.385 (6.993±0.16), LBl 1.285–1.353 (1.322±0.01), LBw 1.368–1.615 (1.453±0.05), F1 6.565–7.644 (6.966±0.18), F1w 1.974–2.131 (2.049±0.03), P1 2.716–3.161 (2.988±0.08), T1 5.185–6.381 (5.695±0.2), M1 4.163–4.72 (4.406±0.1), A1 3.568–3.976 (3.708±0.08), L1 length 22.432–25.667 (23.763±0.54), F3 5.362–6.315 (5.807±0.16), F3w 1.766–1.979 (1.849±0.04), P3 2.217–2.711 (2.47±0.1), T3 3.995–4.755 (4.305±0.13), M3 4.216–4.864 (4.54±0.11), A3 3.90–4.389 (4.041±0.09), L3 length 20.101–22.92 (21.164±0.51), F4 6.84–8.227 (7.376±0.24), F4w 1.766–1.967 (1.897±0.03), P4 2.625–3.112 (2.836±0.09), T4 5.477–6.325 (5.799±0.14), M4 5.589–6.77 (6.14±0.19), A4 4.174–4.678 (4.436±0.1), L4 length 25.043–28.961 (26.587±0.68), SC3 ratio 0.636–0.726 (0.691±0.02), SC4 ratio 0.272–0.419 (0.365±0.03), Coxa I setae = very thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: California: Los Angeles: 0.66 miles N Ft Tejon Rd on Valyermo Rd, 34.468812 -117.8603771, 3532ft., [APH_0756-0757, 4/10/2009, 2 juv, Brent E. Hendrixson, Thomas Martin, AUMNH]; 0.67 miles S Hwy-18 on 263rd St, 34.48904 -117.6610581, 3461ft., [APH_1202, 30/7/2010, 1♀, Brent E. Hendrixson, Brendon Barnes, Nate Davis, AUMNH]; 1.15 miles N of Valyermo, Bobs Gap Rd, 34.448937 -117.829214, 3920ft., [AUMS_2511, 4/10/1992, 1♂, T.R. Prentice, AUMNH]; Piñon Hills off Hwy 138, 34.448134 -117.6687355, 4002ft., [AUMS_2515, 1993, 1♂, unknown, AUMNH]; Valyermo, 34.446108 -117.8522865, 4403ft., [APH_2400, 28/10/1989, 1♂, T.R. Prentice,
Distribution and natural history
Aphonopelma icenoglei is known from portions of the southern Mojave (Fig.

Conservation status
Aphonopelma icenoglei is not morphologically distinct from A. mojave but is genetically unique and should be considered important. The species is moderately common but may experience threats to some populations due to human encroachment and development in portions of the Mojave Desert closest to Los Angeles.
Remarks
Other important ratios that distinguish Aphonopelma icenoglei males: A. icenoglei possess a larger F1/M4 (≥0.90; 0.90–1.02) than A. joshua (≤0.84; 0.78–0.84) and A. xwalxwal (≤0.84; 0.80–0.84); important ratios that distinguish females: A. icenoglei possess a larger A3/T4 (≥0.68; 0.68–0.71) than A. atomicum (≤0.63; 0.59–0.63) and A. joshua (≤0.63; 0.55–0.63). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma iodius
Delopelma iodius Chamberlin & Ivie, 1939: 6; male holotype from 2 miles W of Castle Cliffs, Washington Co., Utah, 37.072028 -113.9197734, elev. 3663ft., 27.xi.1936, coll. unknown. Paratype males from Zion National Park, Washington Co., Utah, 37.205521 -112.9836645, elev. 3973ft., no collecting date, coll. unknown; deposited in
Rhechostica iodius Raven, 1985: 149.
Aphonopelma iodium Smith, 1995: 115.
Aphonopelma iodium Prentice, 1997: 162.
Aphonopelma iodius (spelling change; Platnick, World Spider Catalog)
Aphonopelma angusi Chamberlin, 1940: 21; male holotype from two miles west of Beaver Dam Mts., on Beaver Dam slope, Washington Co., Utah, 37.150885 -113.9170966, elev. 6332ft., 7.x.1939, coll. Dr. Angus M. Woodbury, Ross Hardy, Harold Higgins, and Robert Pendleton; deposited in
Aphonopelma angusi Smith, 1995: 72. previously synonymized by Prentice, 1997: 162.
Delopelma melanius Chamberlin & Ivie, 1939: 5; male holotype and female allotype from Salt Lake City, Salt Lake Co., Utah, 40.760779 -111.8910476, elev. 4284ft., no collecting date, coll. unknown; deposited in
Aphonopelma melanium Smith, 1995: 120. previously synonymized by Prentice, 1997: 162.
Aphonopelma nevadanum Chamberlin, 1940: 12; male holotype from Searchlight, Clark Co., Nevada, 35.465269 -114.9197015, elev. 3590ft., 2.xii.1930, coll. Geo. Carter; deposited in
Aphonopelma nevadanum Smith, 1995: 125. previously synonymized by Prentice, 1997: 162.
Aphonopelma brunnius Chamberlin, 1940: 11; male holotype from Jasper Ridge Biological Preserve, W of Stanford University, Palo Alto, Santa Clara Co., California, 37.405240 -122.2421004, elev. 378ft., 13.xi.1921, coll. C.D. Duncan; deposited in
Rhechostica brunnius Raven, 1985: 149.
Aphonopelma brunnium Smith, 1995: 79. syn. n.
Aphonopelma chamberlini Smith, 1995: 86; female holotype from Camp Roberts, outside of Paso Robles, Co., California, 35.790969 -120.7433645, elev. 642ft., no collecting date, coll. Rupert Hazen; deposited in BMNH. [examined] syn. n.
Aphonopelma iviei Smith, 1995: 115; male holotype from Death Valley, Inyo Co., California, 36.735992 -116.9701886, elev. 2761ft., 30.ix.1883, coll. Chalmers-Hunt; deposited in BMNH. [examined] syn. n.
Aphonopelma lithodomum Chamberlin, 1940: 14; male holotype from House Rock, Coconino Co., Arizona, 36.703978 -111.9471715, elev. 5168ft., 9.ix.1939, coll. D. and S. Mulaik; deposited in
Rhechostica lithodomum Raven, 1985: 149.
Aphonopelma lithodomum Smith, 1995: 119. syn. n.
Aphonopelma smithi Smith, 1995: 145; male holotype from Frank Raines County Park, off Del Puerto Canyon Rd, ~20km W of Patterson, Stanislaus Co., California, 37.422734 -121.3806975, elev. 1153ft., coll. Russell Smith, x.1992; deposited in BMNH. Two male paratypes from Patterson, Stanislaus Co., California, 37.471600 -121.1296565, elev. 107ft., x.1992, coll. Russell Smith; deposited in BMNH. [examined] syn. n.
Aphonopelma zionis Chamberlin, 1940: 24; male holotype from Zion National Park, near entrance, Washington Co., Utah, 37.205521 -112.9836644, elev. 3973ft., 16.viii.1925, coll. Dr. A.M. Woodbury; deposited in
Rhechostica zionis Raven, 1985: 149.
Aphonopelma zionis Smith, 1995: 157. syn. n.
Diagnosis
Aphonopelma iodius (Fig.

Description
Originally described by
Redescription of male exemplar
(APH_2015; Fig.

Aphonopelma iodius (Chamberlin & Ivie, 1939). A–I male specimen, APH_2015 A dorsal view of carapace, scale bar = 7mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 5mm E ventral view of metatarsus IV, scale bar = 4.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 4.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 4.5mm.
Variation (23).Cl 9.69–17.75 (13.959±0.45), Cw 8.51–16.49 (13.019±0.43), LBl 1.21–2.23 (1.734±0.06), LBw 1.397–2.56 (2.079±0.06), F1 10.336–16.69 (14.442±0.38), F1w 2.13–4.38 (3.326±0.12), P1 3.916–6.92 (5.704±0.16), T1 9.343–14.65 (12.294±0.28), M1 7.918–14.62 (11.76±0.34), A1 5.204–9.03 (7.508±0.21), L1 length 36.717–60.79 (51.709±1.33), F3 8.88–14.81 (12.472±0.34), F3w 2.27–4.51 (3.525±0.12), P3 3.248–6.39 (4.935±0.17), T3 6.934–11.84 (9.943±0.28), M3 8.446–14.28 (12.058±0.35), A3 5.067–8.63 (7.204±0.21), L3 length 32.69–54.63 (46.665±1.35), F4 10.308–17.21 (14.341±0.39), F4w 2.127–4.27 (3.258±0.12), P4 3.614–6.59 (5.277±0.17), T4 8.674–14.44 (12.274±0.31), M4 10.951–18.26 (15.385±0.4), A4 5.39–9.58 (7.96±0.24), L4 length 39.514–63.97 (55.346±1.51), PTl 6.198–10.293 (8.387±0.22), PTw 2.04–3.30 (2.687±0.07), SC3 ratio 0.789–0.961 (0.896±0.01), SC4 ratio 0.623–0.876 (0.746±0.01), Coxa I setae = thin tapered/tapered, F3 condition = normal/slightly swollen.
Description of female exemplar
(APH_2016; Figs

Aphonopelma iodius (Chamberlin & Ivie, 1939). A–E female specimen, APH_2016 A dorsal view of carapace, scale bar = 5.5mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 4mm D ventral view of metatarsus IV, scale bar = 5mm E prolateral view of L pedipalp and palpal tibia.



Variation (19).Cl 8.657–21.4 (15.619±0.68), Cw 7.51–18.6 (14.079±0.6), LBl 1.322–2.73 (2.086±0.09), LBw 1.578–3.25 (2.414±0.11), F1 7.493–16.71 (12.928±0.54), F1w 2.162–4.72 (3.69±0.16), P1 3.155–7.60 (5.581±0.26), T1 6.154–13.02 (10.428±0.41), M1 4.613–11.89 (8.701±0.43), A1 3.861–7.94 (6.577±0.24), L1 length 25.276–56.01 (44.214±1.84), F3 6.452–13.52 (11.003±0.42), F3w 1.923–4.02 (3.281±0.13), P3 2.599–6.65 (4.898±0.22), T3 4.381–10.07 (8.138±0.32), M3 4.766–11.50 (8.808±0.36), A3 3.978–7.72 (6.494±0.22), L3 length 22.176–48.63 (39.341±1.49), F4 7.32–17.23 (13.256±0.53), F4w 1.86–4.28 (3.405±0.14), P4 3.187–6.67 (5.289±0.21), T4 6.243–12.71 (10.466±0.39), M4 7.325–15.1 (12.038±0.43), A4 4.46–8.91 (7.111±0.22), L4 length 28.535–58.40 (48.161±1.72), SC3 ratio 0.724–0.949 (0.855±0.02), SC4 ratio 0.593–0.829 (0.739±0.02), Coxa I setae = tapered. Spermathecae variation can be seen in Figures
Material examined
United States: Arizona: Coconino: House Rock - off 89A, 36.729414 -112.0373865, 5337ft., [AUMS_3299, 3/7/89, 1♂, T.R. Prentice, AUMNH]; [AUMS_2386, 2/7/89, 1♂, T.R. Prentice, AUMNH]; off US-89A, Soap Creek Rd, 36.72439 -111.75561, 4211ft., [APH_0303, 8/10/07, 1 juv, Zach Valois, AUMNH]; Mohave: Road to Summit Springs, .25 miles from the UT/AZ border, 36.99566 -113.9177934, 2533ft., [APH_2216, 19/10/1993, 1♂, T.R. Prentice,
Distribution and natural history
Aphonopelma iodius has a wide distribution that stretches across several diverse habitats from the Bay Area of California, south along the Coast Ranges across the Transverse Ranges, into the Mojave Desert (Fig.

Conservation status
Aphonopelma iodius is one of the most widespread and abundant species in the United States. The species is secure.
Remarks
Aphonopelma iodius is a problematic species. As presently defined, there are no major morphological features that can be used to distinguish A. iodius from its sister lineages, and there is not enough evidence at this time to allow us to recognize cryptic diversity within the A. iodius lineage (except for the geographically, genetically, and morphologically distinct A. johnnycashi). During evaluation of traditional two-dimensional PCA morphospace and three-dimensional PCA morphospace (PC1~PC2~PC3), males and females of A. iodius do not separate from any of the other species in the Iodius species group (A. chalcodes, A. eutylenum, A. johnnycashi), but do separate from A. steindachneri and their syntopic miniature tarantulas (A. atomicum, A. icenoglei, A. joshua, A. mojave, and A. prenticei) (see Suppl. material
We examined the holotypes and freshly collected topotypic material of A. brunnius, A. chamberlini, A. iviei, A. lithodomum, A. smithi, and A. zionis. We also examined the holotypes and freshly collected topotypic material of three previously synonymized species: A. angusi, A. melanium, and A. nevadanum. Our morphological and molecular analyses fail to recognize these nine species as separate, independently evolving lineages. As a consequence, we consider A. brunnius, A. chamberlini, A. iviei, A. lithodomum, A. smithi, and A. zionis junior synonyms of A. iodius and confirm the synonymizations of A. angusi, A. melanium, and A. nevadanum proposed by
Prentice formally changed the spelling of A. iodius to A. iodium because the gender is neuter. But, Chamberlin & Ivie (1939, pgs. 5–7) described Delopelma (Aphonopelma) iodius as “...easily distinguishable from melanius, which it apparently resembles most closely, by its rich rust color as contrasted with the dark metallic color of melanius.” It is also noted that in
Mitochondrial DNA (CO1) identifies A. iodius, as presently defined, as a polyphyletic species with individuals grouping together with members of A. chalcodes and A. johnnycashi (Fig.
Aphonopelma johnnycashi , sp. n.
Types
Male holotype (APH_2007) from NE of Ione, on Sutter Creek/Ione Rd, Amador Co., California, 38.37543 -120.902881, elev. 570ft., 12.x.10, coll. Chris Hamilton; deposited in AUMNH. Paratype female (APH_3080) from Coarsegold, off Hwy 41, behind CAL Fire station, Madera Co., California, 37.2507 -119.703921, elev. 2226ft., 10.vii.12, coll. Chris Hamilton; deposited in AUMNH. Paratype male (APH_2032) from Ione, Amador Co., California, 38.363017 -120.926951, elev. 430ft., 1.x.10, coll. Mike Dame; deposited in
Etymology
The specific epithet is in honor of the country music legend, Johnny Cash. This species can be found near the area of Folsom Prison in California, and like Cash’s distinctive style of dress (where he was referred to as “the man in black”), mature males of this species are generally black in color.
Diagnosis
Aphonopelma johnnycashi (Fig.

Significant measurements that distinguish male A. johnnycashi from its closely related phylogenetic and syntopic species are T3 and the extent of scopulation on metatarsus IV. Male A. johnnycashi can be distinguished by possessing a larger T1/T3 (≥1.25; 1.25–1.31) than A. eutylenum (≤1.23; 1.16–1.23); by possessing a larger A1/T3 (≥0.81; 0.81–0.91) than A. chalcodes (≤0.79; 0.67–0.79); and by possessing a larger L4 scopulation extent (70%-76%) than A. steindachneri (21%-31%). There are no significant measurements that separate male A. johnnycashi from A. iodius. The most significant measurement that distinguishes female A. johnnycashi from A. steindachneri is extent of scopulation on metatarsus IV. There are no significant measurements that separate female A. johnnycashi from the other members of the Iodius species group. Female A. johnnycashi can be distinguished by possessing a larger L4 scopulation extent (67%-82%) than A. steindachneri (24%-34%).
Description of male holotype
(APH_2007; Fig.

Aphonopelma johnnycashi sp. n. A–I male holotype, APH_2007 A dorsal view of carapace, scale bar = 4mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 3.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 4.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 5.5mm.
Variation (5).Cl 13.42–13.71 (13.565±0.09), Cw 13.15–13.19 (13.17±0.01), LBl 1.54–1.67 (1.605±0.04), LBw 1.96–2.07 (2.015±0.03), F1 13.07–13.29 (13.18±0.07), F1w 3.23–3.52 (3.375±0.09), P1 5.04–5.69 (5.365±0.21), T1 10.58–11.23 (10.905±0.21), M1 10.57–10.74 (10.655±0.05), A1 7.6–7.69 (7.645±0.03), L1 length 47.08–48.42 (47.75±0.42), F3 11.05–11.16 (11.105±0.03), F3w 3.43–3.55 (3.49±0.04), P3 4.47–4.69 (4.58±0.07), T3 8.31–8.55 (8.43±0.08), M3 10.24–10.74 (10.49±0.16), A3 6.81–7.04 (6.925±0.07), L3 length 41.33–41.73 (41.53±0.13), F4 12.77–12.84 (12.805±0.02), F4w 3.16–3.29 (3.225±0.04), P4 4.7–4.9 (4.8±0.06), T4 10.9–11.33 (11.115±0.14), M4 13.72–14.53 (14.125±0.26), A4 7.46–7.85 (7.655±0.12), L4 length 49.82–51.18 (50.5±0.43), PTl 6.646–8.429 (7.538±0.56), PTw 2.31–2.801 (2.556±0.16), SC3 ratio 0.71–0.758 (0.734±0.02), SC4 ratio 0.709–0.762 (0.736±0.02), Coxa I setae = tapered/thin tapered, F3 condition = normal.
Description of female paratype
(APH_3080; Figs


Variation (6).Cl 15.01–15.81 (15.41±0.23), Cw 13.89–14.01 (13.95±0.03), LBl 1.94–2.05 (1.995±0.03), LBw 2.47–2.48 (2.475±0), F1 11.56–12.64 (12.1±0.31), F1w 3.63–3.64 (3.635±0), P1 5.39–5.84 (5.615±0.13), T1 8.76–9.82 (9.29±0.31), M1 7.36–8.22 (7.79±0.25), A1 5.95–6.29 (6.12±0.1), L1 length 39.36–42.47 (40.915±0.9), F3 9.58–10.20 (9.89±0.18), F3w 3.16–3.18 (3.17±0.01), P3 4.63–4.64 (4.635±0), T3 6.91–7.52 (7.215±0.18), M3 7.61–7.94 (7.775±0.1), A3 5.84–5.86 (5.85±0.01), L3 length 34.57–36.16 (35.365±0.46), F4 11.92–12.28 (12.1±0.1), F4w 3.31–3.39 (3.35±0.02), P4 4.97–5.10 (5.035±0.04), T4 9.23–9.58 (9.405±0.1), M4 10.28–11.11 (10.695±0.24), A4 6.49–6.57 (6.53±0.02), L4 length 42.97–44.56 (43.765±0.46), SC3 ratio 0.774–0.917 (0.846±0.04), SC4 ratio 0.741–0.817 (0.779±0.02), Coxa I setae = tapered/thin tapered, F3 condition = normal. Spermathecae variation can be seen in Figure
Material examined
United States: California: Amador: N of Ione/SW of Plymouth, on Muller Rd, off Carbondale Rd and Hwy 6, 38.44191 -120.921641, 678ft., [APH_2004-2006, 12/10/10, 1♀, 2 juv, Chris Hamilton, AUMNH]; NE of Ione, on Sutter Creek/Ione Rd, 38.37543 -120.902881, 570ft., [APH_2007-2009, 12/10/10, 3♂, Chris Hamilton, AUMNH]; Ione, 38.363017 -120.926951, 430ft., [APH_2032-2034, 1/10/10, 2♂, 1♀, Mike Dame, AUMNH &
Distribution and natural history
Aphonopelma johnnycashi has a distribution running along the western foothills of the Sierra Nevada Mountains in California (Fig.

Conservation status
Aphonopelma johnnycashi is common across its distribution along the foothills and lower elevations of the western Sierra Nevada (Fig.
Remarks
Aphonopelma johnnycashi is a member of the problematic Iodius species group. As of now, we only have evidence to split A. johnnycashi from the remaining populations of A. iodius. Other important ratios that distinguish males: A. johnnycashi possess a smaller T3/A3 (≤1.27; 1.18–1.27) than A. eutylenum (≥1.30; 1.30–1.35) and A. steindachneri (≥1.33; 1.33–1.52); by possessing a smaller F3/A3 (≤1.63; 1.56–1.63) than A. chalcodes (≥1.63; 1.63–1.88); by possessing a larger L3 scopulation extent (71%–82%) than A. steindachneri (40%–54%). Other important ratios that distinguish females: A. johnnycashi possess a larger M1/M4 (≥0.70; 0.70–0.74) than A. steindachneri (≤0.67; 0.62–0.67). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Mitochondrial DNA (CO1) identifies A. johnnycashi as a polyphyletic species with individuals grouping variously together with different lineages of A. iodius (Fig.
Aphonopelma joshua
Aphonopelma joshua Prentice, 1997: 150; male holotype from 2.3 mi. below the Covington Flats entrance to Joshua Tree National Park, San Bernardino Co., California, 34.045652 -116.3155664, elev. 4683ft., 6.ix.1992, coll. Thomas R. Prentice; deposited in
Diagnosis
Aphonopelma joshua (Fig.

Description
Male and female originally described by
Redescription of male exemplar
(APH_1476; Fig.

Aphonopelma joshua Prentice, 1997. A–I male specimen, APH_1476 A dorsal view of carapace, scale bar = 3mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 3mm E ventral view of metatarsus IV, scale bar = 3mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2.5mm.
Variation (6).Cl 7.549–8.48 (7.931±0.14), Cw 6.92–7.62 (7.264±0.11), LBl 1.101–1.339 (1.181±0.04), LBw 1.208–1.709 (1.417±0.08), F1 8.656–9.24 (8.941±0.1), F1w 1.81–2.145 (1.964±0.05), P1 2.84–3.48 (3.158±0.1), T1 7.899–8.96 (8.302±0.17), M1 7.135–7.868 (7.461±0.11), A1 5.13–5.41 (5.266±0.05), L1 length 31.749–34.445 (33.127±0.42), F3 8.028–8.527 (8.23±0.07), F3w 1.97–2.678 (2.5±0.11), P3 2.655–3.20 (2.978±0.1), T3 6.76–7.597 (7.071±0.14), M3 8.213–9.248 (8.707±0.16), A3 5.208–6.21 (5.542±0.14), L3 length 30.918–34.04 (32.527±0.51), F4 9.225–9.93 (9.565±0.11), F4w 1.34–2.084 (1.846±0.12), P4 2.82–3.14 (3.004±0.06), T4 8.172–9.04 (8.676±0.15), M4 10.39–11.45 (11.005±0.19), A4 5.23–6.30 (5.862±0.16), L4 length 36.896–39.52 (38.112±0.41), PTl 4.469–5.021 (4.786±0.08), PTw 1.433–1.58 (1.506±0.03), SC3 ratio 0.532–0.706 (0.628±0.03), SC4 ratio 0.242–0.393 (0.342±0.02), Coxa I setae = very thin tapered, F3 condition = swollen.
Description of female exemplar
(APH_2306; Figs


Variation (6).Cl 6.05–9.82 (8.238±0.59), Cw 5.13–8.02 (7.057±0.48), LBl 0.901–1.36 (1.188±0.07), LBw 1.133–1.52 (1.383±0.07), F1 5.048–8.45 (6.805±0.51), F1w 1.436–2.256 (1.869±0.14), P1 2.07–3.322 (2.712±0.22), T1 4.231–6.91 (5.813±0.4), M1 2.853–5.35 (4.235±0.4), A1 2.597–4.16 (3.541±0.22), L1 length 16.909–28.16 (23.106±1.72), F3 4.163–7.21 (5.864±0.45), F3w 1.319–2.154 (1.732±0.12), P3 1.839–2.92 (2.453±0.2), T3 3.241–5.49 (4.486±0.32), M3 3.484–5.82 (4.887±0.4), A3 2.98–4.147 (3.718±0.2), L3 length 15.707–25.52 (21.409±1.55), F4 5.668–8.55 (7.329±0.47), F4w 1.265–2.182 (1.818±0.14), P4 2.175–3.54 (2.745±0.22), T4 4.788–7.34 (6.271±0.38), M4 5.019–8.15 (6.732±0.48), A4 3.488–4.73 (4.195±0.22), L4 length 21.138–32.31 (27.766±1.99), SC3 ratio 0.627–0.696 (0.657±0.01), SC4 ratio 0.345–0.399 (0.368±0.01), Coxa I setae = very thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: California: Riverside: Joshua Tree National Park, 34.028757 -116.3156656, 4880ft., [APH_0883, 2007, 1♀, Josh Richards, AUMNH]; Joshua Tree National Park, 1 mile off Keys View Road, 33.87529 -116.0387775, 3589ft., [APH_2306, 3/5/1989, 1♀, T.R. Prentice,
Distribution and natural history
Aphonopelma joshua has a rather limited distribution within Joshua Tree National Park and surrounding areas in south-central San Bernardino and north-central Riverside Counties, California (Fig.

Conservation status
Despite its narrow distribution, A. joshua is largely protected by Joshua Tree National Park. Recreational activities may pose some concern but this species is common throughout the park and is likely secure.
Remarks
Aphonopelma joshua is one of the larger species of miniature Aphonopelma and is morphologically very similar to the new species A. xwalxwal. Other important ratios that distinguish males: A. joshua possess a smaller F1/M4 (≤0.84; 0.78–0.84) than A. atomicum (≥0.89; 0.89–0.91), A. iodius (≥0.89; 0.89–1.04), A. icenoglei (≥0.90; 0.90–1.02), A. mojave (≥0.87; 0.87–0.94), A. prenticei (≥0.91; 0.91–0.95); by possessing a smaller L1/L3 (≤1.03; 0.99–1.03) than A. iodius (≥1.05; 1.05–1.20), A. mojave (≥1.06; 1.06–1.11), A. prenticei (≥1.05; 1.05–1.11), A. xwalxwal (≥1.07; 1.07–1.12); by possessing a smaller L3 scopulation extent (53%-71%) than A. iodius (78%-96%); by possessing a smaller L4 scopulation extent (24%-39%) than A. iodius (62%-88%); by possessing a smaller M1/F3 (<0.92; 0.88–0.92) than A. xwalxwal (>0.98; 0.98–1.02). Other important ratios that distinguish females: A. joshua possess a larger Cl/P1 (≥2.77; 2.77–3.53) than A. atomicum (≤2.50; 2.43–2.50); by possessing a smaller L1/Cl (≤2.87; 2.67–2.87) than A. icenoglei (≥2.92; 2.92–3.13); by possessing a smaller Cl/M3 (≤1.74; 1.62–1.74) than A. prenticei (≥1.77; 1.77–2.20); by possessing a smaller A1/T4 (≤0.59; 0.54–0.59) than A. mojave (≥0.58; 0.58–0.67; small overlap). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Aphonopelma madera , sp. n.
Types
Male holotype (APH_3177) from Madera Canyon, Coronado National Forest, on road to Bog Springs Campground, Pima Co., Arizona, 31.72812 -110.8788181, elev. 4809ft., 12.xi.2013, coll. Chris A. Hamilton and Brent E. Hendrixson; deposited in AUMNH. Paratype female (APH_1393) from Madera Canyon, Mt. Wrightson picnic area, Santa Cruz Co., Arizona, 31.71273 -110.8749361, elev. 5401ft., 5.x.2011, coll. Brent E. Hendrixson and Thomas Martin; deposited in AUMNH. Paratype male (APH_1627) from Madera Canyon, Madera Picnic Area, Cochise Co., Arizona, 31.72722 -110.8808121, elev. 4809ft., 15.xi.2012, coll. Brent E. Hendrixson; deposited in
Etymology
The specific epithet is a noun in apposition taken from type locality, Madera Canyon, in the Santa Rita Mountains where this species was first discovered.
Diagnosis
Aphonopelma madera (Fig.

Description of male holotype
(APH_3177; Fig.

Aphonopelma madera sp. n. A–I male holotype, APH_3177 A dorsal view of carapace, scale bar = 2.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2.5mm E ventral view of metatarsus IV, scale bar = 3mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 3mm.
Variation (6).Cl 7.815–9.43 (8.488±0.22), Cw 7.478–8.57 (7.861±0.16), LBl 0.986–1.138 (1.065±0.03), LBw 1.139–1.449 (1.321±0.06), F1 8.374–9.78 (8.918±0.22), F1w 1.917–2.11 (2.012±0.03), P1 3.016–3.59 (3.302±0.08), T1 7.321–8.89 (7.813±0.23), M1 4.977–5.52 (5.213±0.1), A1 3.542–4.84 (4.135±0.22), L1 length 27.831–32.29 (29.38±0.7), F3 6.215–7.06 (6.611±0.12), F3w 1.9–2.13 (1.991±0.03), P3 2.581–3.136 (2.851±0.09), T3 4.618–5.70 (5.029±0.15), M3 5.038–5.89 (5.467±0.12), A3 3.945–4.56 (4.29±0.1), L3 length 22.958–26.13 (24.247±0.46), F4 7.521–8.92 (8.154±0.21), F4w 1.82–1.99 (1.905±0.03), P4 2.603–3.234 (2.983±0.09), T4 6.379–7.51 (6.92±0.16), M4 7.043–8.20 (7.58±0.2), A4 4.288–5.89 (5.027±0.25), L4 length 28.706–33.63 (30.663±0.83), PTl 4.947–6.201 (5.447±0.17), PTw 1.79–2.158 (2.043±0.06), SC3 ratio 0.459–0.702 (0.548±0.03), SC4 ratio 0.238–0.481 (0.339±0.03), Coxa I setae = very thin tapered, F3 condition = normal.
Description of female paratype
(APH_1393; Figs


Variation (5).Cl 9.17–15.72 (12.176±1.11), Cw 8.43–13.73 (10.914±0.95), LBl 1.18–1.76 (1.475±0.09), LBw 1.51–2.19 (1.781±0.11), F1 7.332–12.62 (9.914±0.86), F1w 2.342–3.82 (3.088±0.25), P1 3.046–5.31 (4.265±0.39), T1 5.999–9.22 (7.798±0.57), M1 3.416–6.56 (5.147±0.55), A1 3.951–5.58 (4.83±0.27), L1 length 23.744–39.29 (31.955±2.62), F3 7.36–8.79 (8.203±0.34), F3w 2.46–3.34 (2.845±0.19), P3 3.34–4.52 (3.768±0.27), T3 5.07–6.84 (5.898±0.37), M3 5.11–6.81 (6±0.38), A3 4.69–5.73 (5.253±0.22), L3 length 25.67–32.69 (29.12±1.5), F4 7.514–12.38 (9.841±0.8), F4w 2.188–3.58 (2.816±0.25), P4 3.234–4.87 (4.003±0.29), T4 6.137–9.55 (7.823±0.59), M4 6.197–9.91 (8.183±0.66), A4 4.842–6.47 (5.694±0.29), L4 length 27.924–43.18 (35.545±2.61), SC3 ratio 0.483–0.684 (0.595±0.04), SC4 ratio 0.296–0.348 (0.33±0.01), Coxa I setae = tapered/thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Cochise: Carr Canyon Rd, 31.449658 -110.2820281, 5257ft., [APH_1595, 8/11/2012, 1♂, Brent E. Hendrixson, AUMNH]; Copper Canyon, Huachuca Mtns, 31.363172 -110.299795, 6082ft., [APH_0881, 2007, 1 juv, Josh Richards, AUMNH]; [APH_0980, 2007, 1♀, Josh Richards, AUMNH]; Huachuca Mtns, Ash Canyon, 31.38339 -110.244862, 5290ft., [APH_1249, 11/2010, 1♂, Jim Murray, AUMNH]; Huachuca Mtns, Garden Canyon Rd, 31.47306 -110.351112, 5360ft., [APH_1250-1251, 4/12/2010, 2♂,
Distribution and natural history
Aphonopelma madera is known from the Huachuca, Pajarito, and Santa Rita Mountains in southeastern Arizona at elevations ranging from 1230 to 2110 meters, inhabiting the Madrean Archipelago Level III Ecoregion. The species has been collected from riparian, oak-grassland, oak woodland, and pine-oak woodland communities (Figs

Conservation status
Of the three sky island endemics, A. madera is represented by the most specimens, has the largest distribution, and is the only species that is not restricted to a single mountain range. The species appears to be fairly common in Madera Canyon but we would not argue that the conservation status for this species is secure at the moment. Our sampling of mitochondrial haplotypes (
Remarks
Aphonopelma madera is morphologically similar to other high-elevation species in the Marxi species group, especially A. catalina (see
Aphonopelma mareki , sp. n.
Types
Male holotype (APH_1615) collected 0.4 miles E Hwy-89 on Stanton Rd, Yavapai Co., Arizona, 34.18203 -112.8188621, elev. 2852ft., 12.xi.2012, coll. Brent E. Hendrixson; deposited in AUMNH. Paratype female (APH_1590) from Bartlett Dam Rd, Maricopa Co., Arizona, 33.847258 -111.7932911, elev. 2793ft., 7.xi.2012, coll. Brent E. Hendrixson; deposited in AUMNH. Paratype male (APH_1569) from just outside N boundary of Montezuma Well National Monument, along Beaver Creek Rd, Yavapai Co., Arizona, 34.653811 -111.7574971, elev. 3596ft., 27.x.2012, coll. Brent E. Hendrixson; deposited in
Etymology
The specific epithet is a patronym in recognition of our friend, colleague, and myriapod biologist Paul Marek, who collected several important specimens examined as part of this revision.
Diagnosis
Aphonopelma mareki (Fig.

Description of male holotype
(APH_1615; Fig.

Aphonopelma mareki sp. n. A–I male holotype, APH_1615 A dorsal view of carapace, scale bar = 2.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2mm E ventral view of metatarsus IV, scale bar = 2mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2.5mm.
Variation (5).Cl 5.41–6.411 (5.981±0.19), Cw 4.751–5.844 (5.348±0.18), LBl 0.711–0.858 (0.78±0.02), LBw 0.857–1.16 (0.981±0.06), F1 6.017–6.875 (6.501±0.14), F1w 1.306–1.622 (1.449±0.05), P1 1.995–2.609 (2.382±0.1), T1 5.392–5.782 (5.62±0.08), M1 4.137–4.531 (4.352±0.07), A1 2.729–3.457 (3.183±0.13), L1 length 20.39–22.933 (22.039±0.47), F3 4.776–5.561 (5.248±0.15), F3w 1.451–1.875 (1.655±0.07), P3 1.726–1.975 (1.86±0.04), T3 3.732–4.434 (4.098±0.12), M3 4.271–5.129 (4.66±0.16), A3 3.092–3.607 (3.39±0.09), L3 length 17.686–20.706 (19.256±0.53), F4 5.849–6.639 (6.296±0.14), F4w 1.268–1.589 (1.399±0.06), P4 1.757–2.207 (2.022±0.08), T4 4.729–5.931 (5.475±0.21), M4 5.494–6.687 (6.158±0.24), A4 3.463–4.148 (3.858±0.12), L4 length 21.575–25.57 (23.808±0.74), PTl 3.542–4.044 (3.793±0.09), PTw 1.285–1.499 (1.392±0.03), SC3 ratio 0.502–0.563 (0.533±0.01), SC4 ratio 0.186–0.421 (0.286±0.04), Coxa I setae = very thin tapered, F3 condition = normal & slightly swollen.
Description of female paratype
(APH_1590; Figs


Variation (6).Cl 7.315–8.778 (7.717±0.22), Cw 6.021–7.405 (6.649±0.19), LBl 1.012–1.195 (1.116±0.03), LBw 1.171–1.364 (1.281±0.03), F1 5.83–7.175 (6.362±0.18), F1w 1.839–2.276 (2.067±0.07), P1 2.412–3.416 (2.8±0.16), T1 4.712–5.646 (5.159±0.14), M1 3.348–4.089 (3.684±0.13), A1 2.981–3.742 (3.318±0.11), L1 length 19.528–23.553 (21.323±0.62), F3 4.494–5.928 (5.185±0.21), F3w 1.669–2.058 (1.794±0.06), P3 1.856–2.736 (2.341±0.13), T3 3.194–4.041 (3.647±0.12), M3 3.336–4.525 (3.859±0.16), A3 3.127–3.606 (3.455±0.07), L3 length 16.007–20.751 (18.487±0.66), F4 5.886–7.124 (6.504±0.17), F4w 1.679–2.053 (1.842±0.06), P4 2.285–3.222 (2.617±0.14), T4 4.687–5.565 (5.247±0.14), M4 4.705–5.878 (5.463±0.18), A4 3.636–4.188 (3.958±0.09), L4 length 21.384–25.866 (23.789±0.64), SC3 ratio 0.429–0.617 (0.554±0.03), SC4 ratio 0.218–0.273 (0.241±0.01), Coxa I setae = thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Coconino: Schnebly Hill Road, Sedona, 34.86533 -111.7530035, 4328ft., [AUMS_2272, 15/5/1992, 1♂, unknown, AUMNH]; Gila: 1.1 miles NW AZ-87 on Barnhardt Trail Rd (FR 419), 34.09815 -111.361651, 3200ft., [APH_0345, 10/5/2008, 1 juv, Brent E. Hendrixson, Zach Valois, Josh Richards, AUMNH]; 1.4 miles E US-87 on Control Rd, 34.35949 -111.403021, 5430ft., [APH_0347, 10/5/2008, 1 juv, Brent E. Hendrixson, Zach Valois, Josh Richards, AUMNH]; Mazatzal Mtns, Four Peaks, Pigeon Spring Rd. junction, 33.72156 -111.337531, 5780ft., [APH_0297, 6/10/2007, 1♀, Zach Valois,
Distribution and natural history
Aphonopelma mareki can be found inhabiting the Arizona/New Mexico Mountains and Sonoran Basin and Range Level III Ecoregions of central Arizona (Fig.

Conservation status
Aphonopelma mareki is very common throughout its distribution but can be difficult to find due to the cryptic nature of its burrows and small window of activity during the year. The species is secure.
Remarks
Other important ratios that distinguish Aphonopelma mareki males: A. mareki possess a larger T1/M1 (≥1.26; 1.26–1.33) than A. chalcodes (≤1.13; 0.99–1.13) and A. prenticei (≤1.16; 1.07–1.16), and smaller than A. marxi (≥1.39; 1.39–1.52); a larger PTl/M1 (≥0.83; 0.83–0.95) than A. chalcodes (≤0.79; 0.67–0.79), A. prenticei (≤0.82; 0.72–0.82), and A. superstitionense (≤0.82; 0.79–0.82); a larger T3/M3 (≥0.85; 0.85–0.94) than A. paloma (≤0.85; 0.79–0.85) and A. prenticei (≤0.85; 0.80–0.85); a smaller PTl/F1 (≤0.61; 0.53–0.61) than A. saguaro (≥0.61; 0.61–0.64). Other important ratios that distinguish A. mareki females: A. mareki possess a larger F3L/W (≥2.66; 2.66–3.00) than A. saguaro (2.41 ± (only 1 specimen)); a larger M3/M4 (≥0.66; 0.66–0.77) than A. superstitionense (0.62 ± (only 1 specimen)); a smaller M4/A4 (≤1.48; 1.23–1.48) than A. chalcodes (≥1.61; 1.61–1.98); a smaller F1/M3 (≤1.75; 1.58–1.75) than A. marxi (≥1.76; 1.76–1.91). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Mitochondrial DNA (CO1) identifies A. mareki as a polyphyletic group with some members grouping with the A. superstitionense lineage (Fig.
Aphonopelma marxi
Eurypelma marxi Simon, 1891: 324; no original labeled types known to exist; male neotype designated (
Delopelma marxi Petrunkevitch, 1939: 252.
Rhechostica marxi Raven, 1985: 149.
Aphonopelma marxi Smith, 1995: 119.
Aphonopelma marxi Prentice, 1997: 147.
Delopelma simulatum Chamberlin & Ivie, 1939: 8; male holotype from Fruita, Wayne Co., Utah, 38.285536 -111.2468366, elev. 5442ft., 14.vii.1931, coll. Gertsch and Johnson; deposited in
Aphonopelma (Delopelma) simulatum Chamberlin, 1940: 26.
Rhechostica simulatum Raven, 1985: 149.
Aphonopelma simulatum Smith, 1995: 144. previously synonymized by Prentice, 1997: 147.
Aphonopelma behlei Chamberlin, 1940: 26; male holotype and male paratype from Grand Canyon Village, Coconino Co., Arizona, 36.054444 -112.1401114, elev. 6882ft., 15.ix.1939, coll. Dr. W.H. Behle; deposited in
Rhechostica behlei Raven, 1985: 149.
Aphonopelma behlei Smith, 1995: 76. syn. n.
Aphonopelma vogeli Smith, 1995: 154; male holotype from Aztec, San Juan Co., New Mexico, 36.822226 -107.9928466, elev. 5657ft., 20.x.1982, coll. W.A. Drew; deposited in Oklahoma State University collection [presumed lost]
Aphonopelma vogelae - male neotype designated (APH_1431) from 0.4 miles E. CR-7635 along dirt road, San Juan Co., New Mexico, 36.28073 -107.87572, elev. 6721ft., 4.xi.2011, coll. Amber Williams; deposited in AUMNH. [examined] syn. n.
Diagnosis
Aphonopelma marxi (Fig.

Description
Male originally described by
Redescription of male exemplar
(APH_1396; Fig.

Aphonopelma marxi (Simon, 1891). A–I male specimen, APH_1396 A dorsal view of carapace, scale bar = 3.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 3.5mm E ventral view of metatarsus IV, scale bar = 3mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 3.5mm.
Variation (8).Cl 8.32–10.53 (9.348±0.24), Cw 8.03–10.55 (8.854±0.28), LBl 1.087–1.397 (1.235±0.03), LBw 1.222–1.64 (1.446±0.05), F1 9.556–12.45 (10.532±0.36), F1w 1.957–2.89 (2.444±0.11), P1 3.546–4.81 (3.896±0.15), T1 7.366–10.21 (8.463±0.3), M1 4.912–7.33 (5.847±0.26), A1 4.61–6.05 (5.051±0.17), L1 length 30.157–40.85 (33.79±1.2), F3 7.184–9.23 (7.864±0.24), F3w 2.12–2.67 (2.315±0.06), P3 2.768–3.66 (3.179±0.11), T3 5.386–7.37 (5.985±0.22), M3 5.247–7.27 (6.183±0.23), A3 4.902–5.964 (5.421±0.14), L3 length 26.005–33.32 (28.632±0.86), F4 8.677–11.04 (9.536±0.28), F4w 2.04–2.79 (2.268±0.09), P4 2.943–4.11 (3.398±0.13), T4 7.058–9.39 (8.073±0.25), M4 7.548–9.74 (8.539±0.24), A4 5.573–6.80 (6.247±0.15), L4 length 32.388–41.08 (35.791±1), PTl 5.663–6.774 (6.152±0.13), PTw 1.94–2.49 (2.134±0.06), SC3 ratio 0.524–0.668 (0.605±0.02), SC4 ratio 0.38–0.465 (0.411±0.01), Coxa I setae = thin tapered, F3 condition = normal.
Description of female exemplar
(APH_0452; Figs


Variation (5).Cl 13.49–15.30 (14.454±0.34), Cw 12.85–15.02 (13.714±0.38), LBl 1.49–2.27 (1.86±0.13), LBw 1.95–2.31 (2.148±0.07), F1 11.48–13.62 (12.498±0.36), F1w 3.68–4.47 (4.004±0.16), P1 5.24–6.76 (5.95±0.26), T1 9.17–10.91 (9.914±0.31), M1 6.22–7.23 (6.76±0.16), A1 5.58–7.13 (6.276±0.27), L1 length 37.69–44.70 (41.398±1.17), F3 8.87–10.83 (9.832±0.32), F3w 2.94–4.06 (3.39±0.2), P3 4.36–5.54 (4.912±0.19), T3 6.36–7.93 (6.958±0.28), M3 6.49–7.30 (6.818±0.14), A3 5.45–6.66 (6.182±0.23), L3 length 31.67–38.0 (34.702±1.04), F4 10.85–13.28 (11.936±0.4), F4w 3.36–3.81 (3.566±0.1), P4 4.35–5.50 (4.936±0.21), T4 8.64–9.54 (9.146±0.18), M4 8.71–10.69 (9.734±0.32), A4 6.18–7.61 (7.124±0.28), L4 length 38.73–46.55 (42.876±1.26), SC3 ratio 0.597–0.74 (0.674±0.02), SC4 ratio 0.339–0.51 (0.438±0.04), Coxa I setae = tapered/medium tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Apache: 0.75 miles NE Hwy-191 on Hwy-61, 34.908575 -109.2273411, 6251ft., [APH_1408, 9/10/2011, 1♂, Brent E. Hendrixson, Thomas Martin, AUMNH]; 9.5 miles N Hwy-61 on Hwy-191, 35.030934 -109.2545851, 6415ft., [APH_1423, 12/10/2011, 1 juv, Brent E. Hendrixson, Thomas Martin, AUMNH]; Ganada, 35.711279 -109.5420495, 6398ft., [APH_2576, 5/10/1962, 1♂, Charles Supplee,
Distribution and natural history
Aphonopelma marxi has a wide distribution across higher elevation areas of northern Arizona (Fig.

Conservation status
Aphonopelma marxi has a wide distribution across higher elevation habitats centered on the Four Corners region of Arizona, Colorado, New Mexico, and Utah. Some populations appear to be restricted to isolated mountains (e.g., Mount Ord) but are otherwise very common (but can be quite difficult to find due to the cryptic nature of their burrows). The species is likely secure.
Remarks
Because no labeled types existed,
Other important ratios that distinguish males: A. marxi possess a larger T1/M1 (≥1.39; 1.39–1.52) than A. chalcodes (≤1.13; 0.99–1.13), A. hentzi (≤1.24; 1.08–1.24), A. mareki (≤1.33; 1.26–1.33), A. vorhiesi (≤1.36; 1.09–1.36); by possessing a larger PTl/M3 (≥0.92; 0.92–1.16) than A. chalcodes (≤0.76; 0.65–0.76), A. hentzi (≤0.81; 0.71–0.81), A. mareki (≤0.92; 0.76–0.92), and A. vorhiesi (≤0.87; 0.71–0.87). Other important ratios that distinguish females: A. marxi possess a larger L1/L3 (≥1.17; 1.17–1.21) than A. hentzi (≤1.15; 1.07–1.15); by possessing a larger T1/T4 (≥1.06; 1.06–1.14) than A. hentzi (≤1.05; 0.85–1.05), A. mareki (≤1.01; 0.94–1.01), and A. vorhiesi (≤0.98; 0.93–0.98); by possessing a larger L4 scopulation extent (33%-51%) than A. mareki (21%-27%) and smaller than A. chalcodes (56%-81%). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Aphonopelma moderatum
Delopelma moderatum Chamberlin & Ivie, 1939: 9; male holotype from 5 miles E of Rio Grande City, Starr Co., Texas, 26.342436 -98.7287495, elev. 231ft., 1.v.1937, coll. S. Mulaik; deposited in
Rhechostica moderatum Raven, 1985: 149.
Aphonopelma moderatum Smith, 1995: 122.
Aphonopelma heterops Chamberlin, 1940: 29; female syntypes from Edinburg, Hidalgo, Co., Texas, 26.30173 -98.163355, elev. 96ft., ix-xii.1933, coll. S. Mulaik; deposited in the
Diagnosis
Aphonopelma moderatum (Fig.

Description
Male originally described by
Redescription of male exemplar
(APH_0890; Fig.

Aphonopelma moderatum (Chamberlin & Ivie, 1939). A–I male specimen, APH_0890 A dorsal view of carapace, scale bar = 5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4.5mm E ventral view of metatarsus IV, scale bar = 5.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 3mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 3.5mm.
Variation (9).Cl 11.46–16.06 (14.409±0.52), Cw 9.49–14.51 (12.722±0.57), LBl 1.52–1.99 (1.811±0.05), LBw 1.7–2.36 (2.019±0.09), F1 12.35–17.39 (15.218±0.51), F1w 2.66–3.84 (3.453±0.12), P1 4.99–7.03 (6.133±0.23), T1 10.23–14.28 (12.43±0.43), M1 10.1–13.38 (11.754±0.35), A1 6.29–7.76 (6.957±0.18), L1 length 43.96–59.84 (52.491±1.65), F3 10.11–13.82 (12.24±0.43), F3w 2.88–4.21 (3.611±0.14), P3 4.17–6.13 (5±0.25), T3 7.62–10.88 (9.55±0.41), M3 9.45–13.75 (11.829±0.44), A3 5.36–7.51 (6.729±0.24), L3 length 36.71–52.05 (45.348±1.72), F4 11.54–16.23 (14.024±0.47), F4w 2.67–3.84 (3.347±0.14), P4 4.27–6.10 (5.234±0.24), T4 9.97–13.53 (12.232±0.38), M4 12.82–17.54 (15.129±0.52), A4 6.62–8.18 (7.478±0.21), L4 length 45.64–61.46 (54.098±1.77), PTl 6.258–8.692 (7.7±0.23), PTw 2.033–2.70 (2.441±0.07), SC3 ratio 0.481–0.782 (0.646±0.03), SC4 ratio 0.313–0.503 (0.411±0.02), Coxa 1 setae = tapered/thin tapered, F3 condition = normal/slightly swollen.
Description of female exemplar
(APH_0532; Figs

Aphonopelma moderatum (Chamberlin & Ivie, 1939). A–E female specimen, APH_0532 A dorsal view of carapace, scale bar = 6.5mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 3.5mm D ventral view of metatarsus IV, scale bar = 3.5mm E prolateral view of L pedipalp and palpal tibia.
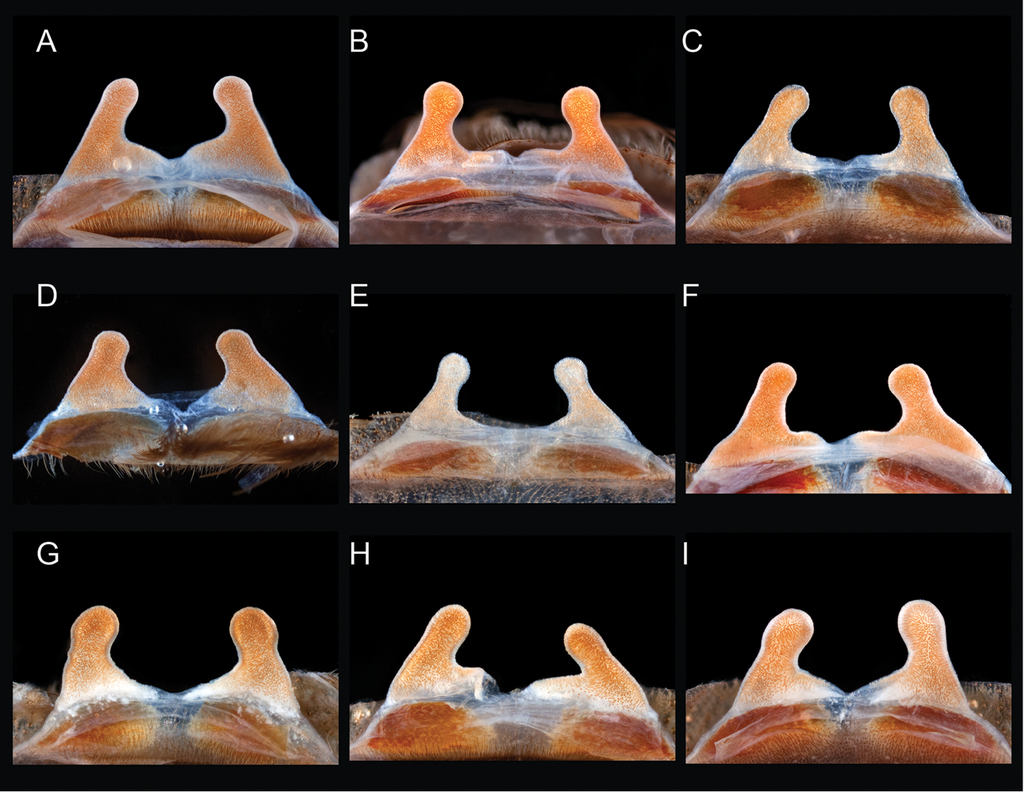
Variation (9).Cl 9.98–15.62 (12.719±0.67), Cw 8.33–13.47 (10.64±0.61), LBl 1.37–2.23 (1.701±0.1), LBw 1.53–2.37 (1.914±0.1), F1 7.936–13.14 (10.457±0.6), F1w 2.497–3.98 (3.17±0.17), P1 3.662–6.09 (4.812±0.28), T1 6.485–10.17 (8.135±0.4), M1 4.719–8.26 (6.4±0.4), A1 3.974–6.27 (5.317±0.25), L1 length 26.776–43.44 (35.122±1.9), F3 5.974–10.15 (8.053±0.51), F3w 2.164–3.62 (2.766±0.19), P3 2.982–5.45 (3.686±0.33), T3 4.486–7.38 (5.541±0.35), M3 5.15–8.57 (6.447±0.45), A3 4.086–6.41 (5.171±0.32), L3 length 22.678–37.96 (28.898±1.92), F4 7.761–12.66 (10.041±0.54), F4w 2.287–3.66 (2.976±0.16), P4 3.37–5.76 (4.291±0.29), T4 6.417–10.42 (8.373±0.43), M4 7.353–11.91 (9.486±0.52), A4 4.808–6.91 (5.836±0.26), L4 length 29.709–47.28 (38.028±2.01), SC3 ratio 0.657–0.736 (0.699±0.01), SC4 ratio 0.334–0.486 (0.413±0.02), Coxa 1 setae = tapered/thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Texas: Dimmit: Picnic area 3 miles NW Catarina on US-83, 28.375443 -99.648111, 566ft., [APH_0471-0472, 11/4/2009, 2 juv, Brent E. Hendrixson, AUMNH]; Hidalgo: Edinberg, 26.290053 -98.1550735, 92ft., [APH_2208, 12/1937, 1♀, S. Mulaik,
Distribution and natural history
Aphonopelma moderatum is widely distributed in counties bordering the Rio Grande in Texas from Langtry (Val Verde County) to Edinburg (Hidalgo County) to more interior areas such as McMullen County (Figs

Conservation status
The type specimens of A. heterops (considered a synonym of A. moderatum) were collected from Edinburg (Hidalgo County) in 1933. In the 80+ years since their collection, much of the land in this section of the Lower Rio Grande Valley has been converted for agricultural use. It is no surprise then that we failed to find these spiders near Edinburg despite traveling to the area numerous times. Consequently, A. moderatum may be extirpated from the Edinburg-McAllen-Mission Metropolitan area, but these spiders may have never been particularly abundant in these parts in the first place given that the location is near the easternmost portion of their range. These tarantulas are among the most attractive species in the United States (Fig.
Remarks
Aphonopelma moderatum exhibits considerable sexual dimorphism in coloration. Adult females and immature specimens of both sexes possess perhaps the most distinctive color pattern of any species in the United States (i.e., tan to orange body with distinctly darker leg patellae, metatarsi, and tarsi). Adult males, on the other hand, are often solid black or dark brown. We suspect that
Mitochondrial DNA (CO1) identifies A. moderatum as a polyphyletic species with two divergent lineages spread across the tree (Fig.
Other important ratios that distinguish males: A. moderatum possess a larger L1/L4 (≥0.95; 0.95–0.99) than A. gabeli (≤0.95; 0.90–0.95). For both males and females, certain morphometrics have potential to be useful but due to the amounts of variation, small number of specimens, and/or the small differences between species no others are claimed to be significant at this time (see Suppl. material
Aphonopelma moellendorfi , sp. n.
Types
Male holotype (APH_0925) from N of Del Rio on Hwy 277, Val Verde Co., Texas, 29.488717 -100.9076335, elev. 1190ft., 2006, coll. Dave Moellendorf; deposited in AUMNH. Paratype male (APH_0928) from Del Rio - Hwy 90 W of Lake Amistad, Val Verde Co., Texas, 29.618868 -101.1043855, elev. 1406ft., 2006, coll. Dave Moellendorf; deposited in
Etymology
The specific epithet is a patronym in recognition of Dave Moellendorf, a friend and mentor to CAH. Moellendorf introduced CAH to the tarantula fauna of Texas and encouraged and fostered CAH’s early research on the genus Aphonopelma. Moellendorf has also spent countless hours educating the public on the importance of spiders on our planet.
Diagnosis
Aphonopelma moellendorfi belongs to the Moderatum species complex and can be distinguished by a combination of morphological, molecular, and geographic characteristics. Nuclear and mitochondrial DNA identifies A. moellendorfi as a phylogenetically distinct monophyletic lineage (Figs
Description of male holotype
(APH_0925; Fig.

Aphonopelma moellendorfi sp. n. A–I male holotype, APH_0925 A dorsal view of carapace, scale bar = 6mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4.5mm E ventral view of metatarsus IV, scale bar = 3mm F prolateral view of L pedipalp and palpal tibia, scale bar = 4mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 5.5mm.
Variation (5).Cl 15.05–17.56 (16.192±0.51), Cw 13.27–15.48 (14.308±0.39), LBl 1.65–2.04 (1.9±0.07), LBw 1.84–2.28 (2.124±0.09), F1 16.36–18.10 (17.338±0.32), F1w 3.42–4.15 (3.81±0.14), P1 6.31–7.25 (6.698±0.17), T1 13.57–15.75 (14.396±0.41), M1 13.11–15.27 (13.814±0.42), A1 7.68–8.49 (8.126±0.14), L1 length 57.15–64.29 (60.372±1.25), F3 13.56–15.47 (14.28±0.34), F3w 3.7–4.52 (4.034±0.14), P3 4.84–5.79 (5.36±0.18), T3 10.02–11.79 (10.972±0.38), M3 13.11–15.48 (14.032±0.45), A3 7.04–8.47 (7.678±0.24), L3 length 49.71–56.88 (52.322±1.34), F4 17.29–15.38 (16.23±0.31), F4w 4.35–3.51 (3.846±0.15), P4 6.02–5.14 (5.536±0.17), T4 12.52–14.12 (13.38±0.31), M4 16.04–18.88 (17.518±0.54), A4 8.14–9.17 (8.616±0.17), L4 length 58.64–64.39 (61.28±1.15), PTl 8.6–9.175 (8.898±0.12), PTw 2.051–2.67 (2.418±0.11), SC3 ratio 0.571–0.785 (0.674±0.03), SC4 ratio 0.341–0.481 (0.409±0.03), Coxa I setae = thin tapered, F3 condition = normal.
Material examined
United States: Texas: Presidio: Hwy 169, 29.938343 -104.0340116, 3992ft., [APH_0948, 2006, 1♂, Dave Moellendorf, AUMNH]; Sierra Vieja Mtns, 13 miles west Valentine, 30.723313 -104.6693914, 4127ft., [APH_0938, 2006, 1♂, Dave Moellendorf, AUMNH]; [APH_0969, 2006, 1♂, Dave Moellendorf, AUMNH]; Val Verde: Del Rio - Hwy 90 W of Lake Amistad, 29.618868 -101.1043855, 1406ft., [APH_0928, 2006, 1♂, Dave Moellendorf,
Distribution and natural history
Aphonopelma moellendorfi is presently known from only a handful of localities in southwestern Texas, along the border with Mexico from Del Rio west to the Sierra Vieja Mountains (Fig.

Conservation status
This enigmatic species is only known from a handful of localities in the United States so it would be premature to comment on its conservation status. Fieldwork in West Texas and northern Mexico is necessary to more fully understand the abundance and extent of this species.
Remarks
Aphonopelma moellendorfi males are morphologically similar to A. gabeli and A. moderatum, and lack the characteristic shiny copper or bronze carapace found in A. hentzi. But interestingly, while females of A. moellendorfi remain unknown, it should be noted that immature males closely resemble A. hentzi (D. Moellendorf, pers. comm., collected and raised immature males to maturity). Other important ratios that distinguish males: A. moellendorfi possess a larger L1/L4 (≥0.96; 0.96–1.00) than A. gabeli (≤0.95; 0.90–0.95) and A. anax (≤0.94; 0.88–0.94); by possessing a larger M1/F4 (≥0.81; 0.81–0.88) than A. anax (≤0.75; 0.69–0.75) and A. hentzi (≤0.77; 0.68–0.77). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma mojave
Aphonopelma mojave Prentice, 1997: 157; male holotype and female allotype from S of Red Mountain, 20 miles N of Kramer Jct. on Hwy 395, 1–2 miles W of Hwy 395, San Bernardino and Kern Co. line, California, 35.310573 -117.6443504, elev. 3405ft., 28.x.1989 (male), 20.x.1991 (female), coll. Thomas R. Prentice; deposited in
Diagnosis
Aphonopelma mojave (Fig.

Description
Male and female originally described by
Redescription of male exemplar
(APH_1561; Fig.

Aphonopelma mojave Prentice, 1997. A–I male specimen, APH_1561 A dorsal view of carapace, scale bar = 2.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2mm E ventral view of metatarsus IV, scale bar = 2mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 3mm.
Variation (6).Cl 7.374–8.988 (7.862±0.25), Cw 6.689–8.035 (7.185±0.2), LBl 1.03–1.206 (1.079±0.03), LBw 1.22–1.371 (1.272±0.02), F1 7.352–9.266 (8.251±0.25), F1w 1.764–2.218 (1.908±0.07), P1 2.873–3.421 (3.16±0.08), T1 6.69–8.041 (7.323±0.19), M1 6.01–7.277 (6.492±0.17), A1 3.87–4.994 (4.451±0.17), L1 length 27.868–32.999 (29.677±0.76), F3 6.577–7.938 (7.23±0.19), F3w 1.915–2.449 (2.081±0.08), P3 2.537–2.931 (2.686±0.07), T3 5.162–6.539 (5.683±0.19), M3 6.462–7.865 (7.072±0.2), A3 4.293–5.387 (4.685±0.17), L3 length 25.19–30.66 (27.355±0.77), F4 8.283–9.518 (8.682±0.18), F4w 1.754–2.14 (1.874±0.06), P4 2.468–3.188 (2.791±0.11), T4 6.99–8.241 (7.506±0.17), M4 8.358–9.946 (8.998±0.23), A4 4.446–5.929 (4.984±0.24), L4 length 30.958–36.657 (32.961±0.83), PTl 4.866–5.584 (5.153±0.1), PTw 1.536–2.029 (1.75±0.08), SC3 ratio 0.601–0.792 (0.666±0.03), SC4 ratio 0.303–0.438 (0.377±0.02), Coxa I setae = very thin tapered, F3 condition = normal.
Description of female exemplar
(APH_1558; Figs

Aphonopelma mojave Prentice, 1997. A–E female specimen, APH_1558 A dorsal view of carapace, scale bar = 3mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 2.5mm D ventral view of metatarsus IV, scale bar = 2.5mm E prolateral view of L pedipalp and palpal tibia.

Variation (7).Cl 6.911–8.471 (7.803±0.21), Cw 6.203–7.648 (6.858±0.2), LBl 1.071–1.48 (1.24±0.05), LBw 1.263–1.70 (1.453±0.06), F1 5.84–7.52 (6.877±0.21), F1w 1.66–1.962 (1.873±0.04), P1 2.15–3.163 (2.774±0.12), T1 4.72–6.53 (5.642±0.21), M1 3.73–4.66 (4.242±0.12), A1 3.16–4.18 (3.588±0.12), L1 length 19.6–25.75 (23.122±0.72), F3 5.311–6.34 (5.747±0.13), F3w 1.37–1.849 (1.666±0.07), P3 2.06–2.774 (2.426±0.1), T3 3.38–4.76 (4.153±0.17), M3 4.05–4.87 (4.477±0.12), A3 3.08–4.218 (3.737±0.13), L3 length 18.14–22.45 (20.54±0.56), F4 6.68–7.76 (7.196±0.14), F4w 1.29–1.942 (1.771±0.09), P4 2.433–2.93 (2.673±0.08), T4 5.37–6.26 (5.767±0.12), M4 5.68–6.65 (6.192±0.14), A4 3.56–4.753 (4.238±0.15), L4 length 23.73–28.17 (26.067±0.56), SC3 ratio 0.653–0.768 (0.706±0.02), SC4 ratio 0.343–0.525 (0.407±0.03), Coxa I setae = very thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: California: Kern: 7 miles north of Johannesburg, 35.4804 -117.6546835, 3412ft., [APH_2669, 9/10/1968, 1♂, W. Icenogle,
Distribution and natural history
The redefined A. mojave has a narrow distribution centered on the western Mojave Desert and extreme southeastern foothills of the Sierra Nevada Mountains in eastern Kern and northwestern San Bernardino Counties, California (Fig.

Conservation status
In addition to having a relatively narrow distribution, habitat degradation due to Off-Highway Vehicle (OHV) recreational activity probably poses the greatest threat to A. mojave. However, these spiders are abundant (especially near the type locality), with several areas within the western Mojave Desert that are managed by the State of California or Bureau of Land Management (e.g., West Mojave Desert Ecological Reserve, Fremont Valley Ecological Reserve, Red Rock Canyon State Park, Desert Tortoise Natural Area). This species is likely secure.
Remarks
Aphonopelma paloma
Aphonopelma paloma Prentice, 1993: 189; male holotype and female allotype from 3 miles NE exit 151 off I-8 (jct with Hwy 84), Pinal Co., Arizona, 32.856167 -112.0866092, elev. 4310ft., 17-18.xi.1989, coll. Tom Prentice; deposited in
Apachepelma paloma Smith, 1995: 45.
Aphonopelma paloma Prentice, 1997: 140.
Diagnosis
Aphonopelma paloma (Fig.


Description
Male and female originally described by
Redescription of male exemplar
(APH_1603; Fig.

Aphonopelma paloma Prentice, 1993. A–I male specimen, APH_1603 A dorsal view of carapace, scale bar = 2mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 1.5mm E ventral view of metatarsus IV, scale bar = 2mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 1.5mm.
Variation (6).Cl 5.126–5.502 (5.343±0.06), Cw 4.468–5.124 (4.824±0.1), LBl 0.675–0.797 (0.741±0.02), LBw 0.868–0.898 (0.878±0), F1 5.289–6.208 (5.723±0.15), F1w 1.176–1.402 (1.27±0.04), P1 1.741–2.44 (2.047±0.1), T1 4.034–5.248 (4.839±0.18), M1 3.62–4.297 (4.017±0.11), A1 2.785–2.876 (2.827±0.01), L1 length 18.374–20.949 (19.453±0.47), F3 4.466–4.958 (4.737±0.08), F3w 1.456–1.831 (1.673±0.06), P3 1.528–1.804 (1.694±0.04), T3 3.244–3.795 (3.532±0.09), M3 4.019–4.569 (4.332±0.09), A3 2.658–3.047 (2.837±0.06), L3 length 15.988–17.891 (17.131±0.33), F4 5.308–5.984 (5.703±0.11), F4w 1.112–1.445 (1.313±0.05), P4 1.532–2.102 (1.829±0.08), T4 4.578–5.096 (4.847±0.08), M4 5.412–6.102 (5.71±0.13), A4 3.042–3.374 (3.28±0.05), L4 length 20.357–22.272 (21.37±0.4), PTl 3.273–3.706 (3.53±0.07), PTw 1.159–1.431 (1.263±0.04), SC3 ratio 0.219–0.41 (0.309±0.03), SC4 ratio 0.056–0.245 (0.12±0.03), Coxa I setae = very thin tapered, F3 condition = swollen/slightly swollen.
Description of female exemplar
(APH_1255; Figs



Variation (12).Cl 4.635–7.058 (5.66±0.24), Cw 4.192–6.092 (5.023±0.21), LBl 0.763–1.197 (0.904±0.04), LBw 0.903–1.346 (1.074±0.04), F1 3.597–5.573 (4.52±0.19), F1w 1.186–1.884 (1.508±0.07), P1 1.494–2.461 (1.971±0.09), T1 2.968–4.516 (3.726±0.14), M1 1.911–3.189 (2.499±0.12), A1 1.927–2.705 (2.295±0.08), L1 length 11.984–18.261 (15.01±0.61), F3 2.973–4.507 (3.635±0.16), F3w 1.037–1.809 (1.346±0.07), P3 1.318–2.141 (1.687±0.08), T3 2.035–3.057 (2.523±0.1), M3 2.067–3.361 (2.607±0.13), A3 2.031–2.976 (2.401±0.09), L3 length 10.438–15.884 (12.853±0.55), F4 3.883–5.675 (4.735±0.18), F4w 1.088–1.864 (1.374±0.07), P4 1.521–2.33 (1.911±0.09), T4 3.135–4.529 (3.799±0.14), M4 3.083–4.612 (3.718±0.16), A4 2.308–3.263 (2.741±0.1), L4 length 13.93–20.232 (16.904±0.65), SC3 ratio 0.26–0.496 (0.392±0.02), SC4 ratio 0–0.247 (0.169±0.02), Coxa I setae = very thin tapered. Spermathecae variation can be seen in Figures
Material examined
United States: Arizona: Maricopa: 1.2 miles S Aqueduct on Aguila-Wickenburg Rd, 33.571504 -112.8181081, 1306ft., [APH_0432, 16/11/2008, 1 juv, Brent E. Hendrixson, AUMNH]; 4.65 miles N I-8 (Sentinel) on Agua Caliente Rd, 32.908339 -113.2621791, 584ft., [APH_0430, 16/11/2008, 1 juv, Brent E. Hendrixson, AUMNH]; just NW of Jct. Vulture Mine Rd and Aguila Rd, 33.719699 -112.88241, 1716ft., [APH_1612-1613, 12/11/2012, 2♂, Brent E. Hendrixson, AUMNH]; just S of White Tank Mtn Rd, 33.56397 -112.491472, 1360ft., [APH_1101-1103, 30/11/2009, 3 juv, June Olberding, AUMNH]; Pima: 0.75 miles S Ball Rd in Why along Hwy-85, 32.25235 -112.7452061, 1765ft., [APH_0424, 15/11/2008, 1 juv, Brent E. Hendrixson, AUMNH]; [APH_0426-0427, 15/11/2008, 1♀, 1♂, Brent E. Hendrixson, AUMNH]; Bates Well Rd, S of Ajo, 32.27002 -112.860872, 1673ft., [APH_1114-1116, 15/12/2009, 2♀, 1♂, June Olberding, AUMNH]; in desert east of Bates Well Rd, about 7 miles W of Why, AZ, 32.28612 -112.854354, 1720ft., [APH_0442-0449, 12/2008, 3♀, 5 juv, June Olberding, AUMNH]; Tucson area, near Catalina State Park, N side of Tangerine Rd and E of Tangerine Crossing, E of Marana, 32.42413 -111.034271, 2631ft., [APH_3170-3172, 12/11/2013, 3♀, Chris A. Hamilton, Brent E. Hendrixson, AUMNH]; W of Why, 32.26683 -112.8472, 1650ft., [APH_1253-1258, 1/12/2010, 2♂, 4♀, June Olberding, AUMNH]; Saguaro National Park (West) - western portion, 32.32247 -111.126082, 2790ft., [APH_1675, 1/12/14, 1♂, Derrick Smith, AUMNH; Pinal: 0.7 miles S Skyline Dr. on Quail Run Ln, 33.180864 -111.4965162, 1552ft., [APH_1671-1674, 6/12/2012, 4♀, June Olberding, Tim Cota, AUMNH]; in desert NW jct of Hwy-84 and Amarillo Valley Rd (3.2 miles NW I-8), 32.856511 -112.0855641, 1541ft., [APH_0421-0423, 15/11/2008, 3 juv, Brent E. Hendrixson, Paul Marek, Charity Hall, Kojun, AUMNH]; [APH_1603-1606, 10/11/2012, 3♂, 1♀, Brent E. Hendrixson, AUMNH]; off Florence-Kelvin Hwy, BLM land E of Florence, 32.99957 -111.264841, 1968ft., [APH_3195, 15/11/2013, 1♀, Chris A. Hamilton, Brent E. Hendrixson, AUMNH]; [APH_3197-3198, 15/11/2013, 2♀, Chris A. Hamilton, Brent E. Hendrixson, AUMNH]; off Hwy 79 around 1/2 way between 77 split and Florence at Tom Mix Rest Area, 32.818271 -111.2047511, 2338ft., [APH_3194, 15/11/2013, 1♀, Brent E. Hendrixson, Chris A. Hamilton, AUMNH]; off Hwy 79, E side of road, N of Tucson past Hwy 77 split, near Ninety Six hills, 32.682347 -111.0725311, 2972ft., [APH_3189-3190, 14/11/2013, 2♀, Chris A. Hamilton, Brent E. Hendrixson, AUMNH]; W of Chuck’s Corner off S. Amarillo Valley Rd, off Hwy 84, N of I-8, 32.85665 -112.084191, 1501ft., [APH_3166, 11/11/2013, 1♀, Brent E. Hendrixson, Chris A. Hamilton, AUMNH]; [APH_3168, 11/11/2013, 1 juv, Chris A. Hamilton, Brent E. Hendrixson, AUMNH].
Distribution and natural history
Aphonopelma paloma is widely distributed across lower elevation sections of southern and southwestern Arizona (Figs

Conservation status
Aphonopelma paloma is abundant throughout its distribution but can be difficult to find due to the cryptic nature of their burrows and narrow window of activity during the year. This species is secure.
Remarks
Other important ratios that distinguish Aphonopelma paloma males: possess a smaller L3 scopulation extent (21%–41%) than A. parvum (60%–65%) and A. xwalxwal (65%–95%); by possessing a larger PTl/M1 (≥0.81; 0.81–0.92) than A. chalcodes (≤0.75; 0.67–0.75) and A. xwalxwal (≤0.64; 0.57–0.64); by possessing a larger M3/A3 (≥1.46; 1.46–1.61) than A. mareki (≤1.42; 1.27–1.42) and A. parvum (≤1.41; 1.21–1.41), but smaller than A. xwalxwal (≥1.66; 1.66–1.91); by possessing a larger Cl/M3 (≥1.17; 1.17–1.31) than A. superstitionense (≤1.12; 1.05–1.12) and A. xwalxwal (≤1.01; 0.91–1.01). Other important ratios that distinguish females: A. paloma possess a larger Cl/A4 (≥1.92; 1.92–2.21) than A. parvum (≤1.91; 1.63–1.91), but smaller than A. chalcodes (≥2.32; 2.32–2.64); by possessing a smaller L3 scopulation extent (25%–50%) than A. chalcodes (78%-93%), A. parvum (62–67%), A. superstitionense (64% ± (only 1 specimen)), and A. vorhiesi (49%–69%, with slight overlap). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Aphonopelma parvum , sp. n.
Types
Male holotype (APH_1264) collected along CR-C078, Hidalgo Co., New Mexico, 32.0679 -108.946082, elev. 4296ft., 6.xii.2010, coll. June Olberding; deposited in AUMNH. Paratype female (APH_1622) from 0.2 miles N Hwy-533 (Portal Rd) on Stateline Rd., Hidalgo Co., New Mexico, 31.874121 -109.0473771, elev. 4107ft., 14.xi.2012, coll. Brent E. Hendrixson; deposited in AUMNH. Paratype male (APH_1106) from 0.11 miles N Portal Rd (Hwy 533) on State Line Rd, Hidalgo Co., New Mexico, 31.87232 -109.047872, elev. 4127ft., 28.xi.2009, coll. June Olberding; deposited in
Etymology
The specific epithet is a neuter adjective for “very small” or “very little” and is in reference to the diminutive size of this new tarantula species.
Diagnosis
Aphonopelma parvum (Fig.

Description of male holotype
(APH_1264; Fig.

Aphonopelma parvum sp. n. A–I male holotype, APH_1264 A dorsal view of carapace, scale bar = 3mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2mm E ventral view of metatarsus IV, scale bar = 2.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 3mm.
Variation (7).Cl 5.702–7.057 (6.333±0.19), Cw 5.141–6.376 (5.836±0.2), LBl 0.764–0.889 (0.829±0.02), LBw 0.888–0.989 (0.958±0.01), F1 6.212–7.966 (7.063±0.25), F1w 1.312–1.704 (1.54±0.05), P1 1.967–2.597 (2.35±0.09), T1 5.269–6.919 (6.012±0.21), M1 3.837–4.863 (4.483±0.16), A1 3.282–4.182 (3.593±0.13), L1 length 20.633–26.355 (23.501±0.76), F3 4.964–6.228 (5.631±0.2), F3w 1.305–1.865 (1.637±0.08), P3 1.806–2.173 (2.025±0.05), T3 3.791–4.869 (4.318±0.14), M3 4.408–5.41 (4.817±0.14), A3 3.141–4.143 (3.758±0.13), L3 length 18.197–22.807 (20.548±0.63), F4 6.055–7.436 (6.741±0.2), F4w 1.226–1.634 (1.453±0.06), P4 2.085–2.476 (2.214±0.05), T4 5.104–6.65 (5.772±0.2), M4 5.78–6.86 (6.336±0.18), A4 3.915–4.963 (4.374±0.13), L4 length 23.105–28.206 (25.438±0.71), PTl 3.619–4.618 (4.139±0.13), PTw 1.377–1.673 (1.567±0.05), SC3 ratio 0.601–0.649 (0.63±0.01), SC4 ratio 0.368–0.442 (0.399±0.01), Coxa I setae = very thin tapered, F3 condition = normal.
Description of female paratype
(APH_1622; Figs


Variation (5).Cl 6.181–7.326 (6.974±0.21), Cw 5.827–6.86 (6.28±0.2), LBl 0.964–1.136 (1.074±0.03), LBw 1.072–1.295 (1.206±0.04), F1 5.344–6.154 (5.849±0.14), F1w 1.777–2.043 (1.896±0.05), P1 2.291–2.693 (2.453±0.07), T1 4.492–5.026 (4.721±0.1), M1 2.927–3.487 (3.099±0.1), A1 2.777–3.283 (3.027±0.09), L1 length 18.064–20.322 (19.149±0.4), F3 4.621–5.077 (4.851±0.1), F3w 1.515–1.742 (1.646±0.04), P3 2.127–2.267 (2.168±0.03), T3 3.192–3.467 (3.369±0.05), M3 3.221–3.703 (3.44±0.09), A3 3.027–3.573 (3.227±0.1), L3 length 16.258–17.782 (17.056±0.29), F4 5.735–6.537 (6.147±0.15), F4w 1.598–1.764 (1.696±0.03), P4 2.255–2.544 (2.383±0.05), T4 4.539–5.438 (4.98±0.16), M4 4.592–5.153 (4.942±0.1), A4 3.603–4.035 (3.886±0.08), L4 length 21.215–23.343 (22.339±0.46), SC3 ratio 0.629–0.672 (0.651±0.01), SC4 ratio 0.338–0.422 (0.397±0.02), Coxa I setae = very thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Cochise: 0.8 miles SE Hwy-191 on connecting road, 32.384654 -109.6542691, 4231ft., [APH_1619, 14/11/2012, 1♀, Brent E. Hendrixson, AUMNH]; Cave Creek, 1 mile south of Ranger Station, 31.888981 -109.168954, 5067ft., [AUMS_2247, 28/11/1990, 1♂, Barney Tomberlin, AUMNH]; on Portal Rd going towards Cave Creek Canyon, 31.91472 -109.1462621, 4763ft., [APH_3187, 13/11/2013, 1♂, Chris A. Hamilton, Brent E. Hendrixson, AUMNH]; on Portal Rd, just past the state line, 31.87769 -109.0605871, 4195ft., [APH_3186, 13/11/2013, 1♂, Chris A. Hamilton, Brent E. Hendrixson, AUMNH]; Portal, 31.913703 -109.141455, 4770ft., [APH_2209, 20/11/1963, 1♂, V. Roth,
Distribution and natural history
Aphonopelma parvum can be found inhabiting the Chihuahuan Desert and Madrean Archipelago Level III Ecoregions of southeastern Arizona and southwestern New Mexico (Figs

Conservation status
Aphonopelma parvum is very common throughout their distribution but can be difficult to find due to the cryptic nature of their burrows and narrow window of activity during the year. This species is likely secure.
Remarks
Other important ratios that distinguish males: A. parvum possess a larger F1/M3 (≥1.40; 1.40–1.53) than A. chalcodes (≤1.31; 1.13–1.31), A. gabeli (≤1.24; 1.18–1.24), A. hentzi (≤1.40; 1.20–1.40), A. paloma (≤1.38; 1.24–1.38), A. saguaro (≤1.37; 1.22–1.37), and A. superstitionense (≤1.33; 1.22–1.33). Other important ratios that distinguish A. parvum females: A. parvum possess a larger F3L/W (≥2.85; 2.85–3.05) than A. saguaro (2.41 ± (only 1 specimen)); a smaller P1/F3 (≤0.54; 0.47–0.54) than A. chiricahua (0.69 ± (only 1 specimen)) and A. superstitionense (0.58 ± (only 1 specimen)); a smaller Cl/A4 (≤1.91; 1.63–1.91) than A. paloma (≥1.92; 1.92–2.21); a smaller Cl/F4 (≤1.17; 1.07–1.17) than A. gabeli (≥1.20; 1.20–1.32), A. hentzi (≥1.20; 1.20–1.40), and A. vorhiesi (≥1.23; 1.23–1.37); a smaller L4 scopulation extent than A. chalcodes (56%–81%). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma peloncillo , sp. n.
Types
Male holotype (APH_0672) from along Hwy-338, Hidalgo Co., New Mexico, 31.607079 -108.8670811, elev. 4934ft., 14.vii.2009, coll. Brent E. Hendrixson and Nate Davis; deposited in AUMNH. Paratype female (APH_1296) from Coronado National Forest, Peloncillo Mtns, Clanton Draw Area, Hidalgo Co., New Mexico, 31.518665 -108.9825971, elev. 5435ft., 25.vii.2011, coll. Brent E. Hendrixson, Brendon Barnes, Nate Davis, Jake Storms; deposited in AUMNH. Paratype male (APH_1190) from along Hwy-338, Hidalgo Co., New Mexico, 31.607079 -108.8670811, elev. 4934ft., 26.vii.2010, coll. Brent E. Hendrixson, Brendon Barnes, Nate Davis; deposited in
Etymology
The specific epithet is a noun in apposition taken from type locality, the Peloncillo Mountains in southwestern New Mexico, where this species was first discovered.
Diagnosis
Aphonopelma peloncillo (Fig.

Description of male holotype
(APH_0672; Fig.

Aphonopelma peloncillo sp. n. A–I male holotype, APH_0672 A dorsal view of carapace, scale bar = 4mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 3.5mm E ventral view of metatarsus IV, scale bar = 4mm F prolateral view of L pedipalp and palpal tibia, scale bar = 5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 4mm.
Variation (9).Cl 10.23–14.90 (12.567±0.44), Cw 9.15–14.10 (11.582±0.46), LBl 1.32–1.95 (1.46±0.06), LBw 1.4–2.07 (1.651±0.07), F1 11.148–14.86 (13.352±0.4), F1w 2.423–3.795 (3.059±0.12), P1 4.151–5.594 (4.945±0.18), T1 9.699–12.83 (11.482±0.36), M1 7.331–10.436 (8.77±0.29), A1 5.179–6.889 (6.104±0.18), L1 length 37.508–50.297 (44.654±1.31), F3 9.27–12.369 (10.739±0.32), F3w 2.766–4.012 (3.286±0.12), P3 3.629–4.891 (4.129±0.14), T3 6.674–9.573 (8.172±0.3), M3 7.465–10.86 (9.588±0.34), A3 5.712–7.983 (6.445±0.25), L3 length 33.029–45.454 (39.073±1.25), F4 10.549–14.471 (12.599±0.38), F4w 2.469–3.765 (2.996±0.12), P4 3.44–4.991 (4.353±0.16), T4 9.249–12.388 (10.832±0.35), M4 10.683–14.17 (12.768±0.4), A4 6.65–8.656 (7.43±0.24), L4 length 40.656–54.24 (47.981±1.42), PTl 6.119–8.133 (7.382±0.24), PTw 2.027–2.91 (2.507±0.1), SC3 ratio 0.526–0.685 (0.594±0.02), SC4 ratio 0.274–0.442 (0.338±0.02), Coxa 1 setae = tapered/thin tapered, F3 condition = normal.
Description of female paratype
(APH_1296; Figs


Variation (5).Cl 15.33–19.32 (16.912±0.69), Cw 13.1–16.76 (14.756±0.65), LBl 1.74–2.33 (1.992±0.1), LBw 2.02–2.65 (2.224±0.11), F1 12.25–14.51 (13.252±0.43), F1w 3.81–4.60 (4.086±0.14), P1 5.3–6.72 (5.9±0.28), T1 9.47–10.97 (10.376±0.27), M1 6.68–8.45 (7.686±0.31), A1 5.95–6.34 (6.2±0.07), L1 length 39.65–46.93 (43.414±1.28), F3 9.48–11.88 (10.484±0.43), F3w 3.14–3.98 (3.51±0.14), P3 4.54–5.73 (4.956±0.21), T3 7.01–8.62 (7.748±0.31), M3 7.51–8.94 (8.262±0.3), A3 5.7–6.68 (6.234±0.17), L3 length 34.5–41.85 (37.684±1.26), F4 12.26–14.42 (13.224±0.41), F4w 3.5–4.13 (3.718±0.11), P4 5.09–6.40 (5.444±0.25), T4 9.66–11.27 (10.602±0.26), M4 9.91–12.05 (11.294±0.39), A4 6.63–7.56 (7.118±0.18), L4 length 43.84–51.26 (47.682±1.31), SC3 ratio 0.586–0.676 (0.64±0.02), SC4 ratio 0.324–0.386 (0.357±0.01), Coxa I setae = tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Cochise: 0.03 miles N Hunt Canyon Rd on Leslie Canyon Rd, 31.64689064 -109.47322912, 4927ft., [APH_0718, 17/8/2009, 1♂, Alice Abela, AUMNH]; 0.51 miles N Davis Ranch Rd on Leslie Canyon Rd, 31.52685642 -109.54466712, 4389ft., [APH_0719, 17/8/2009, 1♂, Alice Abela, AUMNH]; 0.83 miles SW Price Canyon Rd on Hwy-80, 31.62561536 -109.20644592, 4656ft., [APH_0724, 18/8/2009, 1♂, Alice Abela, AUMNH]; 19.8 miles NE of Douglas along Highway 80, 31.53231 -109.2941925, 4562ft., [AUMS_2246, 23/7/1989, 1♂, F.C. Baptista, AUMNH]; along Hwy-80, 31.46999359 -109.35916022, 4346ft., [APH_0723, 18/8/2009, 1♂, Alice Abela, AUMNH]; New Mexico: Hidalgo: along CR C001, 31.608773 -108.866361, 4966ft., [APH_1227, 7/8/2010, 1♂, Brent E. Hendrixson, Ashley Bailey, Andrea Reed, AUMNH]; along CR C004, 31.527331 -108.9057311, 5262ft., [APH_1226, 7/8/2010, 1♂, Brent E. Hendrixson, Ashley Bailey, Andrea Reed, AUMNH]; along Geronimo Trail, 31.528707 -108.9003311, 5032ft., [APH_1516, 8/9/2012, 1♀, Brent E. Hendrixson, AUMNH]; along Hwy-338, 31.607079 -108.8670811, 4934ft., [APH_0671-0673, 14/7/2009, 2♂, 1 juv, Brent E. Hendrixson, Nate Davis, AUMNH]; [APH_1188-1191, 26/7/2010, 4♂, Brent E. Hendrixson, Brendon Barnes, Nate Davis, AUMNH &
Distribution and natural history
Aphonopelma peloncillo can be found inhabiting the Madrean Archipelago Level III Ecoregion where it resides in lower to mid elevation habitats in and around the Peloncillo Mountains in southeastern Arizona and southwestern New Mexico (Fig.

Conservation status
Aphonopelma peloncillo has a somewhat narrow distribution in the United States that is restricted to middle and lower elevations in and around the Peloncillo Mountains of Arizona and New Mexico. These tarantulas are fairly common but this region is subject to grazing, illegal immigration, and other modes of habitat degradation. Future climate change throughout the sky island region is also a concern (
Remarks
This species was originally recognized by
Other important ratios that distinguish males: A. peloncillo possess a smaller PTl/M3 (≤0.82; 0.71–0.82) than A. catalina (≥0.86; 0.86–0.98), A. chiricahua (≥0.94; 0.94–1.10) and A. parvum (≥0.81; 0.81–0.88, with slight overlap), but larger than A. gabeli (≤0.63; 0.57–0.63); by possessing a larger L3 scopulation extent (52%-68%) than A. catalina (48%–53%, with slight overlap); by possessing a smaller F1/M4 (≤1.11; 0.99–1.11) than A. catalina (≥1.15; 1.15–1.22) and A. madera (≥1.12; 1.12–1.22), but larger than A. chalcodes (≤0.99; 0.92–0.99) and A. gabeli (≤0.97; 0.92–0.97); by possessing a larger T1/P4 (≥2.47; 2.47–2.94) than A. hentzi (≤2.46; 1.96–2.46). Other important ratios that distinguish females: A. peloncillo possess a larger M3/A4 (≥1.11; 1.11–1.23) than A. catalina (≤1.10; 1.07–1.10) and A. parvum (≤0.92; 0.86–0.92); by possessing a smaller L3 scopulation extent (58%–68%) than A. chalcodes (78%-93%), A. hentzi (67%–95%, with slight overlap), and A. gabeli (72%–80%). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma phasmus
Aphonopelma
(Delopelma)
phasmus Chamberlin, 1940: 28; male holotype from Phantom Ranch, Grand Canyon, Coconino Co., Arizona, 36.105315 -112.095201, elev. 2536ft., 26.vii.1934, coll. Dr. Lutz; deposited in
Rhechostica phasmus Raven, 1985: 149.
Aphonopelma phasmus Smith, 1995: 129.
Diagnosis
Based on carapace length (a proxy for body size) and evaluation of morphospace, Aphonopelma phasmus probably belongs to the Paloma species group but molecular data is needed to confirm its phylogenetic placement. Although the tip of the embolus is broken, the palpal bulb is unique (Fig.

Aphonopelma phasmus Chamberlin, 1940. A–I male holotype A dorsal view of carapace, scale bar = 1mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2.5mm E ventral view of metatarsus IV F prolateral view of L pedipalp and palpal tibia, scale bar = 3.5mm G dorsal view of palpal bulb (damaged) H retrolateral view of palpal bulb (damaged) I prolateral view of tibia I (mating clasper), scale bar = 2mm.
Description
Male originally described by
Redescription of male holotype
(Fig.
Distribution and natural history
Aphonopelma phasmus is known from a single adult male collected at Phantom Ranch (770 meters) near the Colorado River in Grand Canyon National Park in Coconino County, Arizona (Fig.

Conservation status
Aphonopelma phasmus probably has a very restricted distribution along the Colorado River in Grand Canyon National Park. Although the species is protected by the boundaries of the park, the Phantom Ranch area is prone to habitat degradation due to foot traffic and other recreational activities. New collecting efforts to find this species need to be taken before its conservation status can be assessed.
Species concept applied
Morphological Species Concept.
Remarks
Very little is known about this enigmatic species. Despite our extensive fieldwork throughout Arizona and surrounding areas, we have not encountered any other specimens that appear to be conspecific with A. phasmus. Focused collection efforts at Phantom Ranch and other locations along the Colorado River are necessary to fully grasp the validity of this species and to determine its phylogenetic placement. A handful of measurements demonstrate that A. phasmus likely is not conspecific with other species in the surrounding area (i.e., A. iodius, A. marxi, and A. prenticei) but new material is needed to confirm this hypothesis. Unfortunately, the holotype is badly damaged, fragmented, and missing multiple pieces, and because we lack more specimens, we are not able to understand the possible morphological variation that exists within this species. During evaluation of traditional PCA morphospace and three-dimensional PCA morphospace (PC1~PC2~PC3), A. phasmus groups with the other species in the Paloma species group (see Suppl. material
Aphonopelma prenticei , sp. n.
Aphonopelma mojave Prentice, 1997 (in part): 161.
Types
Male holotype (AUMS_2557) collected Alamo Rd., off I-40 and Yucca, 0.4 miles SE of campsite, Mohave Co., Arizona, 34.604435 -113.7512334, elev. 3300ft., 13.x.1993, coll. T.R. Prentice; deposited in AUMNH. Paratype female (APH_0319) from 0.4 miles S of Kelso Dunes Rd on Kelbaker Rd, San Bernardino Co., California, 34.89574 -115.650161, elev. 2825ft., 6.v.2008, coll. Brent E. Hendrixson and Zach Valois; deposited in AUMNH. Paratype male (APH_0415) from Mojave National Preserve, San Bernardino Co., California, 35.173875 -115.6095531, elev. 4130ft., 25.x.2008, coll. Anette Pillau; deposited in
Etymology
The specific epithet is a patronym in recognition of arachnologist Thomas R. Prentice for his work on the genus Aphonopelma.
Diagnosis
Aphonopelma prenticei (Fig.

Description of male holotype
(AUMS_2557; Fig.

Aphonopelma prenticei sp. n. A–I male holotype, AUMS_2557 A dorsal view of carapace, scale bar = 3mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2mm E ventral view of metatarsus IV, scale bar = 2mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2.5mm.
Variation (5).Cl 6.478–7.787 (7.244±0.24), Cw 5.42–6.948 (6.448±0.27), LBl 0.848–1.106 (0.992±0.05), LBw 0.933–1.204 (1.058±0.05), F1 6.693–8.289 (7.534±0.27), F1w 1.547–1.883 (1.744±0.06), P1 2.612–3.597 (3.022±0.17), T1 6.131–6.931 (6.536±0.17), M1 5.269–6.296 (5.817±0.18), A1 3.654–4.532 (4.094±0.14), L1 length 24.359–29.645 (27.003±0.87), F3 5.915–7.23 (6.599±0.22), F3w 1.806–2.612 (2.24±0.13), P3 2.104–3.078 (2.612±0.19), T3 4.631–5.693 (5.237±0.17), M3 5.727–6.952 (6.322±0.2), A3 3.64–4.396 (4.125±0.13), L3 length 22.017–27.275 (24.896±0.87), F4 6.814–8.616 (7.66±0.31), F4w 1.367–2.119 (1.778±0.12), P4 2.314–3.262 (2.688±0.17), T4 6.244–7.306 (6.722±0.2), M4 7.223–8.964 (8.075±0.28), A4 4.037–4.64 (4.326±0.11), L4 length 26.632–32.291 (29.471±0.93), PTl 4.305–4.746 (4.485±0.08), PTw 1.367–1.754 (1.53±0.07), SC3 ratio 0.604–0.728 (0.647±0.02), SC4 ratio 0.308–0.416 (0.355±0.02), Coxa I setae = thin/very thin tapered, F3 condition = swollen.
Description of female paratype
(APH_0319; Figs


Variation (6).Cl 7.186–8.426 (7.795±0.18), Cw 6.143–6.991 (6.582±0.14), LBl 1.115–1.415 (1.225±0.04), LBw 1.216–1.564 (1.308±0.05), F1 5.971–6.94 (6.296±0.15), F1w 1.803–2.093 (1.955±0.04), P1 2.51–3.35 (2.884±0.13), T1 4.764–5.732 (5.26±0.16), M1 3.692–4.337 (3.936±0.11), A1 3.018–3.856 (3.353±0.12), L1 length 20.434–23.665 (21.728±0.57), F3 4.845–5.662 (5.262±0.16), F3w 1.706–2.31 (1.868±0.09), P3 2.211–2.595 (2.458±0.05), T3 3.47–4.381 (3.878±0.13), M3 3.385–4.414 (4.008±0.17), A3 3.246–3.751 (3.44±0.07), L3 length 17.427–20.562 (19.047±0.52), F4 6.25–7.168 (6.744±0.17), F4w 1.736–1.993 (1.878±0.04), P4 2.474–3.124 (2.852±0.1), T4 5.215–6.022 (5.586±0.15), M4 4.96–5.997 (5.531±0.16), A4 3.519–4.473 (3.874±0.14), L4 length 22.644–26.604 (24.587±0.58), SC3 ratio 0.641–0.711 (0.671±0.01), SC4 ratio 0.273–0.384 (0.332±0.02), Coxa I setae = thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: La Paz: 0.4 miles NNW Alamo Rd on Low Mtn Rd, 33.92798 -113.545732, 2321ft., [APH_1113, 2/11/2009, 1♂, June Olberding, AUMNH]; 0.6 miles NE Alamo Rd on Low Mountain Rd, 33.91962 -113.539572, 2262ft., [APH_1112, 3/11/2009, 1♂, June Olberding, AUMNH]; 0.7 miles E Alamo Rd on 77th St, 7 miles N Wenden, 33.928133 -113.5309332, 2319ft., [APH_1586, 10/4/2011, 1♂, June Olberding, AUMNH]; 16 miles N US-60 on Alamo Rd (Mile Marker #16), 34.04226 -113.5665271, 1948ft., [APH_0495, 17/5/2009, 1 juv, Brent E. Hendrixson, Bernadette DeRussy, Sloan Click, AUMNH]; 5.5 miles S La Posa Rd on Plomosa Rd (S of Bouse), 33.85678 -114.028332, 1160ft., [APH_0798-0799, 2/11/2009, 2♀, June Olberding, AUMNH]; [APH_1100, 17/10/2009, 1♀, June Olberding, AUMNH]; 6.9 miles N US-60 on Alamo Rd, 33.9214 -113.54162, 2280ft., [APH_0797, 1/11/2009, 1♂, June Olberding, AUMNH]; 6.9 miles N US-60 on Alamo Rd (near Base of Harcuvar Mtns N of Wenden), 33.92479 -113.544191, 2285ft., [APH_0338-0339, 9/5/2008, 2 juv, Brent E. Hendrixson, Zach Valois, AUMNH]; Harcuvar Mtns, 1.6 miles W Low Mtn Rd along powerline road, 33.938266 -113.5301661, 2462ft., [APH_1563, 26/10/2012, 1♀, Brent E. Hendrixson, June Olberding, Doug Olberding, AUMNH]; [APH_1565-1567, 23/10/2012, 3♀, June Olberding, AUMNH]; Harcuvar Mtns, Low Mountain Rd, 33.91962 -113.539572, 2261ft., [APH_1252, 18/11/2010, 1♂, June Olberding, AUMNH]; Kofa National Wildlife Refuge, 33.4672 -113.805132, 1686ft., [APH_1659-1661, 23/11/2012, 1♀, 2 juv, June Olberding, AUMNH]; Maricopa: 3.7 miles S US-60 (Aguila) on Eagle Eye Rd, 33.891289 -113.1847131, 2323ft., [APH_0496-0497, 17/5/2009, 2 juv, Brent E. Hendrixson, Bernadette DeRussy, Sloan Click, AUMNH]; Mohave: 12 miles NW Chicken Springs Rd, 34.593079 -113.8134865, 3020ft., [AUMS_2552, 18/10/1992, 1♀, T.R. Prentice, AUMNH]; 21 miles NW Chicken Springs Rd on Alamo Rd, 34.80901 -114.049031, 2350ft., [APH_0350, 11/5/2008, 1 juv, Brent E. Hendrixson, Zach Valois, AUMNH]; 4.7 miles SE US-93/AZ-66 on Hualapai Mountain Rd, 35.162771 -113.9617041, 4285ft., [APH_0521, 24/5/2009, 1 juv, Brent E. Hendrixson, Bernadette DeRussy, AUMNH]; 5.6 miles NE Alamo Rd on Chicken Springs Rd, 34.66005 -113.746671, 3860ft., [APH_0348-0349, 11/5/2008, 2 juv, Brent E. Hendrixson, Zach Valois, AUMNH]; 7.4 miles NE Hwy-93 on CR-25 in Dolan Springs, 35.60418 -114.2535631, 3638ft., [APH_1310-1312, 28/7/2011, 3♀, Brent E. Hendrixson, Brendon Barnes, Nate Davis, Jake Storms, AUMNH]; Alamo Crossing Rd. 12 miles north of Chicken Springs Rd. jct, 34.716434 -113.9414245, 3001ft., [AUMS_2549, 24/10/1992, 1♀, T.R. Prentice, AUMNH]; Alamo Rd., 34.547893 -113.7544416, 2852ft., [APH_3115, unknown, 1♀, T.R. Prentice, AUMNH]; [AUMS_2544, 25/10/1994, 1♀, T.R. Prentice, AUMNH]; [AUMS_2545, 27/10/1994, 1♀, T.R. Prentice, AUMNH]; [AUMS_2548, 29/9/1994, 1♀, T.R. Prentice, AUMNH]; [AUMS_2561, 25/10/1994, 1♂, T.R. Prentice, AUMNH]; [AUMS_2562, 29/9/1994, 1♂, T.R. Prentice, AUMNH]; [AUMS_2568, unknown, 1♀, T.R. Prentice, AUMNH]; [AUMS_2569, 25/10/1994, 1♂, T.R. Prentice, AUMNH]; Alamo Rd., 0.5 miles NW of Alamo Rd., 34.614423 -113.7947584, 3220ft., [AUMS_2555, 13/10/1993, 1♂, T.R. Prentice, AUMNH]; Alamo Rd., 0.5 miles S of campsite, 34.580474 -113.7857336, 3083ft., [AUMS_2559, 29/9/1994, 1♂, T.R. Prentice, AUMNH]; Alamo Rd., off I-40 and Yucca, 0.4 miles SE of campsite, 34.604435 -113.7512334, 3300ft., [AUMS_2557, 13/10/1993, 1♂, T.R. Prentice, AUMNH]; Alamo Road, 12 miles W of Chicken Springs jct, 34.593874 -113.7872425, 3120ft., [AUMS_2563, 30/3/1993, 1♂, T.R. Prentice, AUMNH]; along Hualapai Mountain Rd, 35.162658 -113.9614191, 4315ft., [APH_0775-0778, 7/10/2009, 4 juv, Brent E. Hendrixson, Thomas Martin, AUMNH]; campsite, Alamo rd. crossing, 34.597711 -113.7969625, 3100ft., [AUMS_2564, 29/3/1993, 1♂, T.R. Prentice, AUMNH]; Chicken Springs Rd., 34.607061 -113.7676136, 3250ft., [AUMS_2566, 23/6/1992, 1♂, T.R. Prentice, AUMNH]; Chicken Springs Rd., 1.4 miles W of Alamo rd. crossing, 34.582903 -113.8139534, 3060ft., [AUMS_2558, 20/9/1992, 1♂, T.R. Prentice, AUMNH]; Chicken Springs Rd., 1.5 miles E of jct, 34.6148 -113.7696045, 3300ft., [AUMS_2560, 23/6/1992, 1♂, T.R. Prentice, AUMNH]; Chicken Springs Rd., 1.7 miles N of Alamo Rd. jct., 34.607695 -113.7718494, 3250ft., [AUMS_2556, 23/6/1992, 1♂, T.R. Prentice, AUMNH]; Chicken Springs Rd., 2 miles N of Alamo rd. crossing, 34.609856 -113.774015, 3250ft., [AUMS_2553-2554, 20/9/1992, 2♀, T.R. Prentice, AUMNH &
Distribution and natural history
Aphonopelma prenticei has a large range throughout the eastern Mojave (southeastern California, southern Nevada, southwestern Utah, northwestern Arizona) and northwestern Sonoran (western Arizona) deserts at elevations between 350 and 1525 meters. Much of the distribution is east of the Death Valley drainage system with the exception of a few populations along the Panamint Range (Fig.

Conservation status
Aphonopelma prenticei is the most abundant and widespread turret-building tarantula species in the United States and has recently expanded its range into the more northern portions of its distribution (
Remarks
Other important ratios that distinguish females: A. prenticei possess a larger Cl/M4 (≥1.31; 1.31–1.52) than A. mojave (≤1.31; 1.20–1.31) and A. joshua (≤1.25; 1.20–1.25). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma saguaro , sp. n.
Types
Male holotype (APH_3220) collected from Sabino Canyon, Tucson, Pima Co., Arizona, 32.322533 -110.8095612, elev. 2774ft., 23.xi.2014, coll. Philip MacDuff; deposited in AUMNH. Paratype female (APH_3176) from Saguaro National Park - Saguaro East (Rincon Mountain District), off Cactus Forest Loop Dr., before the Mica View picnic area, Pima Co., Arizona, 32.198568 -110.7356141, elev. 2944ft., 12.xi.2013, coll. Brent E. Hendrixson and Chris A. Hamilton; deposited in AUMNH. Paratype male (APH_2540) from Sabino Canyon, Tucson, Pima Co., Arizona, 32.316614 -110.817455, elev. 2818ft., date unknown, coll. O. Bryant; deposited in
Etymology
The specific epithet is a noun in apposition taken from type locality, Saguaro National Park, where this new species can be found in and around the foothills of the Santa Catalina and Rincon Mountains.
Diagnosis
Aphonopelma saguaro (Fig.

Description of male holotype
(APH_3220; Fig.

Aphonopelma saguaro sp. n. A–I male holotype, APH_3220 A dorsal view of carapace, scale bar = 2.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2mm E ventral view of metatarsus IV, scale bar = 2mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2mm.
Variation (6).Cl 4.554–6.165 (5.362±0.25), Cw 4.074–5.719 (4.998±0.23), LBl 0.576–0.961 (0.745±0.06), LBw 0.816–1.148 (0.997±0.05), F1 4.764–6.232 (5.717±0.21), F1w 1.121–1.593 (1.387±0.07), P1 1.732–2.444 (2.106±0.1), T1 4.33–5.674 (5.158±0.2), M1 3.394–4.346 (3.964±0.14), A1 2.542–3.241 (2.949±0.1), L1 length 16.76–21.695 (19.894±0.74), F3 3.941–5.31 (4.772±0.21), F3w 1.268–1.985 (1.666±0.11), P3 1.481–2.071 (1.777±0.09), T3 3.289–4.241 (3.892±0.15), M3 3.643–4.952 (4.449±0.21), A3 2.633–3.581 (3.156±0.15), L3 length 14.985–20.023 (18.046±0.79), F4 4.683–6.314 (5.591±0.23), F4w 1.037–1.736 (1.306±0.09), P4 1.541–2.216 (1.917±0.09), T4 4.435–5.664 (5.152±0.19), M4 4.675–6.393 (5.661±0.25), A4 3.212–3.993 (3.497±0.12), L4 length 18.545–23.99 (21.817±0.82), PTl 3.036–3.953 (3.612±0.14), PTw 1.238–1.644 (1.449±0.06), SC3 ratio 0.306–0.521 (0.402±0.03), SC4 ratio 0.146–0.263 (0.202±0.02), Coxa I setae = fine & thin hair-like, F3 condition = slightly swollen/swollen.
Description of female paratype
(APH_3176; Fig.

Aphonopelma saguaro sp. n. A–F female paratype, APH_3176 A dorsal view of carapace, scale bar = 3mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 1.5mm D ventral view of metatarsus IV, scale bar = 1.5mm E prolateral view of L pedipalp and palpal tibia F cleared spermathecae.
Material examined
United States: Arizona: Pima: Sabino Canyon, Tucson, 32.316614 -110.817455, 2818ft., [APH_2540, date unknown, 1♂, O. Bryant,
Distribution and natural history
Aphonopelma saguaro can be found inhabiting the Madrean Archipelago and Sonoran Basin and Range Level III Ecoregions in southeastern Arizona where their distribution occurs in the foothills and lower canyons of the Santa Catalina and Rincon Mountains (Fig.

Conservation status
Aphonopelma saguaro appears to have a limited distribution restricted to the foothills and lower canyons in and around the Santa Catalina and Rincon Mountains. As evidenced by only one specimen previously collected and deposited in a museum collection prior to this study, A. saguaro can be difficult to find due to the cryptic nature of their burrows and narrow window of activity during the year. The Tucson Metropolitan area is experiencing rapid growth and recreational activities in the Santa Catalina and Rincon Mountains threaten suitable habitat for this species. The status of Aphonopelma saguaro seems secure (e.g., protection in Saguaro National Park) but should be monitored.
Remarks
Other important ratios that distinguish males: A. saguaro possess a larger T3/F4 (≥0.67; 0.67–0.71) than A. paloma (≤0.64; 0.57–0.64) and A. parvum (≤0.66; 0.61–0.66); a larger PTl/F4 (≥0.62; 0.62–0.66) than A. chalcodes (≤0.62; 0.52–0.62), A. superstitionense (≤0.59; 0.55–0.59), and A. vorhiesi (≤0.62; 0.55–0.62); a smaller Cl/M3 (≤1.25; 1.17–1.25) than A. catalina (≥1.26; 1.26–1.42); a smaller F1/T3 (≤1.51; 1.42–1.51) than A. parvum (≥1.56; 1.56–1.70); a smaller Cl/PTw (≤4.27; 3.13–4.27) than A. chalcodes (≥5.06; 5.06–6.05) and A. superstitionense (≥4.40; 4.40–4.72); a smaller L3 scopulation extent (30%–52%) than A. chalcodes (65%–86%), A. parvum (60%–65%), and A. vorhiesi (53%–62%). Other important ratios that distinguish females: A. saguaro possess a smaller F3L/W (2.41 ± (only 1 specimen)) than A. catalina (≥2.93; 2.93–2.99), A. chalcodes (≥2.84; 2.84–3.15), A. mareki (≥2.66; 2.66–3.00), A. parvum (≥2.85; 2.85–3.05), A. superstitionense (2.75 ± (only 1 specimen)), and A. vorhiesi (≥2.77; 2.77–3.11). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma steindachneri
Eurypelma steindachneri Ausserer, 1875: 199; male holotype from Wruck Canyon, SE San Ysidro, San Diego Co., California, 32.548946 -117.0061354, elev. 237ft., unknown collection date, coll. Dr. Steindachner; deposited in
Delopelma steindachneri Petrunkevitch, 1939: 253.
Rhechostica steindachneri Raven, 1985: 160.
Aphonopelma steindachneri Smith, 1995: 147.
Aphonopelma steindachneri - male neotype designated (APH_1023) from Wruck Canyon, San Ysidro, off Cactus Rd., San Diego Co., California, 32.55002 -116.991921, elev. 398ft., 16.v.2010, coll. Chris A. Hamilton, Xavier Atkinson, Jordan Satler; deposited in AUMNH.
Aphonopelma phanus Chamberlin, 1940: 24; male holotype from Laguna Beach, Orange Co., California, 33.542248 -117.7831105; elev. 18ft., vii.1931, coll. unknown; deposited in
Rhechostica phanus Raven, 1985: 149.
Aphonopelma phanus Smith, 1995: 128.
Aphonopelma phanum. syn. n.
Aphonopelma reversum Chamberlin, 1940: 8; male holotype, male paratype, three female paratypes from San Diego, San Diego Co., California, 32.715738 -117.1610856, elev. 54ft., 1935, coll. unknown; deposited in
Rhechostica reversum Raven, 1985: 149.
Aphonopelma reversum Smith, 1995: 133. syn. n.
Diagnosis
Aphonopelma steindachneri (Fig.

Description
Originally described by
Description of male neotype
(APH_1023; Fig.

Aphonopelma steindachneri (Ausserer, 1875). A–I male neotype, APH_1023 A dorsal view of carapace, scale bar = 7mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 5mm E ventral view of metatarsus IV, scale bar = 3.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 5.5mm.
Variation (7).Cl 12.74–19.08 (15.197±0.84), Cw 12.19–17.33 (14.029±0.75), LBl 1.53–2.20 (1.77±0.09), LBw 1.77–2.91 (2.123±0.15), F1 13.84–17.76 (15.367±0.59), F1w 3.1–4.31 (3.621±0.17), P1 5.69–7.38 (6.231±0.25), T1 12.58–15.12 (13.684±0.4), M1 10.6–14.12 (11.837±0.5), A1 6.64–8.71 (7.49±0.26), L1 length 49.48–63.09 (54.61±1.96), F3 11.58–14.89 (12.794±0.47), F3w 3.43–4.89 (4.053±0.2), P3 4.66–6.22 (5.437±0.23), T3 9.39–11.77 (10.309±0.39), M3 11.31–14.94 (12.511±0.52), A3 6.58–8.72 (7.356±0.27), L3 length 43.72–56.54 (48.407±1.83), F4 13.38–17.36 (14.827±0.58), F4w 3.06–4.52 (3.624±0.2), P4 4.96–6.38 (5.604±0.21), T4 11.63–15.45 (12.823±0.54), M4 15.04–20.16 (16.714±0.73), A4 7.31–9.01 (7.997±0.23), L4 length 52.75–68.36 (57.966±2.26), PTl 8.359–10.514 (9.091±0.31), PTw 2.45–3.05 (2.708±0.08), SC3 ratio 0.401–0.542 (0.479±0.02), SC4 ratio 0.22–0.314 (0.282±0.01), Coxa I setae = tapered/thin tapered, F3 condition = slightly swollen.
Description of female exemplar
(APH_1022; Figs

Aphonopelma steindachneri (Ausserer, 1875). A–E female specimen, APH_1022 A dorsal view of carapace, scale bar = 7.5mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 3mm D ventral view of metatarsus IV, scale bar = 4.5mm E prolateral view of L pedipalp and palpal tibia. Notice the eye deformity, a feature seen in the original species description.

Variation (6).Cl 16.59–20.22 (18.295±0.53), Cw 14.55–17.96 (16.215±0.52), LBl 1.97–2.43 (2.178±0.09), LBw 2.27–2.77 (2.458±0.09), F1 11.91–14.61 (13.75±0.39), F1w 3.98–4.73 (4.41±0.12), P1 5.57–7.18 (6.538±0.24), T1 8.95–12.03 (11.19±0.47), M1 7.46–8.74 (8.207±0.23), A1 6.24–7.33 (6.608±0.16), L1 length 40.13–48.99 (46.293±1.35), F3 10.46–12.21 (11.417±0.29), F3w 3.71–4.35 (3.967±0.09), P3 4.79–6.50 (5.662±0.23), T3 7.62–8.91 (8.312±0.17), M3 7.87–9.44 (8.897±0.24), A3 6.33–6.93 (6.595±0.1), L3 length 37.25–43.66 (40.882±0.89), F4 12.85–14.79 (13.858±0.29), F4w 3.94–4.49 (4.157±0.08), P4 5.08–6.89 (5.93±0.25), T4 10.37–11.44 (11.083±0.16), M4 11.71–13.73 (12.688±0.32), A4 6.86–7.54 (7.175±0.1), L4 length 47.11–52.78 (50.735±0.89), SC3 ratio 0.513–0.608 (0.574±0.01), SC4 ratio 0.245–0.344 (0.3±0.01), C1 setae = tapered/thin tapered. Spermathecae variation can be seen in Figure
Material examined
United States: California: Kern: above Frazier Park Ranger Station, 34.800403 -118.9965825, 5700ft., [AUMS_3298, 18/7/1987, 1♀, T.R. Prentice, AUMNH]; Caliente Creek, 20 miles N Hwy 58 + 2.5 mils N of Twin Oaks country store, Piute Mtns, 35.345195 -118.3863525, 3348ft., [AUMS_2603, 11/6/1989, 1♀, T.R. Prentice, AUMNH]; Erskine Creek (Canyon), Erskine Creek road, 35.593637 -118.4459715, 2933ft., [AUMS_3322, 10/1970, 1♀, J. Anderson, AUMNH]; N of Tehachapi, off Tehachapi Loop (Woodford-Tehachapi Rd), 35.20873 -118.550611, 2821ft., [APH_3100, 17/7/2012, 1♀, Chris A. Hamilton, Amy Skibiel, AUMNH]; near Lake Isabella, off Kern River Canyon Rd, near Bodfish and Lake Isabella Blvd intersection, 35.5912 -118.515631, 2662ft., [APH_3096-3098, 14/7/2012, 3♀, Chris A. Hamilton, Jim Starrett, Amy Skibiel AUMNH]; S of Tehachapi, off Sand Canyon Rd - off Hwy 58, 35.16025 -118.325071, 4144ft., [APH_3099, 17/7/2012, 1♂, Chris A. Hamilton, Amy Skibiel, AUMNH]; Los Angeles: 27832 Stonehill Way, Canyon Country, 34.42464 -118.452572, 1525ft., [APH_0362, 6/7/2008, 1♂, Anette Pillau, AUMNH]; [APH_0413, 23/8/2008, 1♀, Anette Pillau, AUMNH]; Azusa, 3 miles west in Fish Canyon, 34.135573 -117.9409725, 581ft., [APH_2694, 23/9/1956, 1♂, V. Roth,
Distribution and natural history
Aphonopelma steindachneri is distributed from northern Baja California, north into California along the Southern Californian Coast Ranges, bounded by the Mojave Desert, across the Tehachapi Mountains, and into the southern portions of the Sierra Nevada Mountains (Fig.

Conservation status
Aphonopelma steindachneri is widely distributed across Southern California and is very common. The species is likely secure although some localized populations in urbanized areas (e.g., Los Angeles and San Diego) are likely threatened by human encroachment and development.
Remarks
Other important ratios that distinguish males: A. steindachneri possess a smaller L3 scopulation extent (40%–54%) than A. eutylenum (72%–93%), A. iodius (78%–96%) and A. johnnycashi (71%–82%); by possessing a larger T1/F3 (≥1.01; 1.01–1.11) than A. eutylenum (≤1.00; 0.93–1.00) and A. johnnycashi (≤1.02; 0.95–1.02, with slight overlap). Other important ratios that distinguish females: A. steindachneri possess a smaller L3 scopulation extent (51%–61%) than A. eutylenum (77%–91%), A. iodius (72%–95%), and A. johnnycashi (69%–92%); by possessing a smaller M1/M4 (≤0.67; 0.62–0.67) than A. eutylenum (≥0.67; 0.67–0.78) and A. johnnycashi (0.70–0.74). For both males and females, certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no others are claimed to be significant at this time (see Suppl. material
Aphonopelma superstitionense , sp. n.
Types
Male holotype (APH_1443) collected from the Superstition Mtns, Flat Iron, just above Flat Bowl Siphon Draw, 33.440386 -111.4576021, elev. 3450ft., 1.xii.2011, coll. Tim Cota; deposited in AUMNH. Paratype female (APH_0504) from 0.25 miles S Crimson Rd on Usery Pass Rd., 33.479188 -111.6256081, elev. 2009ft., 19.v.2009, coll. Brent E. Hendrixson, Bernadette DeRussy, Sloan Click; deposited in AUMNH. Paratype male (APH_1607) from the Superstition Mtns, Peralta Canyon Trail, 33.408937 -111.3540861, elev. 2982ft., 11.xi.2012, coll. Brent E. Hendrixson, Tim Cota, Molly Taylor; deposited in
Etymology
The specific epithet is a neuter adjective taken from type locality, the Superstition Mountains, where this species was first discovered.
Diagnosis
Aphonopelma superstitionense (Fig.

Description of male holotype
(APH_1443; Fig.

Aphonopelma superstitionense sp. n. A–I male holotype, APH_1443 A dorsal view of carapace, scale bar = 2.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 2mm E ventral view of metatarsus IV, scale bar = 2mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 0.5mm I prolateral view of tibia I (mating clasper), scale bar = 2.5mm.
Variation (3).Cl 5.594–6.586 (5.961±0.31), Cw 5.171–6.302 (5.592±0.36), LBl 0.824–0.93 (0.869±0.03), LBw 1.006–1.044 (1.026±0.01), F1 6.624–7.28 (6.903±0.2), F1w 1.308–1.469 (1.37±0.05), P1 2.234–2.382 (2.32±0.04), T1 5.731–6.501 (6.078±0.23), M1 4.719–4.839 (4.771±0.04), A1 3.387–3.764 (3.535±0.12), L1 length 22.843–24.729 (23.607±0.57), F3 5.475–6.065 (5.708±0.18), F3w 1.592–1.923 (1.738±0.1), P3 1.878–2.021 (1.957±0.04), T3 4.545–4.916 (4.676±0.12), M3 5.127–5.883 (5.473±0.22), A3 3.398–3.622 (3.53±0.07), L3 length 20.618–22.234 (21.345±0.47), F4 6.418–6.966 (6.753±0.17), F4w 1.294–1.538 (1.389±0.08), P4 2.061–2.265 (2.149±0.06), T4 5.902–6.666 (6.193±0.24), M4 6.571–7.264 (6.983±0.21), A4 3.871–3.929 (3.902±0.02), L4 length 24.823–27.09 (25.98±0.65), PTl 3.786–3.951 (3.841±0.05), PTw 1.236–1.394 (1.308±0.05), SC3 ratio 0.404–0.477 (0.429±0.02), SC4 ratio 0.147–0.204 (0.185±0.02), Coxa I setae = very thin tapered, F3 condition = slightly swollen/swollen.
Description of female paratype
(APH_0504; Fig.

Aphonopelma superstitionense sp. n. A–F female paratype, APH_0504 A dorsal view of carapace, scale bar = 3mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 1.5mm D ventral view of metatarsus IV, scale bar = 1.5mm E prolateral view of L pedipalp and palpal tibia F cleared spermathecae.
Material examined
United States: Arizona: Maricopa: 0.25 miles S Crimson Rd on Usery Pass Rd, 33.479188 -111.6256081, 2009ft., [APH_0504, 19/5/2009, 1♀, Brent E. Hendrixson, Bernadette DeRussy, Sloan Click, AUMNH]; Apache Trail, 33.540608 -111.3454272, 2651ft., [APH_1643, 29/11/2012, 1♂, Tim Cota, AUMNH]; Pinal: Superstition Mtns, Flat Iron, just above Flat Bowl Siphon Draw, 33.440386 -111.4576021, 3450ft., [APH_1443, 1/12/2011, 1♂, Tim Cota, AUMNH]; Superstition Mtns, Peralta Canyon Trail, 33.408937 -111.3540861, 2982ft., [APH_1607, 11/11/2012, 1♂, Brent E. Hendrixson, Tim Cota, Molly Taylor,
Distribution and natural history
Aphonopelma superstitionense can be found inhabiting the Arizona/New Mexico Mountains and Sonoran Basin and Range Level III Ecoregions of central Arizona east of the Phoenix Metropolitan Area (Fig.

Conservation status
Aphonopelma superstitionense appears to have a limited distribution restricted to the foothills and area in or around the Superstition Mountains. These diminutive tarantulas are probably common but can be incredibly difficult to find due to the cryptic nature of their burrows and narrow window of activity during the year. The Phoenix Metropolitan area is experiencing rapid growth and recreational activities in the Superstition Mountains threaten suitable habitat for this species. The status of A. superstitionense seems secure (e.g., it is probably common in remote sections of the Superstition Wilderness Area) but should be monitored.
Remarks
Other important ratios that distinguish males: A. superstitionense possess a smaller PTl/M3 (≤0.74; 0.67–0.74) than A. mareki (≥0.76; 0.76–0.92), A. paloma (≥0.77; 0.77–0.85), A. parvum (≥0.81; 0.81–0.88), and A. saguaro (≥0.78; 0.78–0.84); a smaller L3 scopulation extent (40%–48%) than A. chalcodes (65%-86%), A. mareki (50%–56%), and A. parvum (60%–65). Other important ratios that distinguish females: A. superstitionense possess a smaller M3/M4 (0.62 ± (only 1 specimen)) than A. mareki (≥0.66; 0.66–0.77), A. parvum (≥0.67; 0.67–0.74), and A. saguaro (0.70 ± (only 1 specimen)); a smaller Cl/M4 (1.25 ± (only 1 specimen)) than A. paloma (≥1.44; 1.44–1.60); a larger L1/L3 (1.19 ± (only 1 specimen)) than A. parvum (≥1.09; 1.09–1.14); a larger F1/T1 (1.30 ± (only 1 specimen)) than A. saguaro (1.13 ± (only 1 specimen)). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Aphonopelma vorhiesi
Delopelma vorhiesi Chamberlin & Ivie, 1939: 7; male holotype and male paratype from Tucson, Pima Co., Arizona, 32.221743 -110.9264796, elev. 2473ft., no collecting date, coll. Prof. C.T. Vorhies; deposited in
Rhechostica vorhiesi Raven, 1985: 149.
Aphonopelma vorhiesi Smith, 1995: 155.
Aphonopelma jungi Smith, 1995: 116; male holotype from Portal Rd., Portal, Cochise Co., Arizona, 31.914142 -109.1416765, elev. 4761ft., viii.1992, coll. A. Smith and Michael Sullivan; deposited in BMNH. [examined] syn. n.
Aphonopelma punzoi Smith, 1995: 131; male holotype from Swift Trail (Jct), 5 miles S of Safford, Graham Co., Arizona, 32.729897 -109.7139215, elev. 3161ft., mid-August, coll. A. Smith and Michael Sullivan; deposited in BMNH. [examined] syn. n.
Diagnosis
Aphonopelma vorhiesi (Fig.

Description
Male originally described by
Redescription of male exemplar
(APH_0177; Fig.

Aphonopelma vorhiesi (Chamberlin & Ivie, 1939). A–I male specimen, APH_0177 A dorsal view of carapace, scale bar = 3.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 4mm F prolateral view of L pedipalp and palpal tibia, scale bar = 4mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 4mm.
Variation (11).Cl 10.018–13.98 (11.663±0.4), Cw 9.18–12.95 (10.722±0.4), LBl 1.11–1.658 (1.404±0.06), LBw 1.24–1.921 (1.62±0.07), F1 10.15–14.51 (11.867±0.42), F1w 2.51–3.62 (2.917±0.1), P1 4.1–5.88 (4.885±0.18), T1 8.69–12.39 (10.09±0.36), M1 6.877–10.20 (8.296±0.3), A1 4.74–6.70 (5.584±0.21), L1 length 35.23–49.06 (40.541±1.51), F3 8.37–11.57 (9.858±0.34), F3w 2.65–4.09 (3.18±0.14), P3 3.3–4.47 (3.794±0.12), T3 6.49–9.25 (7.619±0.28), M3 7.717–10.89 (8.927±0.34), A3 5.35–6.97 (6.075±0.17), L3 length 31.4–43.04 (36.647±1.26), F4 10.09–13.94 (11.696±0.41), F4w 2.32–3.64 (2.831±0.13), P4 3.46–4.56 (4.129±0.11), T4 8.996–11.74 (10.128±0.31), M4 10.33–14.66 (12.131±0.44), A4 5.43–7.83 (6.606±0.25), L4 length 38.39–52.73 (44.469±1.57), PTl 6.167–8.125 (6.983±0.18), PTw 1.919–2.611 (2.334±0.06), SC3 ratio 0.45–0.619 (0.554±0.01), SC4 ratio 0.201–0.358 (0.3±0.01), Coxa 1 setae = tapered/thin tapered, F3 condition = slightly swollen.
Description of female exemplar
(APH_1488; Figs

Aphonopelma vorhiesi (Chamberlin & Ivie, 1939). A–E female specimen, APH_1488 A dorsal view of carapace, scale bar = 5mm B prolateral view of coxa I C ventral view of metatarsus III, scale bar = 4mm D ventral view of metatarsus IV, scale bar = 3.5mm E prolateral view of L pedipalp and palpal tibia.

Variation (8).Cl 11.23–16.38 (13.636±0.64), Cw 9.77–14.63 (11.819±0.59), LBl 1.48–1.99 (1.778±0.08), LBw 1.77–2.42 (2.06±0.09), F1 8.86–12.61 (10.74±0.52), F1w 2.86–4.06 (3.401±0.16), P1 4.17–6.22 (5.001±0.24), T1 7.09–9.97 (8.447±0.38), M1 5.08–8.17 (6.301±0.38), A1 4.46–6.37 (5.274±0.23), L1 length 29.66–43.34 (35.763±1.72), F3 7.54–10.52 (8.761±0.42), F3w 2.45–3.79 (2.978±0.15), P3 3.26–4.93 (3.99±0.22), T3 4.87–7.70 (6.312±0.38), M3 5.24–8.13 (6.611±0.39), A3 4.65–6.72 (5.367±0.25), L3 length 25.56–37.60 (31.041±1.6), F4 8.93–12.91 (10.773±0.56), F4w 2.67–3.81 (3.076±0.14), P4 3.39–5.35 (4.438±0.22), T4 7.54–10.69 (8.885±0.4), M4 7.87–11.47 (9.504±0.46), A4 5.44–7.32 (6.122±0.25), L4 length 33.76–47.58 (39.721±1.85), SC3 ratio 0.495–0.69 (0.604±0.02), SC4 ratio 0.269–0.369 (0.332±0.01), Coxa I setae = tapered. Spermathecae variation can be seen in Figure
Material examined
United States: Arizona: Cochise: 0.45 miles NNE Cazador Trl on Hwy 80, 31.4724344 -109.45026412, 4554ft., [APH_0720-0721, 17/8/2009, 2♂, Alice Abela, AUMNH]; 1 mile west of Portal, 31.914744 -109.1625065, 5026ft., [APH_2130, 24/9/1963, 1♂, V. Roth,
Distribution and natural history
Aphonopelma vorhiesi is widely distributed across southeastern Arizona and southern New Mexico (Fig.

Conservation status
Aphonopelma vorhiesi is very common throughout its distribution in southeastern Arizona and southern New Mexico. The species is secure.
Remarks
Mitochondrial DNA identifies A. vorhiesi as a paraphyletic group with respect to A. chalcodes (Fig.
We examined the holotypes and freshly collected topotypic material of A. jungi and A. punzoi. Our morphological and molecular analyses fail to recognize these two species as separate, independently evolving lineages. As a consequence, we consider A. jungi and A. punzoi junior synonyms of A. vorhiesi.
Aphonopelma xwalxwal , sp. n.
Types
Male holotype (APH_3134) collected on Hwy S22 (Montezuma Valley Rd), 3.3 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, San Diego Co., California, 33.225633 -116.4146331, elev. 1710ft., 4.x.2013, coll. Wendell Icenogle; deposited in AUMNH. Paratype male (APH_3133) from Hwy S22 (Montezuma Valley Rd), 4 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, San Diego Co., California, 33.224618 -116.4242321, elev. 1935ft., 5.x.2013, coll. Wendell Icenogle; deposited in
Etymology
The specific epithet is a noun in apposition, referencing the word for “a type of small spider” in the language of the Cahuilla Native Americans, whose traditional territory includes the distribution of this species (from the Coachella Valley to Borrego Springs). Pronounced “hwalhwal”; xw - sounds like a light raspy sound when blowing out a candle; a - sounds like “a” in father; l - sounds like “l” in light.
Diagnosis
Aphonopelma xwalxwal (Fig.

Description of male holotype
(APH_3134; Fig.

Aphonopelma xwalxwal sp. n. A–I male holotype, APH_3134 A dorsal view of carapace, scale bar = 3.5mm B prolateral view of coxa I C dorsal view of femur III D ventral view of metatarsus III, scale bar = 4mm E ventral view of metatarsus IV, scale bar = 4.5mm F prolateral view of L pedipalp and palpal tibia, scale bar = 2.5mm G dorsal view of palpal bulb H retrolateral view of palpal bulb, scale bar = 1mm I prolateral view of tibia I (mating clasper), scale bar = 4mm.
Variation (5).Cl 8.389–10.491 (9.24±0.36), Cw 7.387–9.92 (8.664±0.45), LBl 1.142–1.454 (1.24±0.06), LBw 1.211–1.503 (1.361±0.06), F1 9.357–11.849 (10.398±0.43), F1w 2.233–2.672 (2.407±0.09), P1 3.656–4.797 (4.099±0.21), T1 8.907–11.723 (10.242±0.49), M1 7.681–10.68 (9.069±0.51), A1 4.786–6.363 (5.39±0.27), L1 length 34.436–45.412 (39.197±1.9), F3 7.71–10.492 (9.045±0.47), F3w 2.537–3.297 (2.854±0.13), P3 3.217–3.743 (3.398±0.1), T3 7.011–9.921 (8.28±0.49), M3 8.272–11.209 (9.72±0.5), A3 4.884–6.055 (5.419±0.22), L3 length 31.169–41.42 (35.861±1.74), F4 9.182–12.649 (10.644±0.59), F4w 2.179–2.81 (2.357±0.11), P4 3.471–4.192 (3.708±0.14), T4 9.03–12.006 (10.39±0.51), M4 11.074–14.367 (12.661±0.57), A4 5.047–6.798 (5.724±0.31), L4 length 38.048–50.012 (43.127±2.02), PTl 4.878–6.382 (5.53±0.25), PTw 1.309–1.744 (1.532±0.07), SC3 ratio 0.651–0.947 (0.773±0.07), SC4 ratio 0.348–0.485 (0.405±0.02), Coxa I setae = very thin tapered, F3 condition = swollen.
Material examined
United States: California: Riverside: Carrizo Creek, .25 miles S of Hwy 74 bridge, 4 miles S of Palm Desert or Hwy 111, 33.643993 -116.4003174, 2572ft., [AUMS_3319, 27/3/1972, 1♂, W. Icenogle, AUMNH]; Deep Canyon, 33.669056 -116.3676835, 699ft., [AUMS_2668, 23/10/1963, 1♂, E. Schlinger, AUMNH]; on Hwy 74, 2.5 miles S of Carrizo Creek bridge, 6.5 miles S of Palm Desert or Hwy 111, 33.627668 -116.403534, 2925ft., [AUMS_3318, 15/10/1976, 1♂, W. Icenogle, AUMNH]; P.L. Boyd Deep Canyon Reserve Center, 3.5 miles S of Palm Desert, 33.674866 -116.3712614, 639ft., [AUMS_2670, 13/10/1969, 1♂, Saul Frommer, AUMNH]; P.L. Boyd Deep Canyon Reserve, 0.4 miles N of reserve station, 33.653477 -116.374194, 891ft., [AUMS_2348, 22/9/1995, 1♂, T.R. Prentice, AUMNH]; P.L. Boyd Deep Canyon Reserve, 0.5 miles below reserve gate, 33.677408 -116.3701094, 613ft., [AUMS_2345, 27/9/1995, 1♂, T.R. Prentice, AUMNH]; P.L. Boyd Deep Canyon Reserve, 1.1 miles below reserve gate, 33.684368 -116.3648174, 558ft., [AUMS_2352, 21/9/1995, 1♂, T.R. Prentice, AUMNH]; P.L. Boyd Deep Canyon Reserve, at reserve gate - North, 33.670726 -116.3729494, 684ft., [AUMS_2344, 21/9/1995, 1♂, T.R. Prentice, AUMNH]; P.L. Boyd-Deep Canyon Reserve, 0.2 miles below reserve gate, 33.586027 -116.2764514, 613ft., [AUMS_3280, 27/9/1995, 1♂, T.R. Prentice, AUMNH]; San Diego: Hwy S22 (Montezuma Valley Rd), 1.6 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, 33.2409 -116.4032831, 1148ft., [APH_3132, 5/10/2013, 1♂, W. Icenogle, AUMNH]; Hwy S22 (Montezuma Valley Rd), 2.8 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, 33.231167 -116.4116061, 1474ft., [APH_3130, 4/10/2013, 1♂, W. Icenogle, AUMNH]; Hwy S22 (Montezuma Valley Rd), 3.2 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, 33.225583 -116.4130671, 1673ft., [APH_3131, 4/10/2013, 1♂, W. Icenogle, AUMNH]; Hwy S22 (Montezuma Valley Rd), 3.3 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, 33.225633 -116.4146331, 1710ft., [APH_3134, 4/10/2013, 1♂, W. Icenogle, AUMNH]; Hwy S22 (Montezuma Valley Rd), 4 miles S junction with Palm Canyon Dr. (Borrego Springs), Anza-Borrego State Park, 33.224618 -116.4242321, 1935ft., [APH_3133, 5/10/2013, 1♂, W. Icenogle,
Distribution and natural history
Aphonopelma xwalxwal is presently known from only two areas along the foothills and mountains west of Palm Springs and Borrego Springs (Fig.

Conservation status
The distribution of Aphonopelma xwalxwal appears to be restricted to the mountains and foothills on the eastern side of the Peninsular Ranges and San Jacinto Mountains. In historical collections, this species appears to have been collected somewhat infrequently. Based on our own fieldwork in search of females of this species, we found burrows to be incredibly difficult to find due to the steep, rocky nature of their habitat, and the harsh, dry environments that likely keeps them inactive for most of the year. The species is likely secure, but populations should be monitored due to the continued expansion of human activity and development in Southern California.
Remarks
Other important ratios that distinguish males: A. xwalxwal possess a larger T3/A3 (≥1.41; 1.41–1.64) than A. eutylenum (≤1.35; 1.30–1.35), A. icenoglei (≤1.37; 1.24–1.37), A. joshua (≤1.37; 1.19–1.37), and A. paloma (≤1.31; 1.21–1.31); by possessing a larger L4 scopulation extent (34%–48%) than A. paloma (5%–24%) and smaller than A. eutylenum (62%–77%). Certain morphometrics have potential to be useful, though due to the amounts of variation, small number of specimens, and the small differences between species, no other are claimed to be significant at this time (see Suppl. material
Acknowledgements
This is contribution #722 of the Auburn UniversityMuseum of Natural History (AUMNH). Funding was provided by the National Science Foundation (DEB-0841610 to JEB and BEH; Doctoral Dissertation Improvement Grant - DEB1311494 to CAH); Auburn University Department of Biological Sciences and College of Sciences and Mathematics (JEB); Millsaps College (BEH); the American Tarantula Society (BEH); the American Museum of Natural History (Theodore Roosevelt Memorial Grant to CAH); the American Arachnological Society (The Vincent Roth Fund for Systematic Research to CAH); and an Auburn University Cellular and Molecular Biosciences Peaks of Excellence Research Fellowship (CAH).
Specimens collected from National Park Service lands were permitted by: DEVA-2010-SCI-0012 (Death Valley National Park); JOTR-2008-SCI-0017 and JOTR-2010-SCI-0015 (Joshua Tree National Park); LAME-2010-SCI-0008 (Lake Mead NRA); MOJA-2010-SCI-0032 (Mojave National Preserve); SAGU-2010-SCI-0009 (Saguaro National Park); ZION-2010-SCI-0014 (Zion National Park). Specimens collected from California and California State Parks lands were permitted by Dept. of Fish and Game scientific collecting permit #802021-05 and Dept. of Parks and Recreation #9-2195(08III2010). Specimens were collected from state land in Texas with permission from Texas Parks & Wildlife. Specimens collected from private lands with permission from the landowner. All habitat photos taken by CAH & BEH. All burrow photos taken by CAH. All morphology photos taken by CAH. All photos of live specimens taken by CAH, BEH, & JEB.
The authors would like to thank Tom Prentice, Wendell Icenogle, Andrew Smith, and David Moellendorf for passing on their vast knowledge of the group. We are grateful to the following individuals and institutions for specimens loans and access to their collections: Norm Platnick, curator emeritus, Division of Invertebrate Zoology,
References
- Agnarsson I, Maddison W, Aviles L (2007) The phylogeny of the social Anelosimus spiders (Araneae: Theridiidae) inferred from six molecular loci and morphology. Molecular Phylogenetics and Evolution 43: 833–851. doi: 10.1016/j.ympev.2006.09.011
- Ausserer A (1871) Beiträge zur Kenntniss der Arachniden-Familie der Territelariae Thorell (Mygalidae Autor). Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien 21: 117–224.
- Ausserer A (1875) Zweiter Beitrag zur Kenntniss der Arachniden-Familie der Territelariae Thorell (Mygalidae Autor). Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien 25: 125–206.
- Ayoub N, Garb J, Hedin M, Hayashi C (2007) Utility of the nuclear protein-coding gene, elongation factor-1 gamma (EF-1 [gamma]), for spider systematics, emphasizing family level relationships of tarantulas and their kin (Araneae: Mygalomorphae). Molecular Phylogenetics and Evolution 42: 394–409. doi: 10.1016/j.ympev.2006.07.018
- Barnosky AD, Matzke N, Tomiya S, Wogan GOU, Swartz B, Quental TB, Marshall C, McGuire JL, Lindsey EL, Maguire KC, Mersey B, Ferrer EA (2011) Has the Earth’s sixth mass extinction already arrived? Nature 470: 51–57. doi: 10.1038/nature09678
- Bertani R, Nagahama RH, Fukushima CS (2011) Revalidation of Pterinopelma Pocock 1901 with description of a new species and the female of Pterinopelma vitiosum (Keyserling 1891) (Araneae: Theraphosidae: Theraphosinae). Zootaxa 2814: 1–18. doi: 10.11646/zootaxa.2814.1.1
- Bond J (2012) Phylogenetic treatment and taxonomic revision of the trapdoor spider genus Aptostichus Simon (Araneae, Mygalomorphae, Euctenizidae). ZooKeys 252: 1–209. doi: 10.3897/zookeys.252.3588
- Bond JE, Opell B (2002) Phylogeny and taxonomy of the genera of southwestern North American Euctenizinae trapdoor spiders and their relatives (Araneae: Mygalomorphae, Cyrtaucheniidae). Zoological Journal of the Linnean Society, London136: 487–534. doi: 10.1046/j.1096-3642.2002.00035.x
- Bond JE, Sierwald P (2002) Cryptic speciation in the Anadenobolus excisus millipede species complex on the island of Jamaica. Evolution 56: 1123–1135. doi: 10.1111/j.0014-3820.2002.tb01426.x
- Bond JE, Beamer D (2006) A morphometric analysis of mygalomorph spider carapace shape and its efficacy as a phylogenetic character (Araneae). Invertebrate Systematics 20: 1–8. doi: 10.1071/IS05041
- Bond JE, Hedin M (2006) A total evidence assessment of the phylogeny of North American euctenizine trapdoor spiders (Araneae, Mygalomorphae, Cyrtaucheniidae) using Bayesian inference. Molecular Phylogenetics and Evolution 41: 70–85. doi: 10.1016/j.ympev.2006.04.026
- Bond JE, Stockman AK (2008) An integrative method for delimiting cohesion species: finding the population-species interface in a group of Californian trapdoor spiders with extreme genetic divergence and geographic structuring. Systematic Biology 57: 628–646. doi: 10.1080/10635150802302443
- Bond JE, Hendrixson BE, Hamilton CA, Hedin M (2012) A reconsideration of the classification of the spider infraorder Mygalomorphae (Arachnida: Araneae) based on three nuclear genes and morphology. PLoS ONE 7: e38753. doi: 10.1371/journal.pone.0038753
- Bond JE, Godwin RL (2013) Taxonomic revision of the trapdoor spider genus Eucteniza Ausserer (Araneae, Mygalomorphae, Euctenizidae). ZooKeys 356: 31–67. doi: 10.3897/zookeys.356.6227
- Bond JE, Garrison NL, Hamilton CA, Godwin RL, Hedin M, Agnarsson I (2014) Phylogenomics Resolves a Spider Backbone Phylogeny and Rejects a Prevailing Paradigm for Orb Web Evolution. Current Biology 24(15): 1765–1771. doi: 10.1016/j.cub.2014.06.034
- Brusca RC, Wiens JF, Meyer WM, Eble J, Franklin K, Overpeck JT, Moore W (2013) Dramatic response to climate change in the Southwest: Robert Whittaker’s 1963 Arizona Mountain plant transect revisited. Ecology and Evolution 3: 3307–3319. doi: 10.1002/ece3.720
- Chamberlin RV (1940) New American Tarantulas of the Family Aviculariidae. Bulletin of the University of Utah 30: 1–39.
- Chamberlin RV, Ivie W (1939) New Tarantulas from the Southwestern States. Bulletin of the University of Utah 29: 1–17.
- Collen B, Böhm M, Kemp R, Baillie JEM (2012) Spineless: status and trends of the world’s invertebrates. Zoological Society of London, London, 86 pp.
- Cooke JAL, Roth VD, Miller FH (1972) The urticating hairs of theraphosid spiders. American Museum Novitates, 1–43.
- Coronado Planning Partnership (2008) State of the Coronado National Forest: An Assessment and Recommendations for the 21st Century. http://www.skyislandaction.org/SIAC-Library/state_of_the_coronado/State-of-the-Coronado.pdf
- Coyle FA (1995) A revision of the funnelweb mygalomorph spider subfamily Ischnothelinae (Araneae, Dipluridae). Bulletin of the American Museum of Natural History 226: 1–133.
- De Vos JM, Joppa LN, Gittleman JL, Stephens PR, Pimm SL (2014) Estimating the normal background rate of species extinction. Conservation Biology 29: 452–462. doi: 10.1111/cobi.12380
- Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research 32: 1792–1797. doi: 10.1093/nar/gkh340
- Funk DJ, Omland KE (2003) Species-level paraphyly and polyphyly: Frequency, causes, and consequences, with insights from animal mitochondrial DNA. Annual Review of Ecology, Evolution, and Systematics 34: 397–423. doi: 10.1146/annurev.ecolsys.34.011802.132421
- Gillespie RG, Croom HB, Palumbi SR (1994) Multiple origins of a spider radiation in Hawaii. PNAS 91: 2290–2294. doi: 10.1073/pnas.91.6.2290
- Girard C (1852) Arachnidians. In: Marcy R (Ed.) Natural History of the Red River of Louisiana. Washington, 32nd Congress, 2nd Session, Senate Appendix F (Zoology), 262–271. [reprinted twice in 1854: 33rd Congress, 1st Session, House of Representatives, p. 233–242; and 33rd Congress, 1st Session, Senate, 251–261]
- Goloboff PA (1993) A reanalysis of Mygalomorph spider families (Araneae). American Museum Novitates, 1–32.
- Goloboff PA, Mattoni CI, Quinteros AS (2006) Continuous characters analyzed as such. Cladistics 22: 589–601. doi: 10.1111/j.1096-0031.2006.00122.x
- Graham MR, Hendrixson BE, Hamilton CA, Bond JE (2015) Miocene extensional tectonics explain ancient patterns of diversification among turret-building tarantulas (Aphonopelma mojave group) in the Mojave and Sonoran deserts. Journal of Biogeography 42(6): 1052–1065. doi: 10.1111/jbi.12494
- Hamilton CA, Formanowicz DR, Bond JE (2011) Species delimitation and phylogeography of Aphonopelma hentzi (Araneae, Mygalomorphae, Theraphosidae): cryptic diversity in North American tarantulas. PLoS ONE 6: e26207. doi: 10.1371/journal.pone.0026207
- Hamilton CA, Hendrixson BE, Brewer MS, Bond JE (2014) An evaluation of sampling effects on multiple DNA barcoding methods leads to an integrative approach for delimiting species: A case study of the North American tarantula genus Aphonopelma (Araneae, Mygalomorphae, Theraphosidae). Molecular Phylogenetics and Evolution 71: 79–93. doi: 10.1016/j.ympev.2013.11.007
- Hanken J, Wake DB (1993) Miniaturization of body size: organismal consequences and evolutionary significance. Annual Review of Ecology, Evolution, and Systematics 24: 501–519. doi: 10.1146/annurev.es.24.110193.002441
- Hedin M (1997) Molecular phylogenetics at the population/species interface in cave spiders of the southern Appalachians (Araneae: Nesticidae: Nesticus). Molecular Biology and Evolution 14: 309–324. doi: 10.1093/oxfordjournals.molbev.a025766
- Hedin M, Bond J (2006) Molecular phylogenetics of the spider infraorder Mygalomorphae using nuclear rRNA genes (18S and 28S): conflict and agreement with the current system of classification. Molecular Phylogenetics and Evolution 41: 454–471. doi: 10.1016/j.ympev.2006.05.017
- Hendrixson B, Bond J (2005) Testing species boundaries in the Antrodiaetus unicolor complex (Araneae: Mygalomorphae: Antrodiaetidae). Molecular Phylogenetics and Evolution 36: 405–416. doi: 10.1016/j.ympev.2005.01.021
- Hendrixson BE, Bond JE (2009) Evaluating the efficacy of continuous quantitative characters for reconstructing the phylogeny of a morphologically homogeneous spider taxon (Araneae, Mygalomorphae, Antrodiaetidae, Antrodiaetus). Molecular Phylogenetics and Evolution 53: 300–313. doi: 10.1016/j.ympev.2009.06.001
- Hendrixson BE, DeRussy BM, Hamilton CA, Bond JE (2013) An exploration of species boundaries in turret-building tarantulas of the Mojave Desert (Araneae, Mygalomorphae, Theraphosidae, Aphonopelma). Molecular Phylogenetics and Evolution 66: 327–340. doi: 10.1016/j.ympev.2012.10.004
- Hendrixson BE, Guice AV, Bond JE (2015) Integrative species delimitation and conservation of tarantulas (Araneae, Mygalomorphae, Theraphosidae) from a North American biodiversity hotspot. Insect Conservation and Diversity 8: 120–131. doi: 10.1111/icad.12089
- Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965–1978. doi: 10.1002/joc.1276
- ICZN (1991) Opinion 1637. Bulletin of Zoological Nomenclature 48: 166–167.
- Jung AKS (1975) Morphological relationships among five species of sympatric tarantulas (Araneae, Theraphosidae), with descriptions of four new species. Masters Thesis, The University of Florida, Gainesville, Florida.
- Katoh K, Standley DM (2013) MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Molecular Biology and Evolution 30: 772–780. doi: 10.1093/molbev/mst010
- Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowitz S, Duran C, Thierer T, Ashton B, Meintjes P, Drummond A (2012) Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28: 1647–1649. doi: 10.1093/bioinformatics/bts199
- Knowles LL, Carstens BC (2007) Delimiting species without monophyletic gene trees. Systematic Biology 56: 887–895. doi: 10.1080/10635150701701091
- Knowles LL, Kubatko LS (2010) Estimating Species Trees – Practical and Theoretical Aspects. Wiley & Sons, Hoboken, 215 pp.
- Lanfear R, Calcott B, Kainer D, Mayer C, Stamatakis A (2014) Selecting optimal partitioning schemes for phylogenomic datasets. BMC Evolutionary Biology 14: 1–14. doi: 10.1186/1471-2148-14-82
- Leavitt DJ, Fitzgerald LA (2013) Disassembly of a dune-dwelling lizard community due to landscape fragmentation. Ecosphere 48(8): 1–15. doi: 10.1890/es13-00032.1
- Lemmon AR, Emme SA, Lemmon EM (2012) Anchored Hybrid Enrichment for massively high-throughput phylogenomics. Systematic Biology 61: 717–726. doi: 10.1093/sysbio/sys049
- Lienau EK, Desalle R, Rosenfeld J, Planet P (2006) Reciprocal illumination in the gene content Tree of Life. Systematic Biology 55: 441–453. doi: 10.1080/10635150600697416
- Locke SA, McLaughlin DJ, Marcogliese DJ (2010) DNA barcodes show cryptic diversity and a potential physiological basis for host specificity among Diplostomoidea (Platyhelminthes: Digenea) parasitizing freshwater fishes in the St. Lawrence River, Canada. Molecular Ecology 19: 2813–2827. doi: 10.1111/j.1365-294X.2010.04713.x
- Maddison W (1997) Gene Trees in Species Trees. Systematic Biology 46: 523–536. doi: 10.1093/sysbio/46.3.523
- Maddison WP, Maddison DR (2015) Mesquite: a modular system for evolutionary analysis. Version 3.03. http://mesquiteproject.org
- Masta S, Longhorn S, Boore J (2009) Arachnid relationships based on mitochondrial genomes: asymmetric nucleotide and amino acid bias affects phylogenetic analyses. Molecular Phylogenetics and Evolution 50: 117–128. doi: 10.1016/j.ympev.2008.10.010
- Meyer M, Kircher M (2010) Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harbor Protocols 6: 1–10. doi: 10.1101/pdb.prot5448
- Mirarab S, Reaz R, Bayzid MdS, Zimmerman T, Swenson MS, Warnow T (2014) ASTRAL: genome-scale coalescent-based species tree estimation. Bioinformatics 30: i541–i548. doi: 10.1093/bioinformatics/btu462
- Mitchell SG, Ober KA (2013) Evolution of Scaphinotus petersi (Coleoptera: Carabidae) and the role of climate and geography in the Madrean sky islands of southeastern Arizona, USA. Quaternary Research 79: 274–283. doi: 10.1016/j.yqres.2012.11.001
- Moore W, Meyer WM, Eble JA, Franklin K, Wiens JF, Brusca RC (2013) Introduction to the Arizona Sky Island Arthropod Project (ASAP): systematics, biogeography, ecology and population genetics of the Madrean Sky Islands. In: Gottfried GJ, Folliott PF, Gebow BS, Eskew LG, Collins LC (Eds) Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III. Tucson, AZ, May 2012. USDA Forest Service Proceedings RMRS-P-67, Fort Collins, Colorado, 144–168.
- Mora C, Tittensor DP, Adl S, Simpson AGB, Worm B (2011) How many species are there on Earth and in the ocean? PLoS Biology 9(8): e1001127. doi: 10.1371/journal.pbio.1001127
- Murphey PC, Guralnick RP, Glaubitz R, Neufeld D (2004) Georeferencing of museum collections: A review of problems and automated tools, and the methodology developed by the Mountain and Plains Spatio-Temporal Database-Informatics Initiative (Mapstedi). Phyloinformatics 3: 1–29.
- Niemiller ML, Near TJ, Fitzpatrick BM (2011) Delimiting species using multilocus data: diagnosing cryptic diversity in the southern cavefish, Typhlichthys subterraneus (Teleostei: Amblyopsidae). Evolution 66: 846–866. doi: 10.1111/j.1558-5646.2011.01480.x
- Padial J, Miralles A, de La Riva I, Vences M (2010) The integrative future of taxonomy. Frontiers in Zoology 7(16): 1–14. doi: 10.1186/1742-9994-7-16
- Peters H-J (2003) Tarantulas of the World: Amerika’s Vogelspinnen. Published by the author, Wegberg, 328 pp.
- Peters H-J (2005) Tarantulas of the world: Kleiner Atlas der Vogelspinnen - Band 3. Published by the author, Wegberg, 130 pp.
- Pérez-Miles F (1992) Revisión del género Eupalaestrus Pocock 1901 (Araneae, Theraphosidae). Revista Brasileira de Biologia 52: 27–35.
- Pérez-Miles F, Lucas SM, da Silva PI, Bertani R (1996) Systematic revision and cladistic analysis of Theraphosinae (Araneae - Theraphosidae). Mygalomorph 1: 33–68.
- Petit R, Excoffier L (2009) Gene flow and species delimitation. Trends In Ecology and Evolution 24: 386–393. doi: 10.1016/j.tree.2009.02.011
- Petrunkevitch A (1939) Status of the genus Eurypelma (Order Araneae, Family Theraphosidae). Annals and Magazine of Natural History 4: 561–568. doi: 10.1080/00222933908655402
- Phillips S, Anderson R, Schapire R (2006) Maximum entropy modeling of species geographic distributions. Ecological Modeling 190: 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
- Pickard-Cambridge FO (1897) Arachnida - Araneida and Opiliones. Biologia Centrali-Americana, Zoology, London 2: 1–40.
- Prentice TR (1993) A new species of North American tarantula, Aphonopelma paloma (Araneae, Mygalomorphae, Theraphosidae). Journal of Arachnology 20: 189–199.
- Prentice TR (1997) Theraphosidae of the Mojave Desert west and north of the Colorado River (Araneae, Mygalomorphae, Theraphosidae). Journal of Arachnology 25: 137–176.
- Pocock RI (1901) Some new and old genera of South American Avicularidae. Annals and Magazine of Natural History 8: 540–555. doi: 10.1080/03745480109443359
- Pyron R, Burbrink F (2010) Hard and soft allopatry: physically and ecologically mediated modes of geographic speciation. Journal of Biogeography 37(10): 2005–2015.
- de Queiroz K (2005) A Unified Concept of Species and its consequences for the future of taxonomy. Proceedings of the California Academy of Sciences 56: 196–215.
- Raven RJ (1985) The spider infraorder Mygalomorphae (Araneae) - Cladistics and Systematics. Bulletin of the American Museum of Natural History 182: 1–184.
- Raven RJ (1990) Comments on the proposed precedence of Aphonopelma Pocock, 1901 (Arachnida, Araneae) over Rhechostica Simon, 1892. Bulletin of Zoological Nomenclature 47: 126–127.
- Roewer CF (1942) Katalog der Araneae von 1758 bis 1940. Bremen, 1040 pp.
- Rokyta DR, Lemmon AR, Margres MJ, Aronow K (2012) The venom-gland transcriptome of the eastern diamondback rattlesnake (Crotalus adamanteus). BMC Genomics 13: 1–23. doi: 10.1186/1471-2164-13-312
- Rubinoff D, Cameron S, Will K (2006) A genomic perspective on the shortcomings of mitochondrial DNA for “barcoding” identification. Journal of Heredity 97: 581–594. doi: 10.1093/jhered/esl036
- Rudloff J-P (2008) Eine neue Brachypelma-Art aus Mexiko (Araneae: Mygalomorphae: Theraphosidae: Theraphosinae). Arthropoda 16(2): 26–30.
- Satler JD, Carstens BC, Hedin M (2013) Multilocus species delimitation in a complex of morphologically conserved trapdoor spiders (Mygalomorphae, Antrodiaetidae, Aliatypus). Systematic Biology 62: 805–823. doi: 10.1093/sysbio/syt041
- Simon E (1891) Liste des espéces de la famille des Aviculariidae qui habitent le Mexique et l’Amérique du Nord. Actes de la Société Linnéenne de Bordeaux 44: 307–326.
- Sites J, Marshall J (2004) Operational criteria for delimiting species. Annual Review of Ecology, Evolution, and Systematics 35: 199–227. doi: 10.1146/annurev.ecolsys.35.112202.130128
- Smith AM (1995) Tarantula Spiders - Tarantulas of the USA and Mexico. Fitzgerald Publishing, London, 196 pp.
- Smith AM (2010) Taxonomic and nomenclatural problems concerning the historical North and Central American theraphosid genera Aphonopelma Pocock 1901, Dugesiella Pocock 1901, Rhechostica Simon 1892, Delopelma Petrunkevitch 1939, Chaunopelma Chamberlin 1940, Clavopelma Chamberlin 1940, Gosipelma Chamberlin 1940 and their likely placement in future North American systematic studies. Journal of the British Tarantula Society 25: 129–138.
- Stamatakis A (2006) RAxML-VI-HPC: Maximum Likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688–2690. doi: 10.1093/bioinformatics/btl446
- Thorell T (1869) On European spiders. Part I. Review of the European genera of spiders, preceded by some observations on zoological nomenclature. Nova Acta Regiae Societatis Scientiarum Upsaliensis 7: 1–108.
- Thorell T (1870) On European spiders. Nova Acta Regiae Societatis Scientiarum Upsaliensis 7: 109–242.
- Turner SP, Longhorn SJ, Hamilton CA, Gabriel R, Perez-Miles F, Vogler AP (in review) Molecular phylogeny and conservation of New World tarantulas.
- Valerio CE (1980) Arañas terafósidas de Costa Rica (Araneae: Theraphosidae). III. Sphaerobothria, Aphonopelma, Pterinopelma, Citharacanthus, Crypsidromus y Stichoplastus. Revista de Biología Tropical 28: 271–296.
- Van Devender TR (1990) Late Quaternary vegetation and climate of the Chihuahuan Desert, United States and Mexico. In: Betancourt JL, Van Devender TR, Martin PS (Eds) Packrat Middens: the Last 40,000 Years of Biotic Change. University of Arizona Press, Tucson, 104–133.
- Warriner M (2008) Distribution and taxonomic status of tarantulas in Arkansas (Theraphosidae: Aphonopelma). Journal of the Arkansas Academy of Science 62: 107–114.
- Wilson JS, Gunnell CF, Wahl DB, Pitts JP (2013) Testing the species limits of the tarantulas (Araneae: Theraphosidae) endemic to California’s Southern Coast Ranges, USA. Insect Conservation and Conservation 6: 365–371. doi: 10.1111/icad.12000
- World Spider Catalog (2015) Natural History Museum Bern. http://wsc.nmbe.ch, version 16 [accessed 03/2015]
Supplementary materials
Compressed file (.zip) containing two separate locality datasets of Aphonopelma specimens.
Data type: Microsoft Excel spreadsheet (.xls) and Microsoft Excel comma delimited (.csv).
Explanation note: Locality data for all Aphonopelma specimens examined over the course of this study and listed in the material examined section that accompanies each species, as well as locality data with Darwin Core headers.
Compressed files (.zip) containing Aphonopelma boxplot and PCA morphospace data.
Data type: Microsoft Excel spreadsheet (.xls), Microsoft Excel comma delimited (.csv), Portable Document Format (.pdf), Simple Text (.txt), and Graphics Interchange Format (.gif).
Explanation note: All boxplots and PCA morphospace files used in Aphonopelma species boundary determination.
Compressed file (.zip) containing simple R scripts for analyzing Aphonopelma morphological space.
Data type: R scripts (.R).
Explanation note: R scripts to run boxplot, 2D and 3D PCA morphospace analyses of Aphonopelma morphological space. These scripts will allow the user to include their own specimen morphological data to compare with known Aphonopelma species morphological space. We are hoping to provide a mechanism for species identification based on specimen data from natural history collections or specimens where molecular data is not possible or practical in gathering. Directions for using the scripts are included within the scripts.
Compressed files (.zip) containing supplemental morphology figures.
Data type: Microsoft Word (.docx), Portable Document Format (.pdf), and Simple Text (.txt).
Explanation note: Figures (like those provided in the species descriptions) of additional specimens (for species where additional photographs were available), to provide visual reference of the variation possible within Aphonopelma species.