





Citation: Adams BJ, Wall DH, Virginia RA, Broos E, Knox MA (2014) Ecological Biogeography of the Terrestrial Nematodes of Victoria Land, Antarctica. ZooKeys 419: 29–71. doi: 10.3897/zookeys.419.7180
The terrestrial ecosystems of Victoria Land, Antarctica are characteristically simple in terms of biological diversity and ecological functioning. Nematodes are the most commonly encountered and abundant metazoans of Victoria Land soils, yet little is known of their diversity and distribution. Herein we present a summary of the geographic distribution, habitats and ecology of the terrestrial nematodes of Victoria Land from published and unpublished sources. All Victoria Land nematodes are endemic to Antarctica, and many are common and widely distributed at landscape scales. However, at smaller spatial scales, populations can have patchy distributions, with the presence or absence of each species strongly influenced by specific habitat requirements. As the frequency of nematode introductions to Antarctica increases, and soil habitats are altered in response to climate change, our current understanding of the environmental parameters associated with the biogeography of Antarctic nematofauna will be crucial to monitoring and possibly mitigating changes to these unique soil ecosystems.
Biodiversity, dispersal, climate change, Eudorylaimus, freeliving nematodes, Geomonhystera, habitat suitability, invasive species, Panagrolaimus, Plectus, Scottnema, soil
Understanding the global distribution of biodiversity is critical for studying the evolution, ecology and dynamics of ecosystems and to address how global scale changes in climate, invasive species, and land use will affect ecosystems, ecosystem services, and subsequently, people. Antarctic terrestrial ecosystems might seem less sensitive to global change because this polar desert has low species diversity distributed across a limited area of biologically active ice-free land, comprising less than 0.32% of the continent’s 14 million km2 (
Aboveground, the diversity and biogeography of terrestrial flora (mosses, lichens and liverworts) has been recently assessed and used to further refine the geographic floral regions of Antarctica (
Nematoda are a major component of soil food webs in all terrestrial ecosystems including the exposed lands of Antarctica, though their spatial distribution and abundance are highly heterogeneous. In more productive ecosystems, they typically have much higher diversity (
In Antarctica, soil nematodes have been studied primarily in localized and easily accessible areas largely centered around research bases and concentrated on the Antarctic peninsula and islands of the maritime Antarctic and further south in ice-free areas. As a consequence there is relatively little known of their regional biogeography or of the habitats that are suitable for functioning communities. Additionally, there are many remote inland ice-free areas which have yet to be sampled (
Regional to continental-scale descriptions of the Antarctic nematofauna have pointed to a paucity of distributional records for much of the continent (
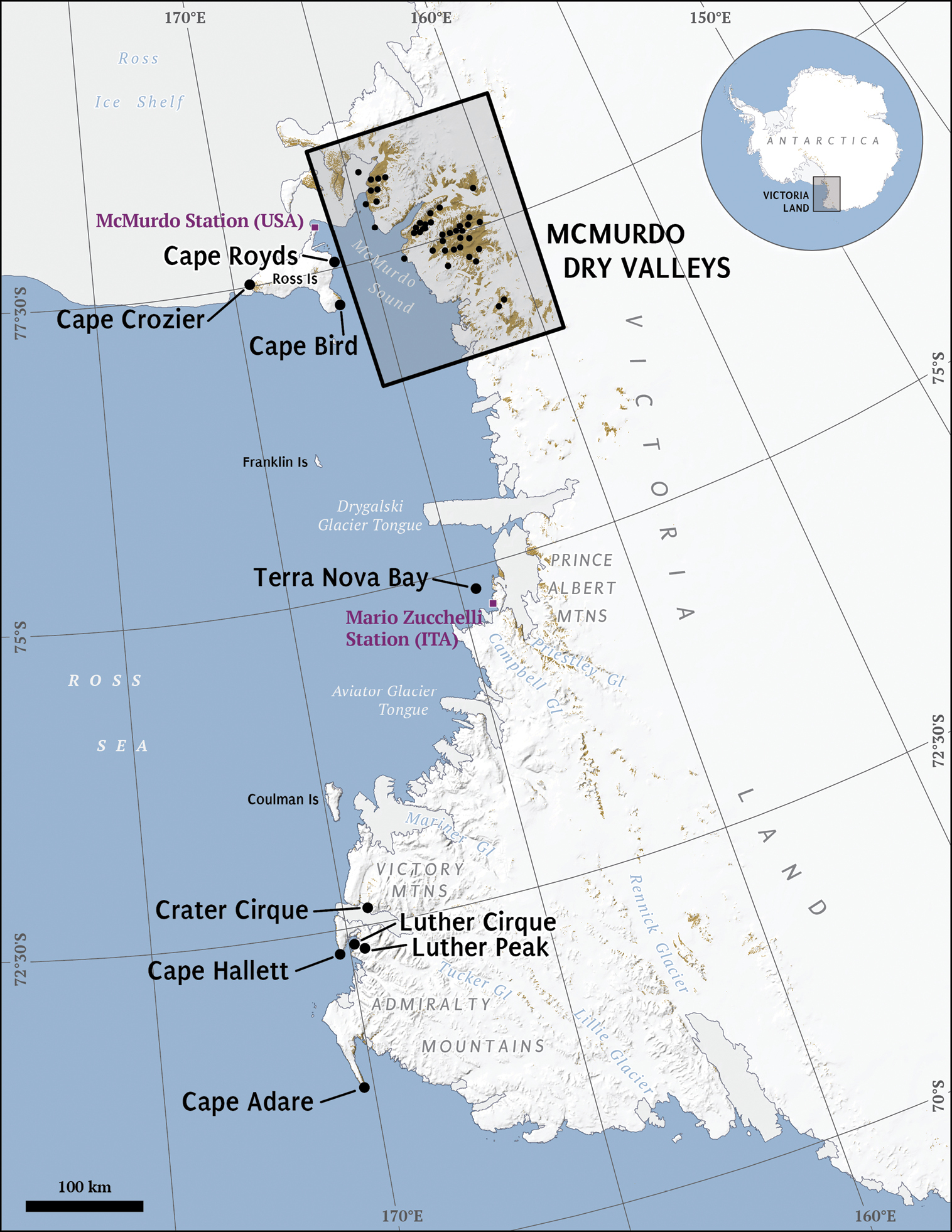
Victoria Land, Antarctica. Labeled areas represent study locations and major geographic features referenced in the tables and text. Box inset of the McMurdo Dry Valleys is rotated 180 °and expanded in Figure 2.

McMurdo Dry Valleys, Antarctica. Labeled areas represent study locations and major geographic features referenced in the tables and text.
The McMurdo Dry Valleys (76°5'to 78°5'S, 160°0' to 164°0'E) are located along the TransAntarctic Mountains in Southern Victoria Land and comprise about 4, 800 km2 of ice-free land and have different geo/ecological legacies and climatic conditions (
The distributions of the Dry Valley metazoan species are associated with specific sites and correlate to soil habitat differences in organic matter content, moisture and salinity, and microclimate differences encountered over environmental gradients of coastal to interior sites, latitude, and soil chronosequences and differences in glacial tills (
Coastal areas of Victoria Land are a moister environment than the Dry Valleys and are habitat for birds and marine mammals (e.g. skua gulls, penguins, and seals). Penguin rookeries are associated with ornithogenic soils with significant inputs of carbon and nitrogen transferred from the marine environment to the soil (
Each of the unique soil ecosystems of Victoria Land imposes considerable physiological constraints on nematode life history traits, requiring adaptive responses to freeze/thaw cycling, osmotic and desiccation stress, and a short growing season (
Nematodes were first collected in Victoria Land by the British ‘Discovery’ expedition of 1901-1903, from Discovery Bay, South Victoria Land and described by
In the McMurdo Dry Valley Region, most nematological studies have investigated the diversity, ecology and distribution patterns of up to three nematode genera; Eudorylaimus, Plectus, Scottnema (
Based on published and unpublished data, we summarized biogeographic information on the species represented within each nematode genus described in Victoria Land. In addition to published papers, we present information obtained from data on soil, and lake and stream sediment samples collected throughout Victoria Land, by the authors and team members during the austral summers between and including 1990 and 2004. Data referred to as “this study (year)” were derived from nematode soil extraction procedures optimized for Antarctic soils and all nematodes were identified to species (
Biogeographic distribution of Scottnema lindsayae in Victoria Land, Antarctica. NP = not published. NA = specific coordinates not available within the named locale identified above. For references to “this paper”, the year collected refers to the year at the beginning of the austral summer in which samples were collected at 0-10 cm depth unless otherwise indicated. For abundance, ‘Present’ indicates no abundance information available, Low = >0 to 20 nematodes per kg dry soil, M-low = 21 to 200 nematodes per kg dry soil, Medium = 201 to 600 nematodes per kg dry soil, M-high = 601 to 1000 nematodes per kg dry soil, High = 1001 to 2000 nematodes per kg dry soil, V-high = >2000 nematodes per kg dry soil, n = number of samples and % = percentage of samples in which Scottnema lindsayae occurred. AThere may have been a typographical error in the original publication reporting this latitude/longitude. *Geographic coordinates associated with the recognized Antarctic place name for a general feature as listed by the USGS Advisory Committee on Antarctic Names (http://geonames.usgs.gov/antarctic/) and updated by the Polar Geospatial Center (http://www.pgc.umn.edu).
| Biogeographic location | Lat, Long | Habitat | Abundance | Reference | |
|---|---|---|---|---|---|
| Victoria Land | *74°15.00'S, 163°00.00'E | ||||
| - | NP, NP | “river”, wet mosses | Present | ( |
|
| McMurdo Dry Valleys | *77°30.00'S, 162°00.00'E | ||||
| - | NP, NP | soil | Present | ( |
|
| - | NP, NP | soil | M-high | ( |
|
| - | NP, NP | soil | Present | ( |
|
| Alatna Valley | *76°52.82'S, 161°13.82'E | ||||
| East, middle and south west end | NA, NA | soil | Medium (n = 20, 40%) | This paper, collected in 1995 | |
| Battleship Promontory | *76°54.85'S, 160°59.34'E | ||||
| - | NA, NA | soil | Medium (n = 17, 88%) | This paper, collected in 1993 | |
| - | 76°55.30'S, 161°04.79'E | soil | M-high (n = 9, 22%) | This paper, collected in 1994 | |
| - | NA, NA | soil | Medium (n = 6, 83%) | This paper, collected in 1996 | |
| - | 76°52.00'S, 161°05.00'E | soil | Present | ( |
|
| Southwestern Bluff | 76°55.00'S, 161°03.00'E | soil | Medium (n = 14, 57%) | This paper, collected in 2001 | |
| - | 76°55.30'S, 161°04.22'E | soil | Medium (n = 6, 83%) | This paper, collected in 2003 | |
| Barwick Valley | *77°20.71'S, 161°06.09'E | soil | Medium (n = 10, 40%) | This paper, collected in 1994 | |
| Beacon Valley | *77°49.00'S, 160°39.00'E | soil | Low (n = 24, 4%) | This paper, collected in 1990 | |
| Garwood Valley | *78°02.00'S, 164°10.00'E | ||||
| - | NA, NA | soil | High (n = 6, 100%) | This paper, collected in 1993 | |
| - | 78°02.00'S, 164°10.00'E | soil | Medium | ( |
|
| - | 78°02.00'S, 164°10.00'E | soil | Present | ( |
|
| - | NA, NA | soil | V-high (n = 13, 100%) | This paper, collected in 2002 | |
| McKelvey Valley | *77°26.00'S, 161°33.00'E | ||||
| Upper | NA, NA | soil | High (n = 18, 50%) | This paper, collected in 1990 | |
| Lower | NA, NA | soil | Low (n = 14, 14%) | This paper, collected in 1990 | |
| Miers Valley | *78°06.00'S, 164°00.00'E | ||||
| Miers Glacier | *78°05.00'S, 163°40.00'E | moraine | Present | ( |
|
| - | NA, NA | soil | M-high (n = 24, 88%) | This paper, collected in 1990 | |
| Taylor Valley | *77°38.82'S, 163°03.08'E | ||||
| Canada Glacier | *77°37.00'S, 162°59.00'E | soil | Present | ( |
|
| Lacroix Glacier | *77°40.00'S, 162°33.00'E | ||||
| Between Lake Bonney and Lacroix Glacier | NP, NP | small runoff stream | Present | ( |
|
| Southeast of Lacroix Glacier | *77°40.00'S, 162°30.00'E | sandy soil | Present | ( |
|
| Lake Bonney | *77°43.00'S, 162°25.00'E | ||||
| South side East Lobe | 77°42.92'S, 162°27.65'E | soil | Low (n = 9, 29%) | This paper, collected in 1993 | |
| NA, NA | soil polygons | Medium (n = 99, 64%) | This paper, collected in 1994 | ||
| - | NA, NA | soil | Medium (n = 2, 50%) | This paper, collected in 1995 | |
| - | 77°42.92'S, 162°27.65'E | soil | Low | ( |
|
| - | NP, NP | soil polygons | Medium | ( |
|
| South side West Lobe | 77°42.5'S, 162°31.2'E | soil | Medium (n = 18, 94%) | This paper, collected in 1999, 2001 and 2002 ( |
|
| - | 77°42.92'S, 162°27.65'E | soil | Present | ( |
|
| - | NA, NA | soil and stream sediment | Medium (n = 20, 45%) | This paper, collected in 2000 | |
| South side West Lobe | NA, NA | soil | M-low (n = 96, 45%) | This paper, collected in 2000, 2002 and 2003 | |
| South side East Lobe | 77°42.55'S, 162°27.39'E | soil | Low | ( |
|
| Lake Chad | *77°38.55'S, 162°45.70'E | soil | Medium (n = 9, 22%) | This paper, collected in 1995 | |
| - | 77°38.10'S, 162°48.15'E | soil | Present | ( |
|
| Lake Fryxell | *77°36.58'S, 163°09.10'E | ||||
| - | NA, NA | soil | Medium (n = 26, 23%) | This paper, collected in 1990 | |
| South side | 77°35.94'S, 163°22.68'E | soil | V-high (n = 9, 100%) | This paper, collected in 1993 | |
| - | 77°35.94'S, 163°22.68'E | soil | High (n = 10, 80%) | This paper, collected in 1993 | |
| - | NA, NA | soil | High (n = 102, 87%) | This paper, collected in 1994 | |
| - | NA, NA | soil | Medium (n = 4, 75%) | This paper, collected in 1995 | |
| - | 77°35.94'S, 163°22.68'E | soil | V-high | ( |
|
| Von Guerard (F6) stream | 77°36.49'S, 163°14.95'E | soil | V-high (n = 30, 100%) | This paper, collected in 1996, 1997, 1998, 2001 and 2003 | |
| - | NA, NA | soil | V-high (n = 5, 100%) | This paper, collected in 1997 | |
| Von Guerard Stream/ Harnish Creek network | *77°37.00'S, 163°15.00'E | soil and stream sediments | Medium | ( |
|
| - | NP, NP | soil polygons | High | ( |
|
| Huey Creek stream | *77°36.00'S, 163°06.00'E | soil | Medium (n = 7, 29%) | This paper, collected in 1999 | |
| Von Guerard stream | *77°37.00'S, 163°15.00'E | soil | High (n = 8, 100%) | This paper, collected in 1999 | |
| South side F6 stream | 77°36.49'S, 163°14.92'E | soil | V-high (n = 12, 100%) | This paper, collected in 1999 and 2001 | |
| - | A77°55.94'S, 163°22.68'E | soil | Present | ( |
|
| Harnish Creek transect | *77°37.00'S, 163°13.00'E | soil and stream sediment | M-high (n = 20, 90%) | This paper, collected in 2000 | |
| South side by F6 (SF) | NA, NA | soil | V-high (n = 96, 100%) | This paper, collected in 2000, 2002 and 2003 | |
| South-east shore | 77°35.56'S, 163°22.41'E | soil | V-high | ( |
|
| - | 77°36.00'S, 162°15.00'E | soil | V-high | ( |
|
| South side near F6 | 77°36.40'S, 163°15.30'E | soil and lake sediment | High (n = 12, 67%) | This paper, collected in 2002 | |
| South side near Green Creek | 77°37.36'S, 163°03.91'E | soil | M-high (n = 20, 85%) | This paper, collected in 2003 | |
| South side near F6 | 77°36.72'S, 163°15.18'E | soil | High (n = 20, 90%) | This paper, collected in 2003 | |
| Von Guerard stream | 77°37.00'S, 163°15.00'E | soil | High | ( |
|
| Green Creek | 77°37.36'S, 163°03.91'E | soil | M-High | ( |
|
| Lake Hoare | *77°38.00'S, 162°51.00'E | ||||
| North side | 77°37.49'S, 162°54.31'E | soil | M-low (n = 18, 100%) | This paper, collected in 1993 | |
| South side | 77°38.03'S, 162°52.75'E | soil | High (n = 9, 100%) | This paper, collected in 1993 | |
| South side | 77°37.59'S, 162°52.57'E | soil | High (n = 56, 100%) | This paper, collected in 1993, 1994, 1995, 1996, 1997 and 2001 | |
| North side | 77°38.00'S, 162°53.00'E | soil (0-2.5, 2.5-5, 5-10, 10-20 cm | High | ( |
|
| South shore | NP, NP | soil at varying elevation | Medium | ( |
|
| - | NA, NA | soil polygons | High (n = 104, 96%) | This paper, collected in 1994 | |
| South side | 77°38.02'S, 162°52.23'E | soil | High (n = 40, 83%) | This paper, collected in 1994, 1995, 1996, 1997 and 2001 | |
| North side | 77°38.00'S, 162°53.00'E | soil | Medium | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil at varying elevation | M-high | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil (0-2.5, 2.5-5, 5-10, 10-20 cm) | Medium | ( |
|
| South side | NA, NA | soil polygons | M-high (n = 24, 100%) | This paper, collected in 1995 | |
| North side | 77°37.49'S, 162°54.31'E | soil | M-low | ( |
|
| South side | 77°38.03'S, 162°52.75'E | soil | M-high | ( |
|
| - | NP, NP | soil | Medium | ( |
|
| South side | NA, NA | soil | M-high (n = 12, 100%) | This paper, collected in 1997 | |
| South side | 77°38.00'S, 162°53.00'E | soil | Medium | ( |
|
| - | NP, NP | soil polygons | High | ( |
|
| North side | NA, NA | soil | V-high (n = 8, 100%) | This paper, collected in 1999 | |
| South side | NA, NA | soil | M-high (n = 8, 100%) | This paper, collected in 1999 | |
| South side | 77°38.07'S, 162°52.59'E | soil | M-high (n = 18, 100%) | This paper, collected in 1999, 2001 and 2002 | |
| North side | 77°37.49'S, 162°54.31'E | soil | Present | ( |
|
| South side | 77°38.03'S, 162°52.75'E | soil | Present | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil | M-high | ( |
|
| North side | 77°37.29'S, 162°54.19'E | soil | M-low | ( |
|
| South side | 77°38.02'S, 162°52.45'E | soil | M-high | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil | M-high | ( |
|
| - | 77°38.00'S, 162°53.00'E | soil | Medium | ( |
|
| - | 77°37.90'S, 162°53.20'E | soil and lake sediments | M-high (n = 11, 73%) | This paper, collected in 2002 | |
| - | NP, NP | soil | Present | ( |
|
| North side | NP, NP | lake bottom detritus | Present | ( |
|
| - | 77°37.00'S, 160°50.00'E | soil | Medium | ( |
|
| South side | NA, NA | wetlands (upland ponds) | M-low (n = 48, 19%) | This paper, collected in 2000 | |
| - | NP, NP | soil | High | ( |
|
| - | NP, NP | 0-5 cm soil (exposed and subnivian) | High | ( |
|
| South side | NP, NP | bare soil >80 m from wetlands (upland ponds) | Medium | ( |
|
| Mummy Pond | 77°40.06'S, 162°39.00'E | soil | Low (n = 5, 20%) | This paper, collected in 1997 | |
| Nussbaum Riegel | 77°38.52'S, 162°46.89'E | soil | V-High (n = 5, 20%) | This paper, collected in 1997 | |
| Victoria Valley | *77°23.00'S, 162°00.00'E | ||||
| Lower Victoria Valley | 77°21.81'S, 162°19.11'E | soil | High (n = 10, 60%) | This paper, collected in 1993 | |
| NP | 77°21.81'S, 162°19.11'E | soil | High (n = 9, 100%) | This paper, collected in 1993 | |
| Lake Vida | *77°23.29'S, 161°56.05'E | ||||
| - | NA, NA | soil | Medium (n = 16, 19%) | This paper, collected in 1990 | |
| - | 77°23.35'S, 162°02.60'E | soil | Medium (n = 10, 50%) | This paper, collected in 1993 | |
| - | NA, NA | soil | Medium (n = 11, 27%) | This paper, collected in 1994 | |
| - | 77°22.58'S, 161°13.56'E | soil | NA (n = 2, 100%) | This paper, collected in 2000 | |
| Vida Met Station | NA, NA | soil | Low (n = 4, 50%) | This paper, collected in 2002 | |
| - | NA, NA | soil | M-high (n = 10, 80%) | This paper, collected in 1997 | |
| - | 77°23.00'S, 162°00.00'E | soil | M-high | ( |
|
| - | NA, NA | soil | NA (n = 6, 83%) | This paper, collected in 2003 | |
| Victoria Lower Glacier | *77°18.00'S, 162°40.00'E | ||||
| - | 77°21.81'S, 162°19.11'E | soil | High | ( |
|
| - | 77°22.57'S, 162°13.56'E | soil | NA (n = 6, 83%) | This paper, collected in 2000 | |
| - | 77°21.81'S, 162°19.11'E | soil | Present | ( |
|
| South-west | 77°21.49'S, 162°19.07'E | soil | High | ( |
|
| Victoria Upper Glacier | *77°16.00'S, 161°25.00'E | ||||
| - | 77°17.35'S, 161°33.03'E | soil | High (n = 10, 60%) | This paper, collected in 1993 | |
| - | 77°17.35'S, 161°33.03'E | soil | Low (n = 9, 11%) | This paper, collected in 1993 | |
| Victoria Upper Lake | *77°19.00'S, 161°35.00'E | soil | M-high (n = 20, 35%) | This paper, collected in 1990 | |
| Wright Valley | *77°31.39'S, 161°58.70'E | ||||
| Dais | *77°33.00'S, 161°16.00'E | ||||
| - | NP, NP | soil | Present | ( |
|
| - | NA, NA | soil | NA (n = 3, 100%) | This paper, collected in 2000 | |
| East of Meserve Glacier | *77°31.00'S, 162°17.00'E | algal mat | NP | ( |
|
| Labyrinth | *77°33.00'S, 160°50.00'E | ||||
| West | 77°33.04'S, 160°43.15'E | soil | M-low (n = 9, 100%) | This paper, collected in 1993 | |
| - | 77°33.04'S, 160°43.15'E | soil | M-low (n = 9, 78%) | This paper, collected in 1993 | |
| - | 77°33.04'S, 160°43.15'E | soil | Low | ( |
|
| - | 77°31.00'S, 161°50.00'E | soil | M-low | ( |
|
| West | 77°33.02'S, 160°43.09'E | soil | Low | ( |
|
| - | NA, NA | soil | Low (n = 12, 17%) | This paper, collected in 2003 | |
| Lake Brownworth | *77°26.00'S, 162°45.00'E | ||||
| - | NP, NP | soil | Present | ( |
|
| - | 77°26.13'S, 162°42.61'E | soil | M-low (n = 9, 33%) | This paper, collected in 1993 | |
| - | 77°26.13'S, 162°42.61'E | soil | M-low | ( |
|
| - | 77°26.13'S, 162°42.61'E | soil | Present | ( |
|
| South-west shore | 77°26.08'S, 162°42.37'E | soil | M-low | ( |
|
| Met Station | NA, NA | soil | Medium (n = 4, 75%) | This paper, collected in 2002 | |
| Bull Pass | *77°28.00'S, 161°46.00'E | soil | Medium (n = 22, 23%) | This paper, collected in 1990 | |
| Lake Bull | *77°31.51'S, 161°42.68'E | soil | Low (n = 12, 17%) | This paper, collected in 1990 | |
| - | 77°28.00'S, 161°46.00'E | soil | High (n = 24, 33%) | ( |
|
| Lake Vanda | *77°32.00'S, 161°33.00'E | ||||
| Near Lake Vanda | 77°32.00'S, 161°33.00'E | soil | Present | ( |
|
| Vanda Station | 77°31.00'S, 161°40.00'E | soil | M-low (n = 2, 100%) | This paper, collected in 2002 | |
| Unspecified Locations | |||||
| - | NA, NA | soil | M-low (n = 5, 80%) | This paper, collected in 1997 | |
| - | NA, NA | soil | Present (n = 1, 100%) | This paper, collected in 2000 | |
| - | NA, NA | soil | Present (n = 10, 60%) | This paper, collected in 2003 | |
| Koettlitz Glacier and Southern Coastal Regions | *78°15.00'S, 164°15.00'E | ||||
| Péwé Lake | *77°56.67'S, 164°16.87'E | stony soil near the lake | Present | ( |
|
| Strand Moraines | *77°45.04'S, 164°29.90'E | sandy soil | Present | ( |
|
| Marble Point | *77°26.00'S, 163°50.00'E | mossy soil (Bryum antarcticum) | Present | ( |
|
| Northern Coastal Region | |||||
| Cape Hallett | 72°19.29'S, 170°13.52'E | soil | Low (n = 67, 56%) | ( |
|
| Crater Cirque | *72°37.49'S, 169°22.48'E | lake bottom detritus and wet mosses | Present | ( |
|
| Edmonson Point | *74°20.00'S, 165°08.00'E | ||||
| - | NA, NA | soil | Medium (n = 10, 80%) | This paper, collected in 1996 | |
| - | NA, NA | soil | Present (n = 28, 36%) | This paper, collected in 1996 | |
| - | NP, NP | soil | Present | ( |
|
| - | NA, NA | soil | Medium (n = 8, 63%) | This paper, collected in 2001 | |
| Gondwana Station | 74°37.57'S, 164°11.91'E | soil | M-Low (n = 371, 79%) | ( |
|
| Luther Peak | *72°21.88'S, 169°50.91'E | ||||
| Luther Cirque | 72°22.20'S, 169°53.10'E | soil | Medium (n = 40, 85%) | This paper, collected in 2003 | |
| Luther Vale North | 72°22.00'S, 169°53.00'E | soil | Medium | ( |
|
| Luther Vale South | 72°22.00'S, 169°53.00'E | soil | Medium | ( |
|
| Terra Nova Bay | *74°54.51'S, 164°27.19'E | ||||
| 600 km north and south of the Italian station | NP, NP | mosses, lichens, fresh water sediments and penguin excrements (there are no details of whether Scottnema lindsayae occurred in all habitats or only in some) | Present | ( |
|
| - | 74°20.00'S, 165°08.00'E | soil | Present | ( |
|
Biogeographic distribution of Plectus species in Victoria Land, Antarctica. NP = not published, NA = not available, mur = Plectus murrayi. frig = Plectus frigophilus, where both exist = spp. For abundance, A abundance is per kg moss and adhering rock fragments not soil, Low = >0 to 20 nematodes per kg dry soil, M-low = 21 to 200 nematodes per kg dry soil, Medium = 201 to 600 nematodes per kg dry soil, M-high = 601 to 1000 nematodes per kg dry soil, High = 1001 to 2000 nematodes per kg dry soil, V-high = >2000 nematodes per kg dry soil, n = number of samples and % = percentage of samples in which Plectus occurred. For references to “this paper”, the year collected refers to the year at the beginning of the austral summer in which samples were collected to 0-10 cm depth. BThis publication refers to a map for more details on sample location.
| Biogeographic location | Lat, Long | Habitat | Species | Abundance | Reference | |
|---|---|---|---|---|---|---|
| McMurdo Dry Valleys | *77°30.00'S, 162°00.00'E | |||||
| - | NP, NP | soil | spp. | Present | ( |
|
| - | NP, NP | soil | spp. | Low | ( |
|
| - | NP, NP | soil | mur | Present | ( |
|
| - | NP, NP | streams | spp. | Present | ( |
|
| Alatna Valley | *76°52.82'S, 161°13.82'E | |||||
| Battleship Promontory | *76°54.85'S, 160°59.34'E | soil | mur | Low (n = 17, 6%) | This paper, collected in 1993 | |
| East, middle and southwestern end | NA, NA | soil | mur | Low (n = 20, 10%) | This paper, collected in 1995 | |
| Garwood Valley | *78°02.00'S, 164°10.00'E | |||||
| Garwood Lake | *78°02.00'S, 164°15.00'E | NP | frig | Present | ( |
|
| - | NA, NA | soil | mur | M-low (n = 6, 50%) | This paper, collected in 1993 | |
| - | 78°02.00'S, 164°10.00'E | soil | mur | M-low | ( |
|
| - | NA, NA | soil | mur | Low (n = 13, 8%) | This paper, collected in 2002 | |
| Miers Valley | *78°06.00'S, 164°00.00'E | |||||
| Miers Glacier | *78°05.00'S, 163°40.00'E | mossy soil from glacier foot, runoff stream | frig | Present | ( |
|
| - | NA, NA | soil | mur | M-low (n = 24, 29%) | This paper, collected in 1990 | |
| Taylor Valley | *77°38.82'S, 163°03.08'E | |||||
| Canada Glacier | *77°37.00'S, 162°59.00'E | |||||
| Near the glacier | NP, NP | soil | frig | Present | ( |
|
| - | 77°37.31'S, 162°58.26'E | windblown sediment on top of glacier | mur | Present (n = 2, 100%) | This paper, collected in 1997 | |
| Waterfall (upper west) | NA, NA | cryconite hole | mur | Present | This paper, collected in 2001 | |
| Lake Bonney | *77°43.00'S, 162°25.00'E | |||||
| - | NP, NP | lake, soil nearby | frig | Present | ( |
|
| - | NA, NA | soil polygon cracks | mur | Low (n = 99, 5%) | This paper, collected in 1994 | |
| - | NA, NA | soil | mur | Medium (n = 2, 100%) | This paper, collected in 1995 | |
| - | NA, NA | algal mat | spp. | Present (n = 5, 100%) | This paper, collected in 1995 | |
| West Lobe | 77°43.50'S, 162°18.95'E | soil | mur | Low (n = 18, 33%) | This paper, collected in 1999, 2001 and 2002 | |
| - | NA, NA | soil and stream sediment | mur | M-low (n = 20, 30%) | This paper, collected in 2000 | |
| West Lobe | NA, NA | soil | mur | Low (n = 72, 7%) | This paper, collected in 2000 and 2003 | |
| - | 77°43.40'S, 162°18.40'E | soil and sediment | mur | Low (n = 12, 25%) | This paper, collected in 2002 | |
| Lake Chad | *77°38.55'S, 162°45.70'E | |||||
| - | NP, NP | NP | frig | Present | ( |
|
| - | NA, NA | algal mat | spp. | NA (n = 1, 100%) | This paper, collected in 1995 | |
| - | NA, NA | soil | mur | M-low (n = 9, 56%) | This paper, collected in 1995 | |
| Lake Fryxell | *77°36.58'S, 163°09.10'E | |||||
| - | NP, NP | NP | frig | Present | ( |
|
| - | NP, NP | algae in a drift stream near the lake | spp. | Present | ( |
|
| - | NA, NA | algal mat | ant | M-low (n = 10, 100%) | This paper, collected in 1990 | |
| - | NA, NA | soil | mur | M-high (n = 26, 77%) | This paper, collected in 1990 | |
| - | 77°35.94'S, 163°22.68'E | soil | mur | Low (n = 10, 10%) | This paper, collected in 1993 | |
| - | NA, NA | algal mat | spp. | NA (n = 1, 100%) | This paper, collected in 1995 | |
| - | NA, NA | soil | mur | Medium (n = 4, 75%) | This paper, collected in 1995 | |
| Von Guerard stream / Harnish Creek network | *77°37.00'S, 163°15.00'E | stream sediments and surrounding soils | spp. | M-low | ( |
|
| Huey Creek | *77°36.00'S, 163°06.00'E | soil | mur | M-low (n = 7, 57%) | This paper, collected in 1999 | |
| Harnish Creek | *77°37.00'S, 163°13.00'E | soil and sediment | mur | M-low (n = 20, 60%) | This paper, collected in 2000 | |
| South side | NA, NA | soil | mur | Low (n = 72, 4%) | This paper, collected in 2000 and 2002 | |
| South side | 77°36.40'S, 163°15.30'E | soil and sediment | mur | V-High (n = 12, 75%) | This paper, collected in 2002 | |
| South side | 77°36.49'S, 163°14.95'E | soil | mur | Low (n = 6, 17%) | This paper, collected in 2003 | |
| South side | 77°36.49'S, 163°14.92'E | soil | mur | Low (n = 6, 17%) | This paper, collected in 2003 | |
| South side near Green Creek | 77°37.36'S, 163°03.91'E | soil | mur | Medium (n = 20, 60%) | This paper, collected in 2003 | |
| Green Creek | 77°37.36'S, 163°03.91'E | soil | mur | Medium | ( |
|
| Lake Hoare | *77°38.00'S, 162°51.00'E | |||||
| North side | 77°37.49'S, 162°54.31'E | soil | mur | Low (n = 18, 6%) | This paper, collected in 1993 | |
| South side | NP, NP | soil at varying elevation | spp. | Low | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil (0-2.5, 2.5-5, 5-10, 10-20 cm) | mur | Low | ( |
|
| South side | 77°37.93'S, 162°53.19'E | soil at varying elevation | mur | Low (n = 150, 6%) | This paper, collected in 1995, 1998 and 2002 | |
| North side | 77°37.49'S, 162°54.31'E | soil | mur | Low | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil | mur | Low | ( |
|
| North side | NA, NA | soil | mur | M-low (n = 8, 50%) | This paper, collected in 1999 | |
| North side | 77°37.29'S, 162°54.19'E | soil | mur | Low | ( |
|
| South side | 77°38.02'S, 162°52.23'E | soil | mur | Low (n = 8, 13%) | This paper, collected in 2001 | |
| South side | 77°38.00'S, 162°53.00'E | soil | mur | Low | ( |
|
| - | 77°37.90'S, 162°53.20'E | soil and lake sediment | mur | V-High (n = 11, 82%) | This paper, collected in 2002 | |
| South side | 77°38.02'S, 162°53.05'E | soil | mur | Low (n = 6, 17%) | This paper, collected in 2003 | |
| - | 77°37.00'S, 162°50.00'E | soil | mur | M-low | ( |
|
| - | NP, NP | soil | mur | M-low | ( |
|
| South side | NA, NA | high elevation upland pond areas | spp. | Low (n = 48, 19%) | This paper, collected in 2000 | |
| - | NP, NP | 0-5 cm soil (subnivian) | mur | M-low | ( |
|
| South side | NP, NP | high elevation upland pond areas | spp. | M-low | ( |
|
| Taylor Glacier | *77°44.00'S, 162°10.00'E | windblown sediment on top of glacier | mur | Present (n = 1, 100%) | This paper, collected in 1998 | |
| Suess Glacier | *77°38.00'S, 162°40.00'E | soil nearby | frig | Present | ( |
|
| Suess Lake | NP, NP | NP | frig | Present | ( |
|
| Victoria Valley | *77°23.00'S, 162°00.00'E | soil | mur | Present (n = 6, 17%) | This paper, collected in 2003 | |
| Wright Valley | *77°31.39'S, 161°58.70'E | |||||
| Along Onyx River | *77°31.31'S, 161°49.39'E | pond | spp. | Present | ( |
|
| East of Meserve Glacier | *77°31.00'S, 162°17.00'E | algal mat | spp. | Present | ( |
|
| Canopus Pond | NP, NP | NP | frig | Present | ( |
|
| Lake Vanda | *77°32.00'S, 161°33.00'E | lake, soil nearby | frig | Present | ( |
|
| Lower Wright Lake (=Lake Brownworth) | *77°26.00'S, 162°45.00'E | NP | frig | Present | ( |
|
| Edge of Lake Canopus | *77°33.00'S, 161°31.00'E | algal growth at the edge of the lake | spp. | Present | ( |
|
| Between Lake Vanda and Lake Bull | NP, NP | dry algae around the edge of small ponds | spp. | Present | ( |
|
| Between Lake Vanda and Lake Bull | NP, NP | wet algae in meltwater and around the edge of small ponds | spp. | Present | ( |
|
| Bull Pass | *77°28.00'S, 161°46.00'E | soil | mur | M-low (n = 22, 18%) | This paper, collected in 1990 | |
| Lake Bull | *77°31.51'S, 161°42.68'E | soil | mur | Low (n = 12, 8%) | This paper, collected in 1990 | |
| - | 77°31.00'S, 161°50.00'E | soil | mur | M-low | ( |
|
| Koettlitz Glacier and Southern Coastal Regions | *78°15.00'S, 164°15.00'E | |||||
| Cape Chocolate (just north of) | *77°56.05'S, 164°34.70'E | moraine | frig | Present | ( |
|
| Marble Point | *77°26.00'S, 163°50.00'E | |||||
| - | NP, NP | moss (Bryum antarcticum) | mur | V-highA | ( |
|
| - | NP, NP | mossy soil and melt pools with abundant algae (Nostoc commune) | mur | Present | ( |
|
| - | NP, NP | meltpools w/ abundant algae (Nostoc commune), mossy soil | frig | Present | ( |
|
| Pewe Lake | NP, NP | NP | frig | Present | ( |
|
| Strand Moraines | *77°45.04'S, 164°29.90'E | |||||
| - | NP, NP | mossy soil and melt pools with abundant algae (Nostoc commune) | spp. | Present | ( |
|
| - | NP, NP | sandy soil, mossy soil, stream with abundant algae | frig | Present | ( |
|
| Northern Coastal Region | ||||||
| Cape Hallett | *72°19.00'S, 170°16.00'E | |||||
| - | NP, NP | NP | spp. | Present | ( |
|
| Willett Cove | 72°19.00'S, 170°14.00'E | soil | mur | Medium | ( |
|
| - | NA, NA | soil amongst penguin rookery | mur | M-low (n = 20, 30%) | This paper, collected in 2003 | |
| - | 72°19.29'S, 170°13.52'E | soil | Low (n = 67, 56%) | ( |
||
| Edmonson Point | *74°20.00'S, 165°08.00'E | |||||
| -B | NP, NP | wet moss near a brook | mur | Present | ( |
|
| - | NP, NP | soil | spp. | Present | ( |
|
| - | NA, NA | soil | mur | M-high (n = 10, 70%) | This paper, collected in 1996 | |
| - | NA, NA | soil | mur | NA (n = 28, 50%) | This paper, collected in 1996 | |
| - | NA, NA | soil | mur | M-low (n = 8, 63%) | This paper, collected in 2001 | |
| Gondwana Station | 74°37.57'S, 164°11.91'E | soil | Low (n = 371, 84%) | ( |
||
| Luther Peak | 72°22.20'S, 169°53.10'E | soil | mur | Low (n = 40, 8%) | This paper, collected in 2003 | |
| Luther Vale South | 72°22.00'S, 169°53.00'E | soil | mur | Low | ( |
|
| Terra Nova Bay | *74°54.51'S, 164°27.19'E | |||||
| 600 km north and south of the Italian station | NP, NP | mosses, lichens, fresh-water sediments and penguin excrements (no details of whether Plectus occurred in all habitats or only in some) | spp. | Present | ( |
|
| Barclay Glacier | ||||||
| - | NP, NP | algae growing in meltwater | mur | Present | ( |
|
Biogeographic distribution of Eudorylaimus species in Victoria Land, Antarctica. NP = not published. NA = not available. ant = Eudorylaimus antarcticus. gla = Eudorylaimus glacialis. For abundance, Low = >0 to 20 nematodes per kg dry soil, M-low = 21 to 200 nematodes per kg dry soil, Medium = 201 to 600 nematodes per kg dry soil, M-high = 601 to 1000 nematodes per kg dry soil, High = 1001 to 2000 nematodes per kg dry soil, V-high = >2000 nematodes per kg dry soil, n = number of samples and % = percentage of samples in which Eudorylaimus occurred. AThere may have been a typographical error in the original publication reporting this longitude. BASPA = Antarctic Specially Protected Area (previously Site of Special Scientific Interest). For references to “this paper”, the year collected refers to the year at the beginning of the austral summer in which samples were collected at 0-10 cm depth.
| Biogeographic location | Lat, Long | Habitat | Species | Abundance | Reference | |
|---|---|---|---|---|---|---|
| McMurdo Dry Valleys | *77°30.00'S, 162°00.00'E | |||||
| - | NP, NP | soil | ant | Present | ( |
|
| - | NP, NP | soil | ant, gla | M-low | ( |
|
| - | NP, NP | soil | ant | Present | ( |
|
| - | NP, NP | soil, sediment | ant | Present | ( |
|
| Alatna Valley | *76°52.82'S, 161°13.82'E | |||||
| Battleship Promontory | *76°54.85'S, 160°59.34'E | |||||
| - | NA, NA | soil | ant | M-low (n = 17, 65%) | This paper, collected in 1993 | |
| - | 76°55.30'S, 161°04.79'E | moist soil, green with algae and between dolomite rocks | ant | Low (n = 9, 22%) | This paper, collected in 1994 | |
| - | NA, NA | soil | ant | M-low (n = 6, 17%) | This paper, collected in 1996 | |
| Southwestern Bluff | 76°55.00'S, 161°03.00'E | soil | ant | Low (n = 14, 7%) | This paper, collected in 2001 | |
| - | NA, NA | soil | ant | Low (n = 6, 50%) | This paper, collected in 2003 | |
| Garwood Valley | *78°02.00'S, 164°10.00'E | |||||
| Garwood Lake | *78°01.58'S, 164°15.42'E | NP | ant | Present | ( |
|
| - | NA, NA | soil | ant | M-low (n = 6, 100%) | This paper, collected in 1993 | |
| - | 78°02.00'S, 164°10.00'E | soil | ant | M-low | ( |
|
| - | NA, NA | soil | ant | M-low (n = 13, 23%) | This paper, collected in 2002 | |
| McKelvey Valley | *77°26.00'S, 161°33.00'E | |||||
| Upper | NA, NA | soil | ant | M-low (n = 18, 33%) | This paper, collected in 1990 | |
| Miers Valley | *78°06.00'S, 164°00.00'E | |||||
| Miers Glacier (the foot of) | *78°05.00'S, 163°40.00'E | moss | ant | Present | ( |
|
| Runoff stream from the Miers Glacier | *78°05.00'S, 163°40.00'E | NP | ant | Present | ( |
|
| Miers Lake | *78°06.00'S, 163°51.00'E | NP | ant | Present | ( |
|
| - | NA, NA | soil | ant | M-low (n = 24, 50%) | This paper, collected in 1990 | |
| Taylor Valley | *77°38.82'S, 163°03.08'E | |||||
| Lake Bonney | *77°43.00'S, 162°25.00'E | |||||
| - | NP, NP | NP | ant | Present | ( |
|
| - | NA, NA | soil | ant | Low (n = 99, 52%) | This paper, collected in 1994 | |
| - | NA, NA | soil | ant | M-low (n = 2, 100%) | This paper, collected in 1995 | |
| - | NA, NA | algal mat | ant | NA (n = 5, 60%) | This paper, collected in 1995 | |
| - | NP, NP | soil, sediment | ant | Low | ( |
|
| West Lobe | 77°43.50'S, 162°18.95'E | soil | ant | Low (n = 18, 61%) | This paper, collected in 1999, 2001 and 2002 | |
| - | NA, NA | soil, sediment | ant | M-low (n = 20, 35%) | This paper, collected in 2000 | |
| West Lobe | NA, NA | soil | ant | Low (n = 48, 2%) | This paper, collected in 2000 | |
| Lake Chad | *77°38.55'S, 162°45.70'E | |||||
| - | NP, NP | NP | ant | Present | ( |
|
| - | NA, NA | soil | ant | M-low (n = 9, 56%) | This paper, collected in 1995 | |
| - | NA, NA | algal mat | ant | NA (n = 1, 100%) | This paper, collected in 1995 | |
| Lake Fryxell | *77°36.58'S, 163°09.10'E | |||||
| - | NP, NP | NP | ant | Present | ( |
|
| - | NP, NP | algae in a drift stream near the lake | ant | Present | ( |
|
| - | NA, NA | plant material | ant | Present (n = 10, 100%) | This paper, collected in 1990 | |
| - | NA, NA | soil | ant | Medium (n = 26, 77%) | This paper, collected in 1990 | |
| South side | 77°35.94'S, 163°22.68'E | soil | ant | Low (n = 9, 11%) | This paper, collected in 1993 | |
| - | NA, NA | soil | ant | Low (n = 102, 41%) | This paper, collected in 1994 | |
| South side | 77°36.49'S, 163°18.95'E | soil | ant | Low (n = 18, 33%) | This paper, collected in 1996, 1998 and 2001 | |
| - | NA, NA | soil | ant | Low (n = 5, 20%) | This paper, collected in 1997 | |
| - | NA, NA | algal mat | ant | Present (n = 1, 100%) | This paper, collected in 1998 | |
| - | NA, NA | soil | ant | Medium (n = 4, 75%) | This paper, collected in 1998 | |
| Von Guerard stream/ Harnish Creek | *77°37.00'S, 163°15.00'E | stream sediments and surrounding soils | ant, gla | Medium | ( |
|
| - | NP, NP | soil, sediment | ant | Low | ( |
|
| Von Guerard stream | *77°37.00'S, 163°15.00'E | soil | ant | M-low (n = 8, 63%) | This paper, collected in 1999 | |
| Huey Creek stream | *77°36.00'S, 163°06.00'E | soil | ant | M-low (n = 7, 29%) | This paper, collected in 1999 | |
| South side | 77°36.49'S, 163°14.92'E | soil | ant | M-low (n = 12, 83%) | This paper, collected in 1999 and 2001 | |
| Harnish Creek | *77°37.00'S, 163°13.00'E | soil and stream sediment | ant | Medium (n = 20, 70%) | This paper, collected in 2000 | |
| South side | NA, NA | soil | ant | M-low (n = 96, 97%) | This paper, collected in 2000, 2002 and 2003 | |
| - | 77°36.00'S, 162°15.00'E | soil | ant | Low | ( |
|
| South side near F6 stream | 77°36.40'S, 163°15.30'E | soil and lake sediment | ant | M-low (n = 12, 33%) | This paper, collected in 2002 | |
| South side near Green Creek | 77°37.36'S, 163°03.91'E | soil | ant | Medium (n = 20, 45%) | This paper, collected in 2003 | |
| South Side near F6 stream | 77°36.72'S, 163°15.18'E | soil | ant | M-low (n = 20, 35%) | This paper, collected in 2003 | |
| Green Creek | 77°37.36'S, 163°03.91'E | soil | ? | Medium | ( |
|
| Von Guerard stream | *77°37.00'S, 163°15.00'E | soil | ? | M-Low | ( |
|
| Lake Hoare | *77°38.00'S, 162°51.00'E | |||||
| North side | 77°37.49'S, 162°54.31'E | soil | ant | Low (n = 18, 78%) | This paper, collected in 1993 | |
| South side | 77°38.03'S, 162°52.75'E | soil | ant | Low (n = 9, 33%) | This paper, collected in 1993 | |
| South side | NA, NA | soil | ant | Low (n = 12, 25%) | This paper, collected in 1993 | |
| South side | 77°37.59'S, 162°52.57'E | soil | ant | M-low (n = 56, 77%) | This paper, collected in 1993, 1994, 1995, 1996, 1997 and 2001 | |
| North side | 77°38.00'S, 162°53.00'E | soil (0-2.5, 2.5-5, 5-10, 10-20 cm) | ant | M-low | ( |
|
| South side | NP, NP | soil at varying elevations | ant, gla | M-low | ( |
|
| - | NA, NA | soil polygons | ant | Low (n = 104, 17%) | This paper, collected in 1994 | |
| North side | 77°38.00'S, 162°53.00'E | soil (0-2.5, 2.5-5, 5-10, 10-20 cm) | ant | M-low | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil at varying elevations | ant | Low | ( |
|
| South side | NA, NA | soil polygons | ant | Low (n = 24, 54%) | This paper, collected in 1995 | |
| South side | 77°37.93'S, 162°53.19'E | soil | ant | M-low (n = 150, 51%) | This paper, collected in 1995, 1998 and 2002 | |
| North side | 77°37.49'S, 162°54.31'E | soil | ant | Low | ( |
|
| - | NP, NP | soil | ant | M-low | ( |
|
| South side | 77°38.00'S, 162°53.00'E | soil | ant, gla | Medium | ( |
|
| - | NP, NP | soil, sediment | ant | Low | ( |
|
| North side | NA, NA | soil | ant | Low (n = 8, 38%) | This paper, collected in 1999 | |
| South side | NA, NA | soil | ant | M-low (n = 8, 75%) | This paper, collected in 1999 | |
| South side | 77°38.00'S, 162°53.00'E | soil | ant | M-low | ( |
|
| North side | 77°37.29'S, 162°54.19'E | soil | ant | Low | ( |
|
| South side | 77°38.07'S, 162°52.59'E | soil | ant | Low (n = 12, 58%) | This paper, collected in 2001, 2002 | |
| South side | 77°38.00'S, 162°53.00'E | soil | ant | M-low | ( |
|
| - | 77°37.90'S, 162°53.20'E | soil and lake sediment | ant | M-low (n = 11, 64%) | This paper, collected in 2002 | |
| - | 77°37.00'S, 160°50.00'E | soil | ant | M-low | ( |
|
| - | NP, NP | soil | ant | M-low | ( |
|
| - | NP, NP | 0-5 cm soil (subnivian) | ant | M-low | ( |
|
| Nussbaum Riegel | 77°38.52'S, 162°46.89'E | soil | ant | Low (n = 5, 60%) | This paper, collected in 1997 | |
| Suess Glacier, 50 m away | *77°38.00'S, 162°40.00'E | soil | ant | Present | ( |
|
| Suess Pond | NP, NP | NP | ant | Present | ( |
|
| Victoria Valley | *77°23.00'S, 162°00.00'E | |||||
| Lake Vida | *77°23.29'S, 161°56.05'E | NP | ant | Present | ( |
|
| Upper | NA, NA | soil | ant | Low (n = 20, 5%) | This paper, collected in 1990 | |
| Victoria Upper Glacier | 77°17.35'S, 161°33.03'E | soil | ant | Low (n = 10, 43%) | This paper, collected in 1993 | |
| - | 77°23.00'S, 162°00.00'E | soil | ant | Low | ( |
|
| - | NA, NA | soil | ant | Present (n = 6, 50%) | This paper, collected in 2003 | |
| Wright Valley | *77°31.39'S, 161°58.70'E | |||||
| Between Lake Vanda and Lake Bull | NP, NP | dry algae around the edge of small ponds | ant | Present | ( |
|
| Dais | *77°33.00'S, 161°16.00'E | soil | ant | Low (n = 3, 67%) | This paper, collected in 2000 | |
| East of Meserve Glacier | *77°31.00'S, 162°17.00'E | algal mat | ant | Present | ( |
|
| Labyrinth | *77°33.00'S, 160°50.00'E | |||||
| West | 77°33.04'S, 160°43.15'E | soil | ant | Low (n = 9, 89%) | This paper, collected in 1993 | |
| - | 77°33.04'S, 160°43.15'E | soil | ant | Low (n = 9, 11%) | This paper, collected in 1993 | |
| - | 77°33.04'S, 160°43.15'E | soil | ant | Low | ( |
|
| West | 77°33.02'S, 160°43.09'E | soil | ant | Low | ( |
|
| - | NA, NA | soil | ant | Low (n = 12, 8%) | This paper, collected in 2003 | |
| Bull Pass | *77°28.00'S, 161°46.00'E | soil | ant | Low (n = 12, 33%) | This paper, collected in 1990 | |
| Bull Lake | *77°31.51'S, 161°42.68'E | soil | ant | Low (n = 22, 5%) | This paper, collected in 1990 | |
| - | *77°28.00'S, 161°46.00'E | soil | ant | M-Low (n = 24, 12.5%) | ( |
|
| Lake Vanda | *77°32.00'S, 161°33.00'E | |||||
| - | NP, NP | NP | ant | Present | ( |
|
| Near Lake Vanda and Péwé Lake | *77°32.00'S, 161°33.00'E | stony soil | ant | Present | ( |
|
| Met Station | NA, NA | soil | ant | M-low (n = 2, 100%) | This paper, collected in 2002 | |
| Lake Brownworth | *77°26.00'S, 162°45.00'E | NP | ant | Present | ( |
|
| - | NA, NA | soil | ant | Low (n = 5, 60%) | This paper, collected in 1997 | |
| - | 77°31.00'S, 161°50.00'E | soil | ant | M-low | ( |
|
| - | NA, NA | soil | ant | Present (n = 10, 50%) | This paper, collected in 2003 | |
| Onyx River pond | *77°32.00'S, 161°45.00'E | NP | ant | Present | ( |
|
| Linnaeus Terrace ASPAB | 77°35.83'S, 161°05.00'E | soil | ant | Low (n = 16, 6%) | This paper, collected in 1990 | |
| Koettlitz Glacier and Southern Coastal Regions | *78°15.00'S, 164°15.00'E | |||||
| Cape Chocolate(north) | *76°56.00'S, 164°35.00'E | moraine | ant | Present | ( |
|
| Strand Moraines | *77°45.04'S, 164°29.90'E | algal mat (in stream bed), sandy soil | ant | Present | ( |
|
| Marble Point | *77°26.00'S, 163°50.00'E | NP | ant | Present | ( |
|
| Northern Coastal Regions | ||||||
| Cape Adare | *71°17.00'S, 170°14.00'E | NP | ant | Present | ( |
|
| Cape Hallett | *72°19.00'S, 170°16.00'E | |||||
| Hallett Station | *72°19.00'S, 170°16.00'E | NP | ant | Present | ( |
|
| - | NA, NA | soil | ant | Low (n = 20, 20%) | This paper, collected in 2003 | |
| Cape Hallett | 72°19.29'S, 170°13.52'E | soil | Low (n = 67, 67%) | ( |
||
| Edmonson Point | *74°20.00'S, 165°08.00'E | |||||
| - | NA, NA | soil | ant | Low (n = 10, 30%) | This paper, collected in 1996 | |
| - | NA, NA | soil | ant | Present (n = 28, 7%) | This paper, collected in 1996 | |
| - | NP, NP | NP | ant, gla | Present | ( |
|
| - | NA, NA | soil | ant | Low (n = 8, 25%) | This paper, collected in 2001 | |
| Gondwana Station | 74°37.57'S, 164°11.91'E | soil | ant | Low (n = 371, 37%) | ( |
|
| Luther Peak | *72°21.88'S, 169°50.91'E | soil | ant | M-low (n = 40, 85%) | This paper, collected in 2003 | |
| Luther Vale North | 72°22.00'S, 169°53.00'E | soil | ? | M-Low | ( |
|
| Luther Vale South | 72°22.00'S, 169°53.00'E | soil | ? | M-Low | ( |
|
| Terra Nova Bay | *74°54.51'S, 164°27.19'E | |||||
| 600 km north and south of the Italian station | NP, NP | mosses, lichens, fresh-water sediments and penguin excrement (there are no details of whether Eudorylaimus occurred in all habitats or only in some) | ant | Present | ( |
|
| Barclay Glacier | NP, NP | algae in meltwater | ant | Present | ( |
|
Biogeographic distribution of Panagrolaimus davidi in Victoria Land, Antarctica. NP = not published. NA = not available. For references to “this paper”, the year collected refers to the year at the beginning of the austral summer in which samples were collected. For abundance, M-low = 21 to 200 nematodes per kg dry soil, Medium = 201 to 600 nematodes per kg dry soil, n = number of samples and % = percentage of samples in which Panagrolaimus occurred.
| Biogeographic location | Lat, Long | Habitat | Abundance | Reference | |
|---|---|---|---|---|---|
| McMurdo Dry Valleys | *77°30.00'S, 162°00.00'E | ||||
| Miers Valley | *78°06.00'S, 164°00.00'E | soil | M-low (n = 24, 29%) | This paper, collected in 1990 | |
| Southern Coastal Region | |||||
| Marble Point | *77°26.00'S, 163°50.00'E | mossy soil (Bryum antarcticum) | Present | ( |
|
| Northern Coastal Region | |||||
| Cape Bird | 77°13.00'S, 166°26.00'E | soil in penguin rookery | Medium (n = 29, 52%) | ( |
|
| Cape Crozier | 77°27.00'S, 169°11.00'E | soil in penguin rookery | M-low (n = 27, 48%) | ( |
|
| Cape Hallett | *72°19.00'S, 170°16.00'E | ||||
| - | NA, NA | soil in penguin rookery | Low (n = 2, 50%) | This paper, collected in 2002 | |
| Willet Cove | 72°19.00'S, 170°14.00'E | soil | M-High | ( |
|
| Seabee Spit | 72°18.83'S, 170°13.00'E | soil | Low | ( |
|
| Cape Hallett | 72°19.29'S, 170°13.52'E | soil | M-Low (n = 56, 56%) | ( |
|
| Cape Royds | 77°33.00'S, 166°10.00'E | soil amongst penguin rookery | M-low (n = 66, 20%) | ( |
|
| - | NA, NA | soil pits amongst penguin rookery | Med (n = 20, 70%) | This paper, collected in 2003 | |
| Edmonson Point | *74°20.00'S, 165°08.00'E | soil | Present (n = 28, 4%) | This paper, collected in 1996 | |
| Gondwana Station | 74°37.57'S, 164°11.91'E | soil | M-Low (n = 371, 34%) | ( |
|
Biogeographic distribution of Geomonhystera antarcticola in Victoria Land, Antarctica. NP = not published. NA = not available. For references to “this paper”, the year collected refers to the year at the beginning of the austral summer in which samples were collected. For abundance, Low = >0 to 20 nematodes per kg dry soil, M-low = 21 to 200 nematodes per kg dry soil, n = number of samples and % = percentage of samples in which Geomonhystera occurred.
| Biogeographic location | Lat, Long | Habitat | Abundance | Reference | |
|---|---|---|---|---|---|
| McMurdo Dry Valleys | *77°30.00'S, 162°00.00'E | ||||
| Alatna Valley | *76°52.82'S, 161°13.82'E | ||||
| Battleship Promontory | *76°54.85'S, 160°59.34'E | ||||
| - | NA, NA | soil | Low (n = 17, 47%) | This paper, collected in 1993 | |
| Southwestern Bluff | 76°55.00'S, 161°03'.00E | soil | Low (n = 14, 14%) | This paper, collected in 2001 | |
| - | NA, NA | soil | Low (n = 6, 50%) | This paper, collected in 2003 | |
| Taylor Valley | *77°38.82'S, 163°03.08'E | ||||
| Lake Bonney | *77°43.00'S, 162°25.00'E | soil | M-low (n = 2, 50%) | This paper, collected in 1998 | |
| Wright Valley | *77°31.39'S, 161°58.70'E | ||||
| 183 m east of Meserve Glacier | *77°31.00'S, 162°17.00'E | algal mat on soil | Present | ( |
|
| Between Lake Vanda and Lake Bull | NP, NP | dry algae from the edge of a small pond | Present | ( |
|
| - | soil | Low (n = 10, 20%) | This paper, collected in 2003 | ||
| Northern Coastal Region | |||||
| Edmonson Point | *74°20.00'S, 165°08.00'E | soil | Present | ( |
|
Ecology of Nematode Genera in Victoria Land.
| Genus | Co-occurs with | Nematode community complexity | Feeding | Reproduction |
|---|---|---|---|---|
| Scottnema | Eudorylaimus, Plectus, Geomonhystera, Panagrolaimus (rare, only in Dry Valleys; |
1 species- most common 2 species- often (usually Eudorylaimus antarcticus), 3 or 4 species- rare 5 species- not recorded ( |
bacteria, yeast ( |
amphimictic ( |
| Plectus | Scottnema, Eudorylaimus, Geomonhystera, Panagrolaimus | 1 species- rare 2 species- most common (usually with Eudorylaimus), 3- often 4 species- rare 5 species- not recorded |
bacteria ( |
usually unisexual (parthenogenic), males do exist but are very rare ( |
| Eudorylaimus | Scottnema, Plectus, Geomonhystera, Panagrolaimus |
1 species- not recorded 2 species- most common (usually with Scottnema or Plectus), 3- often 4 species- rare 5 species- not recorded |
Eudorylaimus are thought to feed on fungi, unicellular algae and soil invertebrates ( |
amphimictic (Eudorylaimus antarcticus) ( |
| Panagrolaimus | Scottnema (rare, only in Dry Valleys), Eudorylaimus, Plectus | 1 species- common (most common outside of Victoria Land) 2 species- rare 3- common (with Eudorylaimus and Plectus), 4 species- rare, only in Dry Valleys 5 species- not recorded ( |
bacteria ( |
amphimictic ( |
| Geomonhystera | Scottnema, Eudorylaimus, Plectus | 1 species- not recorded 2 species- often (with Eudorylaimus antarcticus) 3- most common (with Scottnema lindsayae and Eudorylaimus antarcticus) 4 species- often 5 species- not recorded (this paper) |
algae, fungi, actinobacteria ( |
amphimictic ( |
Only five genera of terrestrial nematodes are recorded from Victoria Land Antarctica: Scottnema, Plectus, Eudorylaimus, Panagrolaimus, and Geomonhystera. For some genera species delimitation remains unresolved (
Scottnema is an exclusively Antarctic genus comprised of only one species, Scottnema lindsayae (
Biogeographic distribution. Scottnema lindsayae is the dominant nematode of Victoria Land (Table 1) based on abundance and widespread distribution in numerous samples from the McMurdo Dry Valleys (
Scottnema lindsayae also occurs on two islands off the coast of Victoria Land: Ross Island (
Habitat. Scottnema lindsayae survives in a wide range of terrestrial habitats (Table 1). In Victoria Land Scottnema lindsayae occurs most commonly in dry, bare and sandy or rocky soils and has been found at 30–40 cm soil depth near south shore of Lake Hoare (
In comparison with other nematodes of Victoria Land, Scottnema lindsayae occurs most frequently and at greater abundances in soil habitats with lower moisture, higher pH, higher EC, and higher inorganic C (
Scottnema lindsayae is recorded at a range of elevations, from the McMurdo Dry Valley floors to about 600 and 1300 m above sea level (at Mt. Suess and Battleship Promentory, respectively) in Victoria Land (
Several Plectus species have been described from Antarctica: Plectus antarcticus (
Biogeographic distribution. Four Plectus species have been recorded from Victoria Land: Plectus antarcticus, Plectus frigophilus, Plectus murrayi and Plectus acuminatus. Specimens of Plectus antarcticus previously described from Victoria Land have been reinterpreted as synonymous with Plectus murrayi (and Plectus belgicae and Plectus parvus) (
In the McMurdo Dry Valleys, only Plectus murrayi and Plectus frigophilus occur, with Plectus murrayi the most abundant and widespread (Table 2). Plectus murrayi and Plectus frigophilus (
Habitat. All Plectus spp. of Victoria Land occupy similar habitats. They are present in soils and sediments (
Soil moisture is a critical factor determining the suitability of habitats for Plectus spp.
There are six recognized Eudorylaimus species endemic to continental Antarctica: Eudorylaimus antarcticus (Yeates, 1970), Eudorylaimus nudicaudatus (Heyns, 1993), Eudorylaimus shirasei (Kito, Shishida & Ohyama, 1996), Eudorylaimus glacialis (Andrássy, 1998), Eudorylaimus quintus (
Biogeographic distribution. Eudorylaimus antarcticus is widely distributed within Victoria Land (Table 3).
Outside of the Victoria Land region, Eudorylaimus antarcticus has been reported from several of the maritime islands (Signy, Alexander, King George, Anvers) (e.g.
Habitat. Eudorylaimus antarcticus in Victoria Land occurs at varying elevation and most commonly in soils and in lake sediments. The genus has also frequently been associated with algal mats, both dry and moist found in meltwater, streambeds and lakes. Eudorylaimus antarcticus has been reported less frequently in areas of moss and from soils. In contrast, outside Victoria Land (e.g. Ross Island) the occurrence of Eudorylaimus antarcticus in a moss habitat (e.g. Bryum argenteum) is common, but it does not occur in penguin rookeries (on Ross Island or in Victoria Land). In soils of the McMurdo Dry Valleys Eudorylaimus antarcticus tends to be found in soils with higher moisture, NH4-N, NO3-N, organic C, and organic C/organic N ratios, and only occurs in soils with low salinity (EC <100 mS cm-1) (
Biogeographic distribution. The Antarctic Panagrolaimus consists of two species, Panagrolaimus magnivulvatus and Panagrolaimus davidi (but see
Panagrolaimus davidi has been recorded from Ross Island (e.g.
Habitat. Penguin rookeries and moss-covered soils appear to be the most favorable habitats for Panagrolaimus davidi in Victoria Land and are consistent with the habitats where Panagrolaimus davidi has been found in other Antarctic ice-free areas (
Several nematode species originally described as Monhystera were redescribed by Andrássy in
Biogeographic distribution. Geomonhystera antarcticola are generally rare, and along with Panagrolaimus davidi are the least abundant and most patchily distributed of all nematodes in Victoria Land. Other species of Geomonhystera occur in the islands of the maritime Antarctic (Signy, Coronation, Elephant, Intercurrence and Galindez) where Geomonhystera antarcticola is one of the most common nematode species (
Sohlenius et al. recorded Monhystera from the Nunataks of Dronning Maud Land, East Antarctica (
Habitat. The habitat of Geomonhystera in Victoria Land differs from that of Geomonhystera as described by
Nematode diversity in Victoria Land is low compared to the Antarctic Peninsula, but the presence of a few cryptic species is likely (
Our knowledge of nematode biodiversity, distribution, and function in Victoria Land is based on clusters of studies from a few distinct regions, such as the McMurdo Dry Valleys, and far northern coastal Victoria Land, which are accessible from established research stations. The rest of Victoria Land (including other inland ice-free areas) has been largely inaccessible. Studies throughout the McMurdo Dry Valleys are also patchy with some valleys being studied heavily (e.g. Taylor Valley) whilst others (e.g. Barwick Valley) have barely been investigated. More undescribed nematodes may occur in these less studied regions.
Habitat suitability for each nematode species is determined primarily by variations in soil factors such as quantities and types of organic material, moisture and salinity (
We have made considerable progress in understanding the basic relationships between soil properties and the distribution of the key nematode taxa throughout Victoria Land. Suitable habitats can be defined by moisture, salinity, organic matter and nutrient content, and the interactions between these factors. Manipulations of soil moisture and field observations of environmental change during pulse warming events show that nematode community composition can respond on time scales of seasons to decades (
The nematofauna of Victoria Land are capable of long distance dispersal by wind (
We thank Rich Alward, Jackie Aislabie, Megan Balks, Jeb Barrett, Yvonne Baskin, Steve Blecker, Pella Brinkman, Dan Bumbarger, Melody Burkins, Iain Campbell, Katie Catapano, C. Page Chamberlain, Tom Cioppa, Ericha Courtright, Nicole DeCrappeo, Joel Feldman, John Freckman, Jerry Garcia, Ian Hawes, Mengchi Ho, David Hopkins, Mary Kratz, Ed Kuhn, Kevin Levine, Elizabeth Marlies, Jen Mercer, Johnson Nkem, Phil Novis, Andy Parsons, Mike Poage, Dorota Porazinska, Laura Powers, Rusty Rodriguez, Jim Schulz, Rebekka Stucker, Amy Treonis, Lewis White, Roger Worland, and David Wynn-Williams for help collecting and processing soil samples. Ethan Adams, Eric Sokol and Ian Hogg helped process the Cuticularia fermata sample, which was collected as part of the NZ TABS project (http://nztabs.ictar.aq). This work could not have been completed without the dedicated, expert helicopter support provided by personnel of the US Coast Guard, US Navy VXE-6, Petroleum Helicopters Inc., and logistic and science support by ITT, Antarctic Support Associates, and Raytheon Polar Services. Brad Herried and Paul Morin of the Polar Geospatial Center (http://www.pgc.umn.edu) helped generate Figure 1. We very much appreciate the thoughtful, constructive criticisms of three anonymous reviewers. This research was supported by National Science Foundation Grants DPP 88-18049 and DPP 89-14655, OPP 9120123, OPP 9421025, and the McMurdo Long Term Ecological Research program (OPP 9211773, OPP 9810219).