





Citation: Constantinescu IC, Chişamera G, Mukhim DKB, Adam C (2014) A new species of pterodectine feather mites (Acarina, Analgoidea, Proctophyllodidae) from the Little Spiderhunter Arachnothera longirostra (Passeriformes, Nectariniidae) in Meghalaya, India. ZooKeys 425: 1–13. doi: 10.3897/zookeys.425.7587
The article describes a new species of the feather mite subfamily Pterodectinae from the Little Spiderhunter Arachnothera longirostra Temminck, 1826 (Passeriformes, Nectariniidae) in India. Pedanodectes angustilobus sp. n. differs from all known Pedanodectes species by having opisthosomal lobes short, at base wider than long, roughly rounded apically in males, and strongly elongated and narrowed lobar region with wide terminal appendages in females. A key to species of the genus Pedanodectes is presented.
Pterodectinae, Pedanodectes angustilobus, new species, systematics
Feather mites are commensals or ectoparasites permanently living on birds. In India, the diversity of feather mites is poorly investigated, and only 26 species have been mentioned so far in various taxonomic papers (
In this paper, we describe a new species of Pedanodectes found on Arachnothera longirostra in India and we present a key for all species of this genus.
The material used in the present paper was collected in Meghalaya (India), in January 2014. The birds were trapped by means of ornithological mist nets, identified and visually checked for the presence of mites and, after mites were collected, released back to the wild. Mite specimens were placed in tubes with 95% ethanol. Later, in the laboratory, the mite specimens were cleared in lactic acid and mounted on microscope slides in Hoyer’s medium. Drawings were made using an Olympus CX21 microscope, using a camera lucida drawing device.
The bird specimens were identified according to
The genus currently includes six species, associated with birds of the order Passeriformes (families Nectariniidae, Malaconotidae, Cisticolidae and Platysteiridae) in Africa (see Table 1).
Pedanodectes species and their host associations.
| Mite species | Host species | Host family | Location | References |
|---|---|---|---|---|
| Pedanodectes hologaster (Gaud, 1953) | Chalcomitra senegalensis (Linnaeus) |
Nectariniidae | Central African Republic | |
| Pedanodectes andrei (Till, 1954) | Tchagra senegalus (Linnaeus) | Malaconotidae | Mozambique | |
| Pedanodectes mesocaulus (Gaud & Mouchet, 1957) | Deleornis fraseri cameroonensis (Bannerman) | Nectariniidae | Cameroon | |
| Pedanodectes marginatus Mironov & Kopij, 1997 | Camaroptera brachyura (Vieillot) | Cisticolidae | South Africa | |
| Pedanodectes latior Mironov & Kopij, 1997 | Platysteira peltata Sundevall | Platysteiridae | South Africa | |
| Pedanodectes blaszaki Mironov, 2008 | Cinnyris cupreus (Shaw) | Nectariniidae | South Africa | |
| Pedanodectes angustilobus sp. n. | Arachnothera longirostra (Latham) | Nectariniidae | India | Present paper |
* - Type host
http://zoobank.org/186A8195-8BE0-4212-A864-0FB3A224DC66
Male holotype (ANA256), 3 male (ANA257, ANA258, ANA259) and 4 female (ANA260, ANA261, ANA262, ANA263) paratypes from the Little Spiderhunter Arachnothera longirostra Temminck, 1826 (Passeriformes, Nectariniidae); INDIA: Meghalaya, Jaintia Hills, Shnongrim village, (25°21'12.36"N, 92°31'3.06"E); 1151 m alt; 24.01.2014, collector Costică Adam.
MALE (Figs 1A, B; 3A–D; holotype, range for 3 paratypes in parentheses): Length of idiosoma 316 (308–317), width 100 (100–104), length of hysterosoma 212 (207–212). Prodorsal shield divided into two parts by transversal band of soft tegument bearing setae se and si, antero-lateral extensions short and rounded, posterior margin slightly convex in median part, total length of shield 102 (102–106), greatest width 90 (84–90), surface without ornamentation (Fig. 1A). Scapular setae se separated by 38 (36–40). Humeral shields absent, setae cp situated ventrally, setae c2 situated dorsally, in anterior angles of hysteronotal shield. Subhumeral setae c3 lanceolate, 20 (19–20) × 6 (5–6). Length of hysteronotal shield from anterior margin to bases of setae h2 195 (199–203), greatest width in anterior part 88 (80–86), anterior margin concave, anterior angles acute, surface without ornamentation with strongly sclerotized transverse fold between h1 setae. Opisthosomal lobes short, at base wider than long, roughly rounded apically, lobar apices bearing setae h2. Terminal cleft almost semicircular, with narrow membranous margin in anterior part, length of terminal cleft 10 (12–13). Supranal concavity absent. Hysteronotal setae c1, d1, e1 absent; setae h3 narrowly lanceolate, length 24 (24–25), greatest width 6 (4–6); setae h2 represented by macrosetae, length 130 (132–140), greatest width 5 (4–5); setae ps2 slightly thickened, 18 (12–16) long; setae ps1 filiform, minute, length 6 (5–6), situated slightly anterior to bases of setae h3 and h2, approximately equidistant from inner and outer margins of opisthosomal lobe. Dorsal measurements: se-c2 72 (66–70), c2-d2 92 (90–104), d2-e2 68 (60–66), e2-h3 32 (32–34), h1-h3 10 (10–12), h2-h2 38 (38–42), h3-h3 26 (26–28), ps2-ps2 50 (46–49). Epimerites I fused into a Y, sternum about ½ of the total length of epimerites, posterior end of sternum with pair of postero-lateral extensions not connected to epimerites II. Epimerites II short, not extending to level of sejugal furrow. Coxal fields I–III open, without wide sclerotized areas. Epimerites IVa long, their anterior ends extending to midlevel of epimerites IV, their basal parts connected by semicircular transverse sclerite (supposedly genital shield) and almost completely encircling genital apparatus (Fig. 1B). Genital arch 11 (11–12) long, 14 (13–14) wide, basal sclerite of genital apparatus large, poorly sclerotized; aedeagus long, extending almost to anterior margin of terminal cleft, length of aedeagus from its anterior bend to tip 74 (72–78). Genital papillae indistinct, adanal shields absent. Anal suckers 11 (11–12) in diameter, corolla without indentations. Opisthoventral shields occupying lateral margin of opisthosoma, with narrow inner projection bearing seta ps3. Ventral measurements: 3a-4b 22 (25–30), 4b-4a 50 (44–50), 4a-g 24 (24–28), g-ps3 32 (28–31), ps3-ps3 40 (32–38), ps3-h3 24 (23–24).
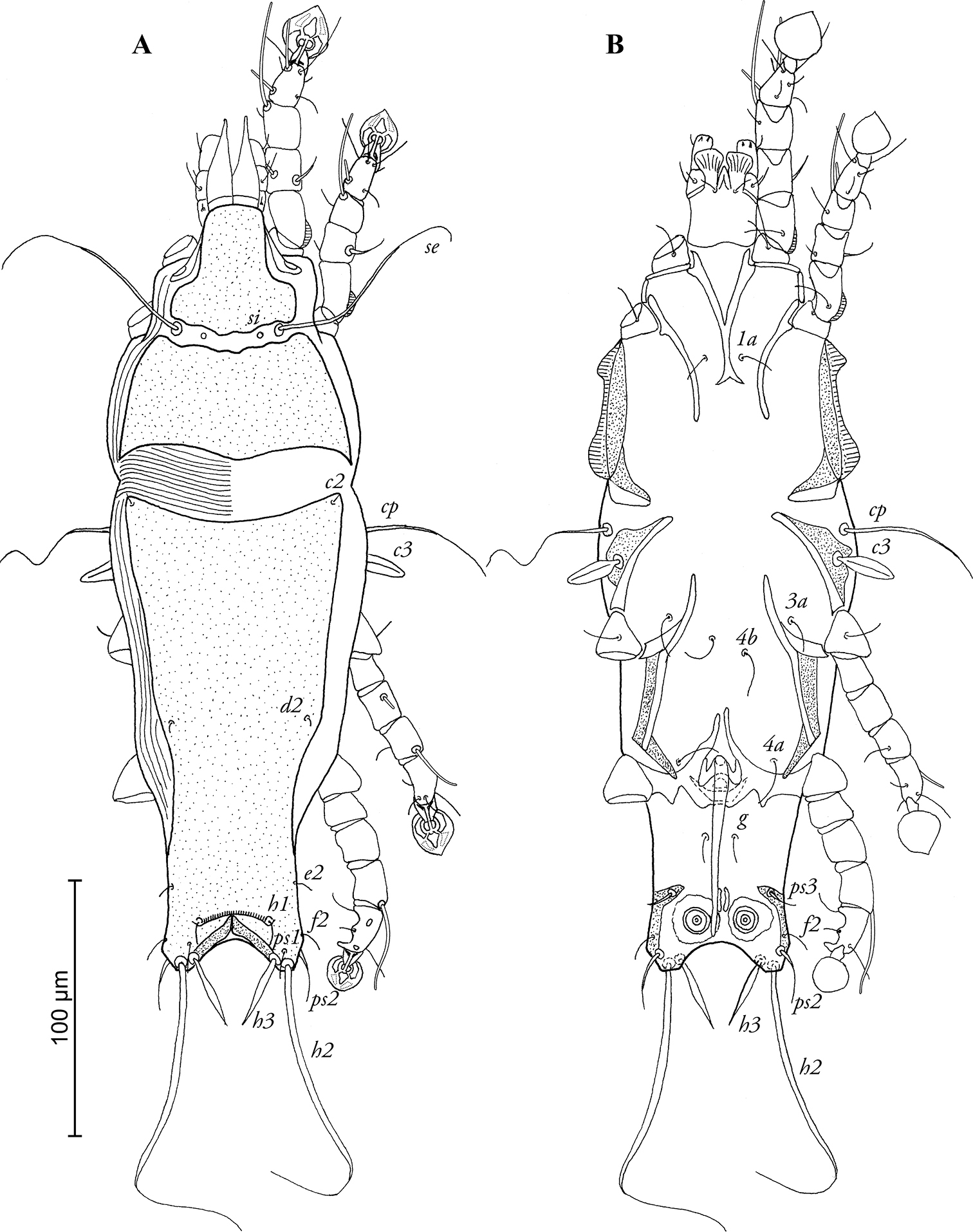
Pedanodectes angustilobus sp. n., male holotype: A dorsal view of idiosoma B ventral view of idiosoma.
Legs I slightly longer and thicker than legs II, femora I and II with ventro-basal crests (Fig. 3A, B). Seta mGII strongly thickened in basal half. Tarsus IV 18 (19–22) long, with apical claw and with small apico-ventral extensions bearing seta r and w; setae d, e button-like, seta d bigger than e, situated in basal and apical parts of segment, respectively (Fig. 3D). Length of solenidia: ω1I 15 (14–15), ω1II 12 (13–16), φI 42 (34–46), φII 29 (32–34), φIII 22 (22–24), φIV 32 (28–32).
FEMALE (Figs 2A, B; 4A–E; range for 4 paratypes): Length of idiosoma excluding terminal appendages 436–456, width 118–124, and length of hysterosoma 312–340. Prodorsal shield divided into two parts by transversal band of soft tegument, bearing setae se and si, antero-lateral extensions short and angular, posterior margin concave, total length of shield 118–122, greatest width 104–112, surface without ornamentation (Fig. 2A). Scapular setae se separated by 48–50. Humeral shields absent, setae cp situated ventrally, setae c2 situated dorsally, in anterior angles of hysteronotal shield. Subhumeral setae c3 lanceolate, 17–20 × 5–6. Hysteronotal shield divided into anterior hysteronotal shield and lobar shield. Anterior hysteronotal shield roughly rectangular, with anterior margin concave, greatest length 204–208, greatest width in anterior part 100–106, surface without ornamentation. Lobar shield elongated and narrowed, with well-developed lateral extensions bearing setae h2, length of lobar shield 86–92, width at level of setae h2 58–62. Terminal cleft narrow, almost parallel-sided, with narrow elliptical part in anterior third, length 72–76, greatest width 3–4. Supranal concavity absent. Terminal appendages wide, their width in basal half similar to that of lobes. Hysteronotal setae c1, d1, e1 absent; setae h1 situated on lobar shield at level of anterior end of terminal cleft, setae h2 spindle-shaped, without terminal filaments, 38–40 × 6–9. Setae ps1 situated approximately equidistant from outer and inner margins of opisthosomal lobes, setae h3 6–8 long. Dorsal measurements: se-c2 88–94, c2-d2 100–112, d2-e2 86–94, e2-h2 36–44, h2-h3 48–52, h1-h2 16–22, h1-h1 18–22, h2-h2 36–40, h3-h3 20–24. Epimerites I fused as a Y, posterior end of sternum with small rounded lateral extensions not reaching epimerites II. Coxal fields I, II open, without heavily sclerotized areas, outer margins of epimerites I and II with narrow sclerotized areas (Fig. 2B). Epimerites IVa rudimentary. Translobar apodemes of opisthosomal lobes present, fused to each other anterior to terminal cleft. Epigynum horseshoe-shaped, outer margin with lateral extensions, greatest width 52–60. Head of spermatheca as in Fig. 4E, primary spermaduct with three different enlargements: ball-like enlargement near very head of spermatheca; moderate enlargement with verrucous external surface in proximal 1/5 of this duct; conical enlargement at copulatory opening. Secondary spermaducts short, not longer that ball-like enlargement of primary spermaduct. Copulatory opening situated ventral, posterior to anal opening. Distance between pseudanal setae: ps2-ps2 24–286, ps3-ps3 24–29, ps2-ps3 10–14.
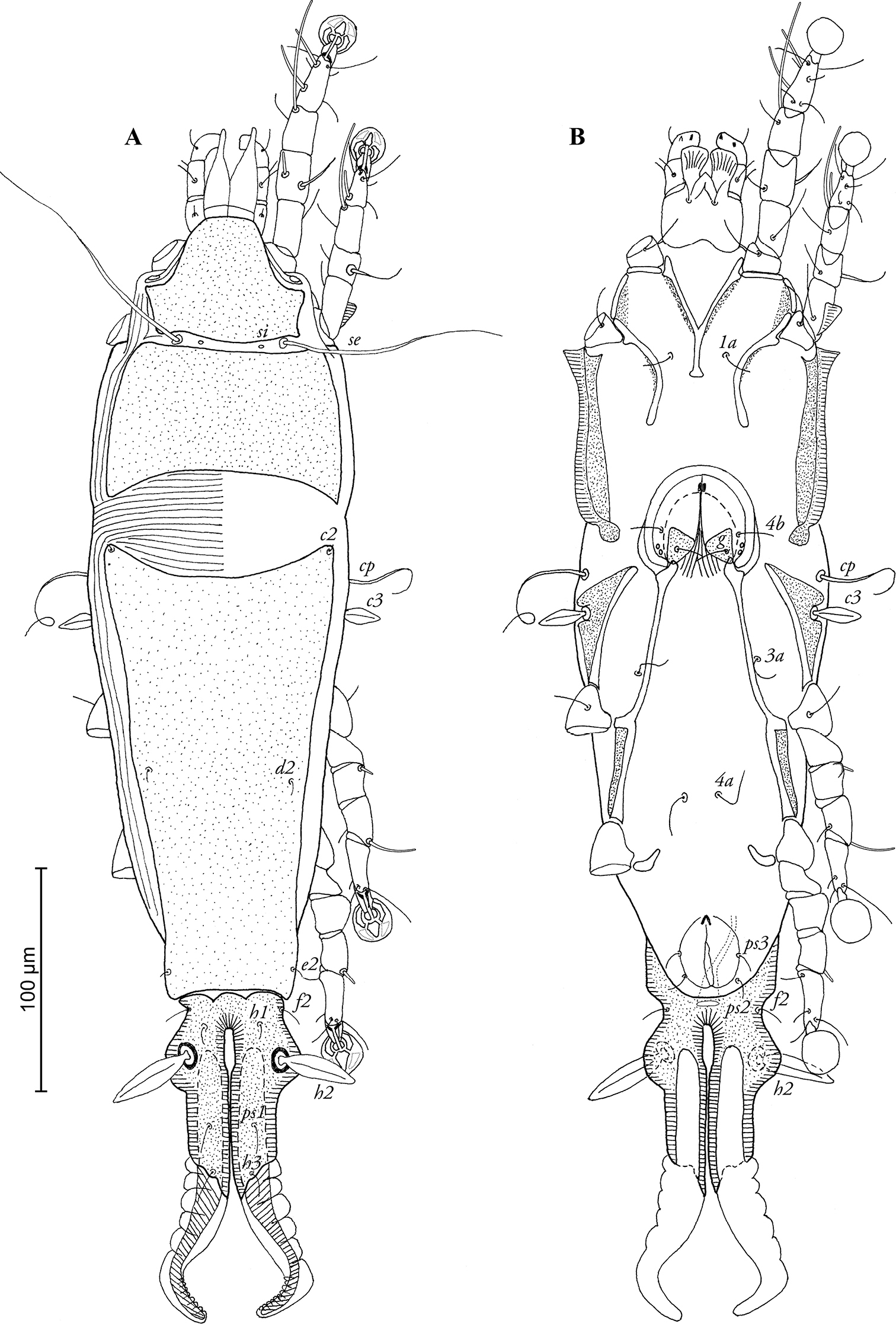
Pedanodectes angustilobus sp. n., female paratype: A dorsal view of idiosoma B ventral view of idiosoma.
Legs I longer and thicker than legs II, femora II with ventro-basal crests, genua III with dorso-basal crest (Fig. 4A–C). Genual setae mGI and mGII noticeably thickened in basal half. Length of solenidia: ω1I 17–18, ω1II 13–16, φI 45–56, φII 32–38, φIII 22–27, φIV 7–8.
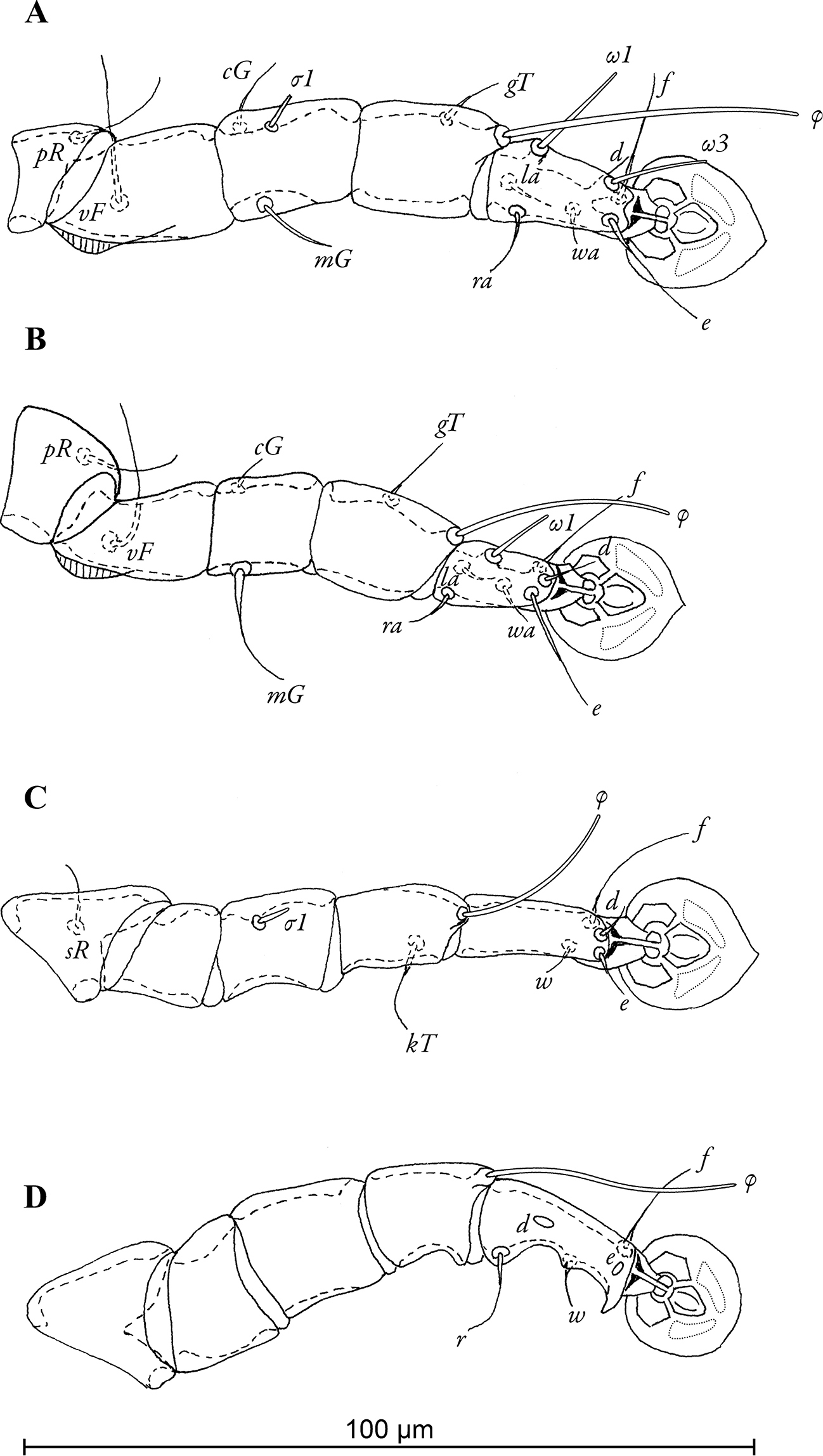
Pedanodectes angustilobus sp. n., details: A–D legs I–IV of male, respectively, dorsal view.
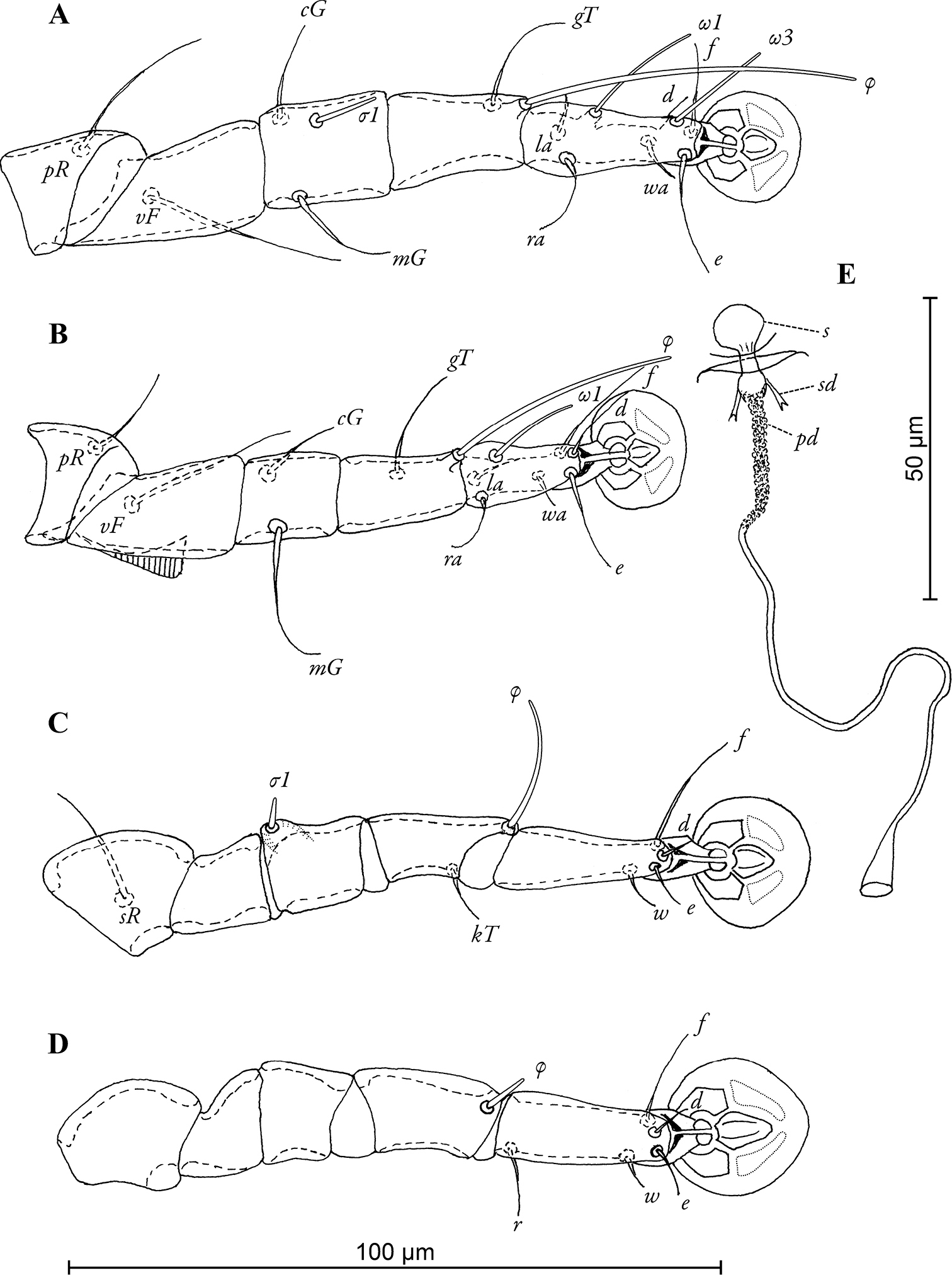
Pedanodectes angustilobus sp. n., details: A–D legs I–IV of female, respectively E spermatheca and spermaducts, dorsal view. Abbreviations: pd - primary spermaduct; s - spermatheca; sd - secondary spermaduct.
The specific epithet refers to the narrowed lobar region of the female, and it is an adjective in the nominative singular.
Pedanodectes angustilobus sp. n. clearly differs from all previously described Pedanodectes species because its males have well expressed opisthosomal lobes, and females have elongated and narrowed lobar region, with wide terminal appendages. Among previously known species of the genus, males of Pedanodectes angustilobus sp. n. appear to be closest to that of Pedanodectes mesocaulus (Gaud & Mouchet, 1957) from Deleornis fraseri cameroonensis (Bannerman, 1921) (Passeriformes: Nectariniidae). Males of both species have setae ps1 situated antero-lateral to the adanal suckers, epimerites I fused into Y, similar shape of opisthoventral shields and epimerites IVa with narrow anterior projections. Males of Pedanodectes angustilobus sp. n. are easily to distinguish from those of Pedanodectes mesocaulus by the following features: epimerites IVa have free the anterior projections and are connected posterior to the genital arch by a transversal sclerite, the tip of aedeagus does not extend beyond the posterior margin of idiosoma. In males of Pedanodectes mesocaulus, the anterior projections of epimerites IVa are fused forming a pregenital sclerite while the transverse sclerite connecting the bases of these epimerites is absent, the tip of aedeagus extends beyond the posterior margin of idiosoma. Females of the new species are clearly different from those of the other species of the genus by the following unique combination of characters: the lobar region has the same width in anterior and posterior part; the terminal cleft is parallel-sided, with the margins almost touching, except for the anterior one third; and the terminal appendages are thick, their basal half is approximately as wide as opisthosomal lobes. In females of the other Pedanodectes species, the lobar region in the anterior part is wider than in the posterior part and the terminal appendages are narrower than lobes. The terminal cleft in females of the other species has the following shape: with the lateral margins parallel and almost touching in Pedanodectes andrei and Pedanodectes mesocaulus, with lateral margins sinuous and almost touching in certain parts in Pedanodectes marginatus and Pedanodectes latior; as a narrow inverted V in Pedanodectes blaszaki, and as a narrow inverted U in Pedanodectes hologaster.
According to the diagnosis of the genus Pedanodectes, some authors considered that males of this genus practically have no opisthosomal lobes and setae ps3 are usually situated lateral or postero-lateral to the adanal suckers (
| 1 | Epimerites I fused V-likely | 2 |
| – | Epimerites I fused Y-likely | 4 |
| 2 | Postero-lateral extensions of epimerites I connected to epimerites II | Pedanodectes latior |
| – | Postero-lateral extensions of epimerites I not connected to epimerites II | 3 |
| 3 | Tip of aedeagus extending beyond posterior margin of idiosoma | Pedanodectes andrei |
| – | Tip of aedeagus not extending beyond posterior margin of idiosoma | Pedanodectes hologaster |
| 4 | Postero-lateral extensions of epimerites I present | 5 |
| – | Postero-lateral extensions of epimerites I absent | Pedanodectes blaszaki |
| 5 | Postero-lateral extensions of epimerites I connected to epimerites II | Pedanodectes marginatus |
| – | Postero-lateral extensions of epimerites I not connected to epimerites II | 6 |
| 6 | Opisthosomal lobes present; tip of aedeagus not extending beyond posterior margin of idiosoma | Pedanodectes angustilobus sp. n. |
| – | Opisthosomal lobes absent; tip of aedeagus extending beyond posterior margin of idiosoma | Pedanodectes mesocaulus |
| 1 | Width of terminal appendages in anterior half similar to that of opisthosomal lobes | Pedanodectes angustilobus sp. n. |
| – | Terminal appendages narrower than opisthosomal lobes | 2 |
| 2 | Lateral margins of terminal cleft spaced in some parts or on their entire length | 3 |
| – | Lateral margins of terminal cleft almost touching on their entire length | 6 |
| 3 | Prodorsal shield split into anterior and posterior pieces at level of scapular setae | Pedanodectes blaszaki |
| – | Prodorsal shield entire | 4 |
| 4 | Lateral margins of prodorsal shield entire | 5 |
| – | Lateral margins of prodorsal shield with deep incisions around setae se | Pedanodectes latior |
| 5 | Epimerites I fused V-likely | Pedanodectes hologaster |
| – | Epimerites I fused Y-likely | Pedanodectes marginatus |
| 6 | Posterior margin of anterior hysteronotal shield straight | Pedanodectes andrei |
| – | Posterior margin of anterior hysteronotal shield concave | Pedanodectes mesocaulus |
We are grateful to the Additional Principal Chief Conservator of Forests, Wildlife & Chief Wildlife Warden from Shillong (Meghalaya, India) for the permission to catch birds (permission No. FWC.G/173/Pt.). We would like to thank reviewers for their patience in analyzing the manuscript and their valuable suggestions that have greatly improved the paper, and also our proofreader, PhD. Ana Wetzl (Assistant Professor of English, Kent State University at Trumbull, USA).