





(C) 2013 Albert Deler-Hernández. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
The Cuban fauna of the genus Berosus Leach, 1817 is reviewed based on newly collected material as well as historical and type specimens. Nine species are recognized, including three recorded from Cuba for the first time: Berosus infuscatus LeConte, 1855, Berosus interstitialis Knisch, 1924 (= Berosus stribalus Orchymont, 1946 syn. n.) and Berosus metalliceps Sharp, 1882. Only one of the nine Cuban species, Berosus chevrolati, remains endemic to Cuba, as two other species previously considered as endemic to Cuba are recorded from elsewhere: Berosus quadridens from Mexico and Central America and Berosus trilobus from the Dominican Republic. Notes on biology and Cuban distribution are provided for all nine species. Berosus quadridens Chevrolat, 1863, stat. restit. is removed from synonym with Berosus truncatipennis and considered a valid species.
Hydrophilinae, Berosini, taxonomy, new synonymy, new records, Caribbean, Neotropical region, identification key
The hydrophilid genus Berosus Leach, 1817 is the largest genus in family Hydrophilidae, containing more than 270 species distributed worldwide (
In this paper we provide a review of the Cuban fauna of Berosus containing redescriptions of the three of four species described as Cuban endemics (Berosus chevrolati, Berosus quadridens and Berosus trilobus), we synonymize the fourth supposedly endemic species Berosus stribalus with a widely distributed Caribbean Berosus interstitialis, provide identification key and illustrations of all Cuban species and notes on their distribution and bionomics based on newly collected material. Three species are newly recorded for the Cuban fauna.
This study is mainly based on the material collected during the field survey of Cuban aquatic beetles conducted between 2008 to 2012 by A. Deler-Hernández, Y. S. Megna and F. Cala-Riquelme. The survey was mainly focused on eastern Cuba, but several areas of western Cuba were also sampled. In total, the samples from 170 localities have been collected, of which only 40 sites yielded Berosus. Specimens were collected with aquatic nets and preserved in 70%-95% ethanol. Except of this material, we also used the following sources of information: i) recently collected specimens provided to us by some Cuban colleagues; ii) material deposited in the zoological collection of the Instituto de Ecología y Sistemática in La Habana, Museo de Historia Natural “Charles T. Ramsden”, Universidad de Oriente in Santiago de Cuba, National Museum in Prague and the Division of Entomology of the University of Kansas in Lawrence; and iii) literature records (
Habitus photographs were taken using Canon D-550 digital camera with attached Canon MP-E65mm f/2.8 1–5× macro lens, and subsequently adapted in Adobe Photoshop CS2. Photographs of genitalia were taken using Nikon Coolpix P6000 digital camera attached to Olympus BX41 compound microscope and subsequently combined with Helicon Focus software. Line drawings were traced from the photographs taken using a Canon PowerShot A620 camera attached to a Zeiss Stemi 2000-C stereomicroscope or with the same equipment as for taking the habitus photographs. Dissections of male genitalia and mounting techniques follow those used by
Examined material is deposited in the following collections:
BSC-E Departamento de Zoología, Centro Oriental de Ecosistemas y Biodiversidad, Santiago de Cuba, Cuba (A. Deler-Hernández);
CZACC Colección Zoológica, Instituto de Ecología y Sistemática, La Habana, Cuba (I. Fernández);
CZCTR Museo de Historia Natural “Charles Ramsden”, Facultad de Ciencias Naturales, Universidad de Oriente, Santiago de Cuba, Cuba (C. T. Ramsden historical collection) (M. Soto);
IRSNB Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium (P. Limbourg);
KSEM Division of Entomology, University of Kansas Natural History Museum, Lawrence, USA (A. Short);
NMPC National Museum, Prague, Czech Republic (M. Fikáček);
MNHN Museum National d’Histoire Naturelle, Paris, Frances (Bedel collection) (A. Mantilleri).
(asterisk indicates the species newly recorded for Cuba)
Berosus chevrolati Zaitzev, 1908
Berosus exiguus (Say, 1825)
*Berosus infuscatus LeConte, 1855
*Berosus interstitialis Knisch, 1924
= Berosus stribalus Orchymont, 1946, syn. n.
*Berosus metalliceps Sharp, 1882
Berosus peregrinus (Herbst, 1797)
Berosus quadridens Chevrolat, 1863 stat. restit.
Berosus trilobus Chevrolat, 1863
Berosus undatus (Fabricius, 1792)
Adults are mostly medium-sized, elongate, and strongly convex. Coloration of the body is brown to yellowish-brown, with or without dark spots on the pronotum and elytra. The head is strongly flexed down, eyes are protuberant, and antennae have 7 antennomeres. The elytral apex is entire or produced into one or two spines. The mesoventral process is usually laminar. The male protarsi are widened and have four tarsomeres; those of females have five tarsomeres. The middle and hind tibiae and tarsi bear a fringe of long natatory setae. Abdominal ventrite 5 has a rectangular (or less frequently semicircular) emargination posteriorly.
Among Cuban hydrophilid genera, Berosus may be easily identified by large globular eyes, scutellum longer that wide and middle and hind tibiae and tarsi with well developed fringe of long natatory setae.
http://species-id.net/wiki/Berosus_chevrolati
Figures 1a–g, 11, 12aCuba.
Holotype: female (MNHN): “aculeatus / Ch. Cuba // this must be / Chevr. unique type of aculeatus / PJS [= P. J. Spangler] 1966”.
CUBA: Santiago de Cuba: 3 exs. (dry-mounted) (NMPC): Dos Caminos, stream, 20°11'2.50"N, 75°46'17.7"W, 150 m a.s.l., 01.viii.2008, leg. A. Deler-Hernández., 1 ex. (dry-mounted) (BSC-E): El Vivero, 1.6 km E of Dos Caminos, 20°11'2.50"N, 75°46'17.7"W, 150 m a.s.l. Guaninicú river, 20–21.vi.2012, leg. Deler-Hernández & Fikáček; 1 ex. (in alcohol) (BSC-E): La Maya, Cuatro Caminos, remanso [backwater] 20°07'58"N, 75°34'01"W, 150 m a.s.l, 24.i.2008, leg. Y. S. Megna, 00019.
Published Cuban records: Pinar del Río: Quemado de Pineda (

Berosus chevrolati Zaitzev, 1908. a habitus in dorsal view b habitus in lateral view c mesoventral process in lateral view d-f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Small, widely elongate species, body length 3.6-4.6 mm. Head dark, metallic; pronotum pale, with median unpaired narrow black longitudinal spot mesally, pronotal punctation not darkened; elytra pale with irregular small dark spots in posterior half of elytral intervals. Elytral apices each without subapical tooth. Mesoventral process highly laminar, square-shaped, with large anterior and posterior teeth. Abdominal ventrite 1 with median keel throughout its length. Emargination of abdominal ventrite 5 rectangular with a median teeth. Median lobe of the aedeagus with short basal projection and rounded apex in lateral view.
Berosus chevrolati resembles Berosus trilobus (with which it may even co-occur) by the small strongly punctate body, metallic head, presence of an unpaired dark spot on the pronotum, mesoventrite with hooded anterior tooth, median keel developed throughout abdominal ventrite 1, emargination of abdominal ventrite 5 rectangular with single median tooth and the median lobe of the aedeagus with long basal lobe projecting far posteriad and enlarged apical portion in lateral view. It differs from Berosus trilobus by the narrow central dark spot on the pronotum (dark spot is large and trilobate in Berosus trilobus), elytra more evenly convex (subapical area of each elytra forms a bump in Berosus trilobus), shortbasal projection of the median lobe (long in Berosus trilobus) and, rounded apex of the median lobe in lateral view (apex is beak-shaped in lateral view in Berosus trilobus).
Habitus as in Figs 1a, b. Body length 3.4–4.6 mm. Body short and wide, moderately convex. Head black with metallic sheen, labrum black. Antennae testaceous. Maxillary palpi testaceous with palpomere 4 brown at apex. Pronotum testaceous with a central elongate metallic spot. Scutellum black with metallic sheen. Elytra testaceous with small brown spots without discrete borders. Pro-, meso- and metafemora testaceous, basal portion of metafemora sometimes slightly darker.
Head with moderately large and rounded punctures. Pronotum with punctures of the same size as on head. Scutellum with a few deeply impressed punctures slightly smaller than those on the pronotum. Elytral striae well-impressed. Interstriae with small and shallow punctures, irregular long setae on posterior half of elytra; spine-like setae absent. Elytral apices entire and rounded, of same shape in males and females. Mesoventral process highly raised, square-shaped, with hood-like anterior tooth, posterior tooth moderately large (Fig. 1c). Metaventral process wide, slightly raised, square-shaped, with large, deep glabrous rhomboid median depression; posterolateral angles raised and rounded, posteromesal projection carinate. Abdominal ventrite 1 with median carina throughout its length. Abdominal ventrite 5 with deep rectangular emargination, bearing a broad median tooth (Fig. 1g). Basal pubescence on basal 0.7 of meso- and of metafemora, the margin between pubescent and bare portions sinuate. Protarsus of male with adhesive soles on the first basal tarsomeres, first and second tarsomere distinctly thickened, third tarsomere very slightly thickened, fourth tarsomere elongate, almost as long as tarsomeres 1-3 combined. Claws moderately long, slender, arcuate.
Male genitalia (Figs 1d–f): Phallobase ca. 0.6× total length of aedeagus. Parameres in lateral view wide basally, apically projecting into rounded apex slightly bent ventrad, bearing a row of subapical setae ventrally. Median lobe C-shaped in lateral view; basal projection short, directing apicad; apex wide and rounded in lateral view.
Currently only known from Cuba.
We collected Berosus chevrolati along the margins of lowland streams and in isolated pools along these streams, in both cases having clear to turbid water and abundant organic matter (Fig. 11a). This species is found at low altitudes (ranging from sea level to ca. 160 m a.s.l.) situated in the Central Valley (Valle Central). Berosus chevrolati is frequently associated with Berosus trilobus in those habitats.
http://species-id.net/wiki/Berosus_exiguus
Figures 2a–g, 11USA: Virginia, Chincoteague Island.
CUBA: Isla de la Juventud: 1 ex. (in alcohol) (BSC-E): Punta del Este, Laguna Cayamás, 21°33'43"N, 82°33'18"W, 3 m a.s.l., 23.v.2006, leg. Y. S. Megna, 00165. Camagüey: 2 exs. (in alcohol) (BSC-E): Nuevitas, Cayo Sabinal, Laguna permanente [permanent pool], 21°38'6.1"N, 77°10'8.2"W, 5 m a.s.l., 06.v.2010, leg. O. Bello, 00153; 2 exs. (dry-mounted) (NMPC): Cayo Sabinal, permanent lagoon, 21°38'6.1"N, 77°10'8.2"W, 5 m a.s.l., 06.v.2010, leg. Y. Torres. Santiago de Cuba: 1 ex. (in alcohol) (BSC-E): San Miguel de Parada, Laguna temporal [temporal pool], 20°11'2.50"N, 75°46'17.7"W, 1 m a.s.l., 29.v.2009, leg. A. Deler-Hernández, 00136; 4 exs. (in alcohol) (BSC-E): San Miguel de Parada, Laguna temporal [temporal pool], 20°11'2.50"N, 75°46'17.7"W, 1 m a.s.l., 05.ix.2009, leg. A. Deler-Hernández, 00151; 2 exs. (dry-mounted) (NMPC): San Miguel de Parada, temporal lagoon, 20°11'2.50"N, 75°46'17.7"W, 1 m a.s.l, 05.ix.2009, leg. A. Deler-Hernández.
Published Cuban records: Pinar del Rio: unspecified locality (

Berosus exiguus (Say, 1825). a habitus in dorsal view b habitus in lateral view c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Habitus as in Figs 2a, b. Body length 3.0–3.7 mm. Head testaceuos, pronotum testaceous without median darker spots, punctation not darkened, elytra testaceous with irregularly arranged ill-defined slightly darker spots. Elytral apices entire and rounded in both sexes. Mesoventral process highly laminar, triangular in shape, anterior tooth large projecting posteriad (Fig. 2c). Abdominal ventrite 1 with median keel developed on basal half only. Emargination of abdominal ventrite 5 rectangular, without teeth (Fig. 2g) (in non-Cuban specimens, a very small medial tooth is present:
Eastern USA (from New York to Florida, westwards reaching to Illinois, Indiana, Mississippi and Oklahoma), Bahamas (
Berosus exiguus is mainly restricted to brackish waters in coastal regions. Cuban specimens have been collected in temporary brackish pools with clear water, abundant organic detritus on the bottom and associated aquatic riparian vegetation.
http://species-id.net/wiki/Berosus_infuscatus
Figures 3a–g, 11USA: “middle and southern States, e.g. New Orleans”.
CUBA: Isla de la Juventud: 1 ex. (in alcohol) (BSC-E): Punta del Este, Laguna temporal [temporal pool], 21°33'43"N, 82°33'18"W, 1 m a.s.l., 21.v.2006, leg. Y. S. Megna, 00160. Pinar del Río: 1 ex. (in alcohol) (BSC-E): Guanahacabibes, 21°54'26"N, 84°39'14"W, 3 m a.s.l., 20.iii.2003, leg. Y. S. Megna and O. Bello, 00176. Camagüey: 2 exs. (dry-mounted) (NMPC): Cayo Sabinal, lagoon, 21°38'6.1"N, 77°10'8.2"W, 5 m a.s.l., 06.v.2010, leg. Y. T. Cambas. Las Tunas: 1 ex. (in alcohol) (BSC-E): Palancón, 21°00'N, 76°54'W, 100 m a.s.l., 04.viii.2004, leg. Y. S. Megna, 00017. Granma: 1 ex. (in alcohol) (BSC-E): Cauto Cristo, Laguna permanente-I [permanent pool-I], 20°33'33.1"N, 76°28'44"W, 44 m a.s.l., 04.i.2005, leg. L. Chávez, 00175. Guantánamo: 1 ex. (dry-mounted) (CZACC): [no locality and date] leg. C. T. Ramsdem.

Berosus infuscatus LeConte, 1855. a habitus in dorsal view b habitus in lateral view c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Habitus as in Figs 3a, b. Body length 5.5–6.0 mm. Head metallic black with paler anterior margin of clypeus; pronotum pale, with a pair of closely associated dark narrow longitudinal spots mesally, elytra brownish with indistinct irregularly arranged slightly darker spots. Head and pronotum with very distinct mesh-like microsculpture on interstices. Elytral apices entire and rounded in both sexes. Mesoventral process laminar, anterior tooth large, projecting posteriad (Fig. 3c). Abdominal ventrite 1 with median keel developed only between metacoxae. Emargination of abdominal ventrite 5 rectangular, with two sharp medial teeth (Fig. 3g). Aedeagus (Figs 3d–f) with median lobe slightly shorter than parameres, arched in lateral view. Parameres sinuate on lateral margin subapically.
USA (Alabama, Arkansas, Florida, Georgia, Illinois, Indiana, Louisiana, Mississippi, Missouri, North Carolina, Texas, Wisconsin), Mexico (
Cuban specimens of Berosus infuscatushave been collected among submerged aquatic vegetation in sun-exposed brackish permanent and temporary lagoons in coastal zones with turbid water and muddy/stony bottoms.
http://species-id.net/wiki/Berosus_interstitialis
Figures 4a–g, 11Guadeloupe, Grande Terre and Trois-Rivières.
Berosus tessellatus:Not examined. Type specimens were not found on our request for loan in MNHN. Instead, we examined the specimens identified as Berosus tessellatus and Berosus interstitialis deposited in coll. d’Orchymont and coll. Knisch in IRSNB (see below).
Berosus stribalus: Holotype: male (IRSNB): “[male sign] / Cuba 10. K / S. of Pinar Rio / Sep. 12-23 ’13 // A. d’Orchymont det. / Berosus (s.str.) / stribalus m. // Type”. Paratype: 1 spec. (IRSNB): “St. / Domin- / go // Berosus / striatus / Say // coll. Orch. // A. d’Orchymont det. / Berosus (s.str.) / stribalus m. // Para- / type”.
CUBA: Isla de la Juventud: 4 exs. (dry-mounted) (CZACC): vii.1960 [no locality and collector indicated]; 8 exs. (in alcohol) (BSC-E): Punta del Este, laguna temporal [temporal pool], 21°33'43"N, 82°33'18"W, 1 m a.s.l., 21.v.2006, leg. Y. S. Megna, 00180; 8 exs. (in alcohol) (BSC-E): Laguna Cayamás, 21°33'43"N, 82°33'18"W, 3 m a.s.l., 23.v.2006, leg. Y. S. Megna, 00178. Pinar del Río: 57 exs. (dry-mounted) (CZACC): Lomas de Soroa, v/vi.1963, [no collector indicated]; 1 ex. (in alcohol) (BSC-E): Viñales, arroyo [stream], 22°33'36.35"N, 83°49'59"W, 170 m a.s.l., 18.iv.2012, leg. A. Deler-Hernández, 00146. Artemisa:8 exs. (dry-mounted) (CZACC): Laguna Ariguanabo, vi.1963, [no collector indicated]. Mayabeque:1 ex. (dry-mounted) (CZACC): Jibacoa, littoral on north coast, v.1962, [no collector indicated]. Matanzas:1 ex. (dry-mounted) (CZACC): Playa Larga, iv.1965, [no collector indicated]; 1 ex. (dry-mounted) (CZACC): Bacunayagua, vi.1940 [no collector indicated]. Camagüey:1 ex. (in alcohol) (BSC-E): Sierra de Cubitas, Río El Roble, 21°32'53.23"N, 77°46'42.31"W, 55 m a.s.l., 14.iv.2012, leg. A. Deler-Hernández, 00148. Las Tunas:1 ex. (in alcohol) (BSC-E): Amancio, Comunales, laguna permanente [permanent pool], 20°49'59"N, 77°32'32"W, 34 m a.s.l., 04.x.2008, leg. Y. S. Megna, 00179; 1 ex. (in alcohol) (BSC-E): La Fé, laguna temporal [temporal pool], 20°49'17.7"N, 77°34'40.8"W, 50 m a.s.l., 18.xii.2008, leg. Y. S. Megna, 00147. Granma: 2 exs. (in alcohol) (BSC-E): Cauto Cristo, laguna permanente [permanent pool], 20°33'33.1"N, 76°28'44"W, 44 m a.s.l., 04.i.2005, leg. L. Chávez, 00150; 1 ex. (dry-mounted) (NMPC): Cauto Cristo, permanent lagoon 20°33'33.1"N, 76°28'44"W, 44 m a.s.l., 04.i.2005, leg. L. Chávez. Santiago de Cuba:1 ex. (in alcohol) (BSC-E): Guamá, La Mula, laguna permanente [permanent pool], 19°58'33.6"N, 76°46'4.8"W, 4 m a.s.l., 20.vi.2008, leg. A. Deler-Hernández, 00018. Guantánamo:1 ex. (dry-mounted) (CZCTR): Guantánamo, San Carlos [at light], 20°26'22"N, 74°42'31"W, 160 m a.s.l., 18.vii.1915, leg. C. T. Ramsden; 1 ex. (dry-mounted) (CZCTR): San Carlos [at light], 20°26'22"N, 74°42'31"W, 160 m a.s.l., 24.viii.1917, leg. C. T. Ramsden; 1 ex. (in alcohol) (BSC-E): San Antonio del Sur, Macambo, Río Macambo, 20°03'26.9"N, 74°44'15"W, 4 m a.s.l., 24.x.2008, leg. A. Deler-Hernández; 1 ex. (in alcohol) (BSC-E): Baracoa, Nibujón, laguna temporal [temporal pool], 20°30'8.6"N, 74°38'88"W, 8 m a.s.l., 03.ii.2010, leg. A. Deler-Hernández, 00149. 1 ex. (dry-mounted) (IRSNB): [without precise locality]:“Cuba / Gundlach // 1541 / 977 // Kniž det. / interstitialis”. GUADELOUPE: 1 ex. (dry-mounted) (IRSNB): “Guadeloupe / coll. A. d’Orchymont // Berosus s.str. / tessellatus / Fleut. & Salle // A. d’Orchymont det.” [based on attached note, d’Orchymont compared this specimen with one of the types of Berosus tessellatus from the collection of Fleutiaux which is currently lost and not available for reexamination; the specimen is a male, but has the abdomen destroyed by a dermestid larva]; 1 ex. (dry-mounted) (IRSNB): Trois Riviéres, leg. Dufau. PUERTO RICO: 1 ex. (dry-mounted) (IRSNB): [without detailed locality data], leg. Moritz. VIRGIN ISLANDS: 2 exs. (dry-mounted) (IRSNB): Saint Thomas, leg. C. Felsche.
Published Cuban records: Pinar del Río: Entronque de Manuel Sanguili (

Berosus interstitialis Knisch, 1924. a-b habitus of the holotype of Berosus stribalus Orchymont, 1946 (a dorsal view b lateral view) c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Habitus as in Figs 4a, b. Body length 5.0–5.3 mm. Head uniformly dark, metallic green; pronotum pale with a pair of closely aggregated longitudinal narrow dark spots mesally; elytra pale with darkened punctation and with dark spots in anterior and posterior third of intervals 1 and 2, in humeral area and at midlength of intervals 7-9, plus with variable number of spots on remaining intervals. Elytral apices entire in both sexes. Mesoventral process laminar, with small anterior tooth projecting ventrad, nearly straight middle portion and rounded posterior part (Fig. 4c). Abdominal ventrite 1 with median keel developed only between metacoxae. Emargination of ventrite 5 deep, subrectangular, with two slender medial teeth (Fig. 4g), not showing sexual dimorphism. Aedeagus (Figs 4d–f) strongly compressed from sides; parameres ca. 2× as long as phallobase, wide throughout in lateral view except for tooth-like apex; bases of the parameres in dorsal view with characteristic basal teeth.
The synonymy of Berosus stribalus with Berosus interstitialis was first proposed in an unpublished thesis by
Cuban specimens were collected mainly in standing waters as well as in isolated pools along streams and rivers in the lowlands. The localities are usually exposed to sun and have turbid water, muddy bottom, submerged vegetation and are rich in organic matter.
Widely distributed Caribbean species, so far recorded from the Bahamas, Haiti Guadeloupe, Virgin Islands Puerto Rico and Cuba (
http://species-id.net/wiki/Berosus_metalliceps
Figures 5a–g, 11México: Tres Marías Island.
CUBA: Camagüey:1 ex. (dry-mounted) (NMPC): Cayo Sabinal, permanent lagoon, 21°38'6.1"N, 77°10'8.2"W, 5 m a.s.l., 06.v.2010, leg. Y. Torres.

Berosus metalliceps Sharp, 1882. a–b habitus of the only known Cuban specimen (a dorsal view b lateral view) c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Habitus as in Figs 5a, b. Body length 4.5 mm. Head metallic black, pronotum pale without dark spots, elytra pale with dark stripes on elytral series and slightly darker spot in posterior third of interval 1. Elytral apices entire and rounded. Mesoventral process laminar, triangular in shape, anterior tooth projecting posteriad (Fig. 5c). Abdominal ventrite 1 with median keel developed only between metacoxae. Emargination of abdominal ventrite 5 rectangular, without tooth median (Fig. 5g). Aedeagus (Figs 5d–f) with median lobe much longer than parameres, with enlarged spatula–shaped apex in ventral view, sinuate on dorsal face in lateral view. Parameres simple, rounded apically, phallobase ca. 0.3× total length of aedeagus.
USA (California), Mexico, Bahamas (
The Cuban specimen was collected in the highly exposed brackish permanent lagoon with muddy bottom.
http://species-id.net/wiki/Berosus_peregrinus
Figures 6a–g“North America”.
CUBA: no material examined. USA: Texas: 1 ex. (dry-mounted) (NMPC); 1 ex. (in alcohol) (BSC-E): Maverick Co., Rt. 277 at Tequesquite Creek, large creek [AS-03-011], 31.viii.2003, leg. A. E. Z. Short.
Published Cuban records: Cuba: without specified locality (

Berosus peregrinus (Herbst, 1797), specimen from USA, Texas. a habitus in dorsal view b habitus in lateral view c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Habitus as in Figs 6a, b. Body length 4.1–5.2 mm. Head metallic black, pronotum pale with two small submedian dark spots anteriorly, elytra pale with rather sharply defined dark spots on intervals 1-2 and in humeral area. Elytral apices entire and rounded. Mesoventral process laminar, triangular in shape, anterior tooth large, projecting posteriad (Fig. 6c). Abdominal ventrite 1 with median keel developed only between metacoxae. Emargination of abdominal ventrite 5 rectangular with a single median broad and short tooth (Fig. 6g). Aedeagus (Figs 6d–f) with median lobe slender, pointed at apex, parameres shorter than median lobe, very wide in lateral view, narrowing into sharply pointed apex bearing tuft of setae apically. Phallobase long, ca. 0.6× total length of aedeagus.
Canada (Nova Scotia, Ontario, Quebec), USA (from New York and Pennsylvania to Florida, Louisiana, Mississippi and Texas, westward at least to Arizona, Illinois, Indiana and Wisconsin) (
http://species-id.net/wiki/Berosus_quadridens
Figures 7a–g, 8e–h, 11Cuba.
Not examined.
Additional material examined CUBA: Pinar del Río: 1 ex. (dry-mounted) (IRSNB): S of Pinar del Rio, 12/23.ix.1913. Isla de la Juventud: 7 exs. (in alcohol) (BSC-E): Punta del Este, Laguna temporal [temporal pool], 21°33'43"N, 82°33'18"W, 1 m a.s.l., 21.v.2006, leg. Y. S. Megna, 00142. Granma: 3 exs. (in alcohol) (BSC-E): Cauto Cristo, Laguna permanente-I [permanent pool-I], 20°33'33.1"N, 76°28'44"W, 44 m a.s.l., 04.i.2005, leg. L. Chávez, 00087; 1 ex. (in alcohol) (BSC-E): Cauto Cristo, Laguna permanente-I [permanent pool-I], 20°33'33.1"N, 76°28'44"W, 44 m a.s.l., 13.vi.2004, leg. L. Chávez, 00174. Santiago de Cuba: 1 ex. (in alcohol) (BSC-E): Palma Soriano, Monte Barranca, 20°20'13.5"N, 76°1'11.6"W, 203 m a.s.l., 05.xii.2007, leg. A. Deler-Hernández and B. Téllez, 00052. MEXICO: Sinaloa:1 ex. (dry-mounted) (IRSNB): Los Mochis Station, x.1921 leg. R. V. van Zwaluwenburg. Veracruz:2 exs. (dry-mounted) (IRSNB): without more detailed locality, leg. Höge. GUATEMALA: 4 exs. (dry-mounted) (IRSNB): Paso Antonio, 400 ft., leg. Champion. NICARAGUA: Chinandega: 1 ex. (dry-mounted) (IRSNB): Posoltega, 06.v.1984, UV light, leg. Algodon. COSTA RICA: Guanacaste:10 exs. (dry-mounted) (KSEM, NMPC): 11.5 km W of Cañas, 15 m a.s.l., HG light by ditch/field [AS-04-026], leg. A. E. Z. Short & D. J. Lebbin; 1 ex. (dry-mounted) (NMPC): Highway 1, 13.1 km SW of Liberia, roadside ditch/pools, 16.vi.2003, leg. A. E. Z. Short.
Published Cuban records: Cuba: Isla de la Juventud: Laguna Base Julio Antonio Mella (

Berosus quadridens Chevrolat, 1863. a habitus in dorsal view b habitus in lateral view c apex offemale elytron d apex of male elytron e mesoventral process in lateral view f–g abdominal ventrite 5 (f female g male).
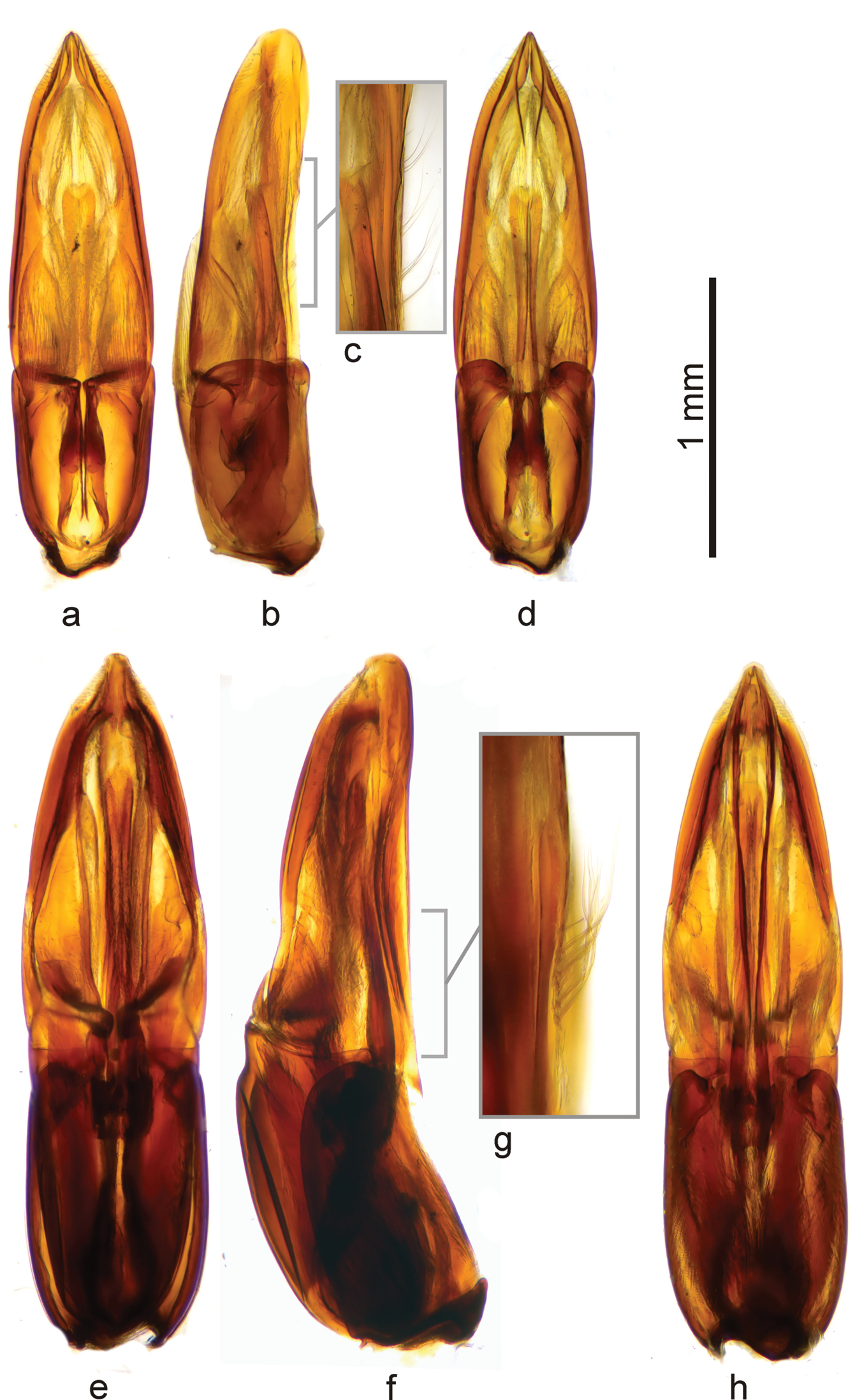
Comparison of the aedeagus of Berosus truncatipennis Castelnau, 1840 (a–d) and Berosus quadridens Chevrolat, 1863 (e–h). a, e dorsal view b, f lateral view c, g detail of setae of parameres d, h ventral view. Both aedeagi shown to scale.
Large elongate species, body length 6.2–6.7 mm. Head testaceous with darker central part of clypeus and frons; pronotum pale, with a pair of vaguely defined narrow black longitudinal spots mesally, pronotal punctation darkened; elytra pale with dark elytral striae, interval punctation and variable number of larger dark spots on elytral intervals. Elytral apices each with a large subapical tooth, sutural angle sexually dimorphic, rounded in males, sharply pointed in females. Mesoventral process highly laminar, subtriangular in shape, anterior tooth weakly developed. Abdominal ventrite 1 with median keel developed only between metacoxae. Emargination of abdominal ventrite 5 deeply and narrowly excised in males, shallowly semicircular in females. Aedeagus large, with joint parameres pointed apically, with subbasal tuft of setae on dorsal surface, ventral membranous lobes minute, median lobe slender and long.
Berosus quadridens is easily distinguishable from Berosus truncatipennis by the relatively larger and more sclerotized aedeagus having stouter and relatively longer phallobase, by ventral face of parameres bearing subbasal tuft of setae (Fig. 8g) (whereas bearing a series of setae (Fig. 8c) in Berosus truncatipennis), by relatively longer and narrower median lobe and minute membranous dorsal projections of the parameres (Figs 8e, f, h) (in contrast to moderately large ones present (Figs 8a, b, d) in Berosus truncatipennis). The aedeagus of Berosus quadridens may resemble that of Berosus megaphallus by its large size and presence of subbasal tuft of setae on ventral face of the paramere, but both species distinctly differ by the size and proportions of the phallobase (ca. half as long as the whole aedeagus and very robust in Berosus megaphallus; ca. third as long as the whole aedeadus and less robust in Berosus quadridens) and by the proportions of the ventral membranous lobe of the paramere (minute in Berosus quadridens, nearly as long as paramere in Berosus megaphallus). In general, the aedeagus of Berosus quadridens looks like an enlarged aedeagus of Berosus truncatipennis on the first view, whereas that of Berosus megaphallus clearly differs from both Berosus truncatipennis and Berosus quadridens by the general proportions of its parts. We failed to find any realiable external differences between Berosus truncatipennis and Berosus quadridens;
Habitus as in Figs 7a, b. Body length 6.2–6.7 mm. Head, labrum and antennae testaceous. Maxillary palpi testaceous with palpomere 4 dark at apex. Pronotum testaceous with two metallic black central spots developed throughout pronotum. Scutellum testaceous. Elytra testaceous with oblique series of dark brown spots in posterior half and laterally, interval punctuation and elytra striae darkened. Pro- and mesofemora testaceous; metafemora with pubescent portion brown, glabrous portion testaceous.
Head and pronotum with punctures moderately fine and rounded. Elytral striae narrow well impressed. Interstriae fine and flat, bearing spine-like setae on posterior half of elytra. Scutellum with few impressed punctures. Elytral apices bidentate, each bearing a projection on sutural angle and subapically; shape sexually dimorphic, with sutural angle rounded in males (Fig. 7d), sharply pointed in females (Figs 7c). Mesoventral process highly laminar, triangular in shape, anterior tooth barely visible, followed by a convex and smooth ridge (Fig. 7e). Metaventral process raised, triangular in shape, with elongate and deep glabrous median depression; posterolateral angles produced into triangular laminae, posterior projection pointed. Abdominal ventrite 1 with median carina only between metacoxae and with large, deep, rounded lateral depressions. Abdominal ventrite 5 with a deep rounded emargination without tooth in males (Fig. 7g), in females with semicircular apical notch (Fig. 7f). Basal pubescence of meso- and metafemora covering basal two thirds of femoral length, borderline between pubescent and glabrous portion sinuate on meso- femur, straight on metafemur. Protarsus of male with adhesive soles on the two basal tarsomeres, protarsomeres 1-2 thickened, tarsomere 1 longer than tarsomere 2, tarsomere 3 elongate; tarsomere 4 elongate, as long as tarsomeres 1-3 combined. Claws long, slender and curved.
Male genitalia (Figs 8e–h): Phallobase robust, ca. 0.4× as long as whole aedeagus, slightly widening basad in lateral view. Parameres joint mesally, together forming a dish-like structure surrounding median lobe; apical portion rounded in lateral view, pointed in ventral view; ventral portion of each paramere with minute membranous lobe; dorsal face of each paramere with a tuft of setae situated subbasally. Median lobe stick-shaped, reaching to apical 0.75 of paremeres.
Described from Cuba, Berosus quadridens was considered endemic to the island, whereas the continental form was supposed to represent the widely distributed South American species Berosus truncatipennis (e.g.,
We were not able to examine the unique type of Berosus quadridens from “Cuba”, as it was not found in MNHN after our loan request in 2012. A single species of Cuban Berosus matching the original description by
By confirming the separate species status of Berosus quadridens, the originally widely understood Berosus truncatipennis is shown to consist of three species: the widely distributed South American Berosus truncatipennis, Berosus quadridens confined to the Caribbean and Central America, and Berosus megaphallus known so far from Venezuela and Guyana. In the material from IRSNB we examined for this study, we have found few specimens from Bolivia (Río Beni) and southern Peru (Ica) which male genitalia are extremely similar to those of Berosus quadridens by their large size, strong sclerotization and relatively longer phallobase; however, they seem to differ from Berosus quadridens by the presence of the series of setae on the paramere (as in Berosus truncatipennis) and the dorsal membranous lobe of the paramere being ca. as long as in Berosus megaphallus (examined only in the Bolivian specimen, indistict in dissected Peruan ones). We suppose that these specimens may represent yet another undescribed species of the formerly broadly understood Berosus truncatipennis.
The Cuban specimens examined in the present work were collected in highly exposed freshwater pools with turbid water, muddy bottom and without cover vegetation.
Based on the specimens examined for this study, we may confirm the occurrence of Berosus quadridens for Mexico, Guatemala, Nicaragua, Costa Rica and Cuba.
http://species-id.net/wiki/Berosus_trilobus
Figures 9a–g, 11, 12a–bCuba.
Holotype: female (MNHN): “Berosus / trifidus / Chv. Cuba / … [illegible] // von / G. Hemiosus / Sharp [= of the genus Hemiosus Sharp] // this must be / Chev. unique / type of trilobus / 1966 / PJS [= P. J. Spangler]”.
CUBA: Sancti Spíritus:50 exs. (in alcohol) (BSC-E): Topes de Collantes, El Nueve, Río Caburny, 21°55'50"N, 80°00'59"W, 539 m a.s.l., 29.vi.2010, leg. A. Deler-Hernández. Camagüey: 19 exs. (in alcohol) (BSC-E): Sierra de Cubitas, Río El Roble, 21°32'53.23"N, 77°46'42.31"W, 55 m a.s.l., 14.iv.2012, leg. A. Deler-Hernández, 00144. Holguín: 6 exs. (in alcohol) (BSC-E): Jardín Botánico, Arroyo [stream], 20°51'46.8"N, 76°13'22.8"W, 84 m a.s.l., 07.xii.2008, leg. A. Deler-Hernández, 00074. Granma: 7 exs. (in alcohol) (BSC-E): Parque Nacional Turquino, La Platica, 20°00'33.80"N, 76°53'38.47"W, 800 m a.s.l., 29.iii.2012, leg. A. Deler-Hernández, 00143; 12 exs. (dry-mounted) (NMPC): Turquino NP, around La Platica, 20°0.7'N, 76°53.4'W, 880 m a.s.l. [MF24], 25-26.vi.2012 leg. A. Deler-Hernández and M. Fikáček. Santiago de Cuba: 6 exs. (in alcohol) (BSC-E): Campo Rico-II, Río Indio, 19°59'54.5"N, 75°32'4.6"W, 150 m a.s.l., 15.ix.2003, leg. A. Deler-Hernández and F. Cala-Riquelme, 00046; 4 exs. (in alcohol) (BSC-E): Gran Piedra, El Olimpo, Arroyo [stream], 20°00'33"N, 75°40'13"W, 820 m a.s.l., 04.viii.2005, leg. A. Deler-Hernández, 00016; 1 ex. (in alcohol) (BSC-E): II Palmas, La Cubana, Laguna temporal-II [temporal pool-II], 20°3'15.48"N, 76°8'3.12"W, 320 m a.s.l., 02.xii.2005, leg. Y. S. Megna, 00086; 30 exs. (in alcohol) (BSC-E): Palma Soriano, Arroyo [stream], 20°06'05"N, 75°58'44"W, 130 m a.s.l., 16.ii.2005, leg. K. Blanco, 00047; 5 exs. (in alcohol) (BSC-E): Guamá, La Mula, Río Turquino, 19°56'57"N, 76°45'36"W, 8 m a.s.l., 21.vi.2005, leg. Y. S. Megna, 00085; 6 exs. (in alcohol) (BSC-E): Guamá, Los Morones, Río Turquino, 19°58'33.6"N, 76°46'4.8"W, 200 m a.s.l., 18.vi.2008, leg. A. Deler-Hernández, 00006; 2 exs. (in alcohol) (BSC-E): San Luis, Dos Caminos, El Vivero, Laguna permanente [permanent pool], 20°11'2.50"N, 75°46'17.7"W, 150 m a.s.l., 01.viii.2008, leg. A. Deler-Hernández, 00028; 3 exs. (in alcohol) (BSC-E): San Luis, Dos Caminos, El Vivero, Río Guaninicú, 20°11'2.50"N, 75°46'17.7"W, 150 m a.s.l., 01.viii.2008, leg. A. Deler-Hernández, 00029; 31 exs. (dry-mounted) (NMPC): El Vivero, 1.6 km E of Dos Caminos, 20°10.8'N, 75°46.4'W, ca. 150 m a.s.l. [MF18], 20–21.vi.2012, leg. A. Deler-Hernández and M. Fikáček; 5 exs. (in alcohol) (BSC-E): Loma del Gato, Chan-Chan, Arroyo [stream], 19°58'27.4"N, 75°53'22.2"W, 353 m a.s.l., 27.vi.2009, leg. A. Deler-Hernández, 00118; 3 exs. (in alcohol) (BSC-E): La Redonda, Río Sevilla, 20°00'54.3"N, 75°45'45.6"W, 15 m a.s.l., 17.v.2009, leg. A. Deler-Hernández, 00154. Guantánamo: 36 exs. (in alcohol) (BSC-E) Imías, Yacabo Abajo, Río Yacabo Abajo, 20°06'05"N, 74°69'00"W, 5 m a.s.l., 24.x.2008, leg. A. Deler-Hernández and S. Muñiz, 00091; 20 exs. (in alcohol) (BSC-E): San Antonio del Sur, Macambo, Río Macambo, 20°03'26.9"N, 74°44'15.82"W, 4 m a.s.l., 25.x.2008, leg. A. Deler-Hernández and S. Muñiz, 00055; 16 exs. (in alcohol) (BSC-E): Baracoa-Maisí, Río Yumurí, 20°17'47.76"N, 74°17'39.5"W, 5 m a.s.l., 27.i.2010, leg. A. Deler-Hernández and R. Correa, 00152; 96 exs. (in alcohol) (BSC-E): Baracoa, Yunque, Río Duaba, 20°19'54.40"N, 74°34'9.08"W, 70 m a.s.l., 31.i.2010, leg. A. Deler-Hernández, 00171; 27 exs. (dry-mounted) (NMPC): El Yunque, 2.5-3.3 km SW of campismo popular, 20°19.4'N, 74°34.2'W, ca. 80-100 m a.s.l., 10.vi.2012 [MF02], leg. A. Deler-Hernández and M. Fikáček; 61 exs. (dry-mounted) (NMPC, KSEM): El Yunque, "La Cascada”, ca. 2.1 km SW of campismo, 20°19.9'N, 74°34'W, ca. 60 m a.s.l. [MF07], 12-13.vi.2012, leg. F. Cala-Riquelme, A. Deler-Hernández and M. Fikáček; 2 exs. (dry-mounted) (NMPC): El Yunque, 3.2 km SW of campismo, right tributary of Duaba river, 20°19'N, 74°34'W, ca. 150 m a.s.l. [MF09], 13.vi.2012; leg. A. Deler-Hernández and M. Fikáček; 14 exs. (dry-mounted) (NMPC): El Yunque, in/around campismo popular, 20°20.4'N, 74°32.9'W, ca. 40 m a.s.l. [MF05], 10-13.vi.2012, leg. M. Fikáček; 20 exs. (dry-mounted) (NMPC): PN Alejandro de Humboldt, ca. 1.7 km NW of Santa María, 20°32'N, 74°43'W, ca. 50 m a.s.l. [MF13], 16.vi.2012, leg. A. Deler-Hernández and M. Fikáček; 18 exs. (in alcohol) (BSC-E): Baracoa, Jamal, 20°17'13.9"N, 74°25'33.6"W, 40 m a.s.l., 09.ii.2010, leg. R. Correa, 00169; 1 ex. (in alcohol) (BSC-E): Baracoa, Cabacú, Laguna permanente [permanent pool], 20°19'14"N, 74°28'58"W, 10 m a.s.l., 04.iii.2010, leg. R. Correa, 00170; 6 exs. (in alcohol) (BSC-E) 6 exs. (dry-mounted) (NMPC): Baracoa, Cabacú, Laguna permanente [permanent pool], 20°19'14"N, 74°28'58"W, 10 m a.s.l., 16.iii.2010, leg. R. Correa , 00164; 1 ex. (in alcohol) (BSC-E): La Marsella, Río Guaso, 20°26'22"N, 74°42'31"W, 60 m a.s.l., 26.i.2004, leg. Y. S. Megna , 00173; 3 exs. (in alcohol) (BSC-E): Baracoa, Loma de los Guineos, Arroyo [stream], 20°19'38.38"N, 74°35'35.37"W, 530 m a.s.l., 07.iv.2012; leg. A. Deler-Hernández, 00177. Without precise locality:2 exs. (dry-mounted) (NMPC): “O. Koechin / Cuba // Collectio / Dr. Jureček / H. Jurečková”; 1 ex. (dry-mounted) (MNHN): “1542 / 1798”. DOMINICAN REPUBLIC: 25 exs. (dry-mounted) (KSEM, NMPC): near Hato Mayor, creek off Ruta 103, 02.xi.2000, leg. A. E. Z. Short.
Published Cuban records: Cuba: without specified locality(Gundlach, 1891). Pinar del Río: Quemado de Pineda (
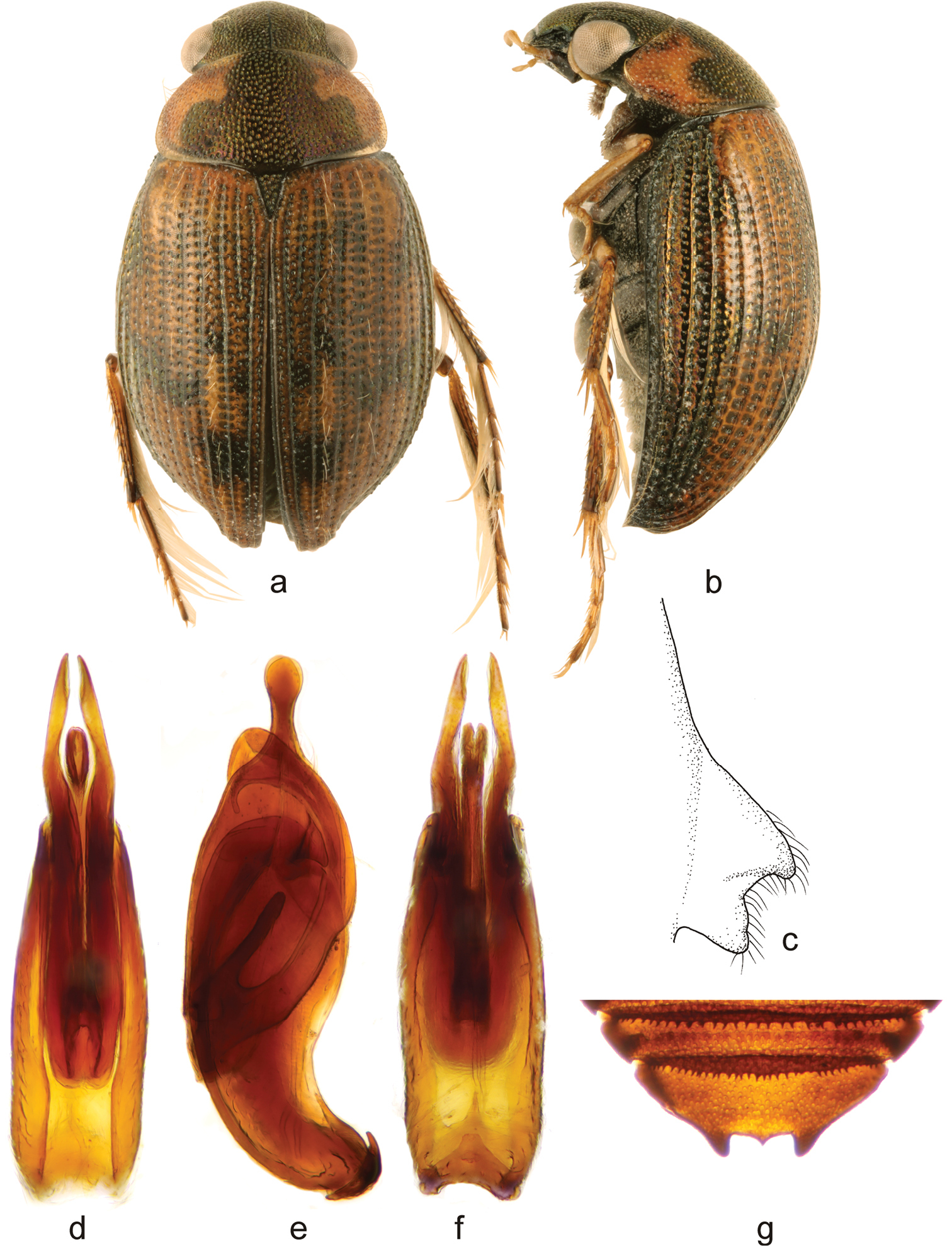
Berosus trilobus Chevrolat, 1863. a habitus in dorsal view b habitus in lateral view c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g abdominal ventrite 5.
Small widely elongate species, body length 3.2–3.7 mm. Head dark, metallic; pronotum pale laterally, with large trilobite central dark spot, pronotal punctation not darkened laterally; elytra pale with dark intervals 8-10 and large transverse dark spots on posterior half of elyttral intervals 1-7. Elytral apices without subapical tooth, bumpy along suture subapically. Mesoventral process highly laminar, rectangular with large anterior and posterior teeth. Abdominal ventrite 1 with median keel throughout its length. Emargination of abdominal ventrite 5 rectangular with a median tooth. Median lobe of the aedeagus with long basal projection and beak-like apex in lateral view.
Differential diagnosis. For diagnostic characters and difference from Berosus chevrolati, see the latter species.
Habitus as in Figs 9a, b. Body length 3.2–3.7 mm. Body short and wide, moderately convex in lateral view. Labrum black, dorsum of head melanic with strong metallic luster. Antennae testaceous. Maxillary palpi testaceous with palpomere 4 dark at apex. Pronotum testaceous with unpaired metallic black spot, the spot expanding laterad posteriorly, and hence trilobite in general shape. Elytra testaceous with small ill-defined dark brown spots on disc and, a broad metallic dark area throughout lateral portion. Pro-, meso- and metafemora with pubescent portion dark brown, glabrous portion testaceous.
Head with moderately large and rounded punctures. Pronotum with punctures slightly larger than on head. Scutellum with few impressed punctures. Elytral striae distinctly impressed; intervals flat and wide, irregular long setae on elytra; spine-like setae absent. Elytral apices entire and rounded in both sexes; highly bumpy along suture, depressed laterally on sides. Mesoventral process raised, rectangular in shape, with hood-like anterior tooth, posterior tooth large (Fig. 9c). Metaventral process widely rectangular, with large and deep elongate glabrous median depression; posterolateral portions bulge-like, posterior projection pointed. Abdominal ventrite 1 with median carina throughout the length. Abdominal ventrite 5 with rectangular emargination bearing broad and sharp median tooth (Fig. 9g). Meso- and metafemora with pubescence covering basal 0.7 of total length, borderline between pubescent and glabrous portion sinuate. Protarsus of male with adhesive soles on tarsomeres 1-2, tarsomeres 1-2 distinctly thickened, tarsomere 3 elongate; tarsomere 4 2× as long as tarsomere 3. Claws long, slender, slightly arched.
Male genitalia (Figs 9d-f). Phallobase ca. 0.7× total length of aedeagus. Parameres in lateral view wide basaly, narrowing subapically and apically projecting into rounded apex, lacking setae. Median lobe G-shaped in lateral view, with long basal projection directing apicad; apex wide, beak-shaped in lateral view.
Dominican Republic and Cuba. The species was until now considered as Cuban endemic (e.g.,
In our survey, the specimens of Berosus trilobus were collected usually in streams and rivers with stony or sandy bottom, clear water and with or without aquatic vegetation (Figs 11a, b), although once it has also been collected in a temporary pool with stony-muddy bottom, abundant organic matter, turbid water and rich submerged vegetation. Berosus trilobus is found in elevations ranging from sea level to ca. 850 m a.s.l.
http://species-id.net/wiki/Berosus_undatus
Figures 10a–i, 11“America meridionalis”.
Material examined. CUBA: Las Tunas: 1 ex. (in alcohol) (BSC-E): Las Cuarenta, 20°00'9.72"N, 76°57'48.6"W, 100 m a.s.l., 27.xi.2004, leg. Y. S. Megna, 00045. Granma: 2 exs. (in alcohol) (BSC-E): Cauto Cristo, Laguna permanente-II [permanent pool-II], 20°33'33.1"N, 76°28'44"W, 44 m a.s.l., 06.iii.2004, leg. L. Chávez, 00053. Santiago de Cuba: 4 exs. (in alcohol) (BSC-E): Laguna Juraguá, 19°56'30.8"N, 75°40'21.3"W, 22 m a.s.l, 17.ix.2003, leg. Y. S. Megna, 00044; 1 ex. (in alcohol) (BSC-E); 3 exs. (dry-mounted) (NMPC): Palma Soriano, Monte Barranca, 20°20'13.5"N, 76°1'11.6"W, 203 m a.s.l., 05.xii.2007, leg. A. Deler-Hernández, 00054; 1 ex. (dry-mounted) (NMPC): Palma Soriano 20°06'05"N, 75°58'44"W, 130 m a.s.l. 01.v.2005, leg. K. Blanco; 2 exs. (in alcohol) (BSC-E): La Maya, Los Reinaldos, Laguna temporal [temporal pool], 20°11'12"N, 75°31'43"W, 100 m a.s.l., 17.iii.2006, leg. Y. S. Megna, 00088. Guantánamo: 1 ex. (in alcohol) (BSC-E): Imías, Yacabo Abajo, Laguna temporal [temporal pool], 20°03'33.1"N, 74°42'29.9"W, 6 m a.s.l., 24.x.2008, leg. A. Deler-Hernández and S. Muñiz, 00060; 3 exs. (in alcohol) (BSC-E): San Antonio del Sur, Macambo, río Macambo, remanso [backwater], 20°03'26.9"N, 74°44'15.82"W, 4 m a.s.l., 25.x.2008, leg. A. Deler-Hernández and S. Muñiz, 00059.
Published Cuban records: Santiago de Cuba: Laguna Juraguá (

Berosus undatus (Fabricius, 1792). a habitus in dorsal view b habitus in lateral view c mesoventral process in lateral view d–f aedeagus (d dorsal view e lateral view f ventral view) g apex of male elytron h apex of female elytron i abdominal ventrite 5.
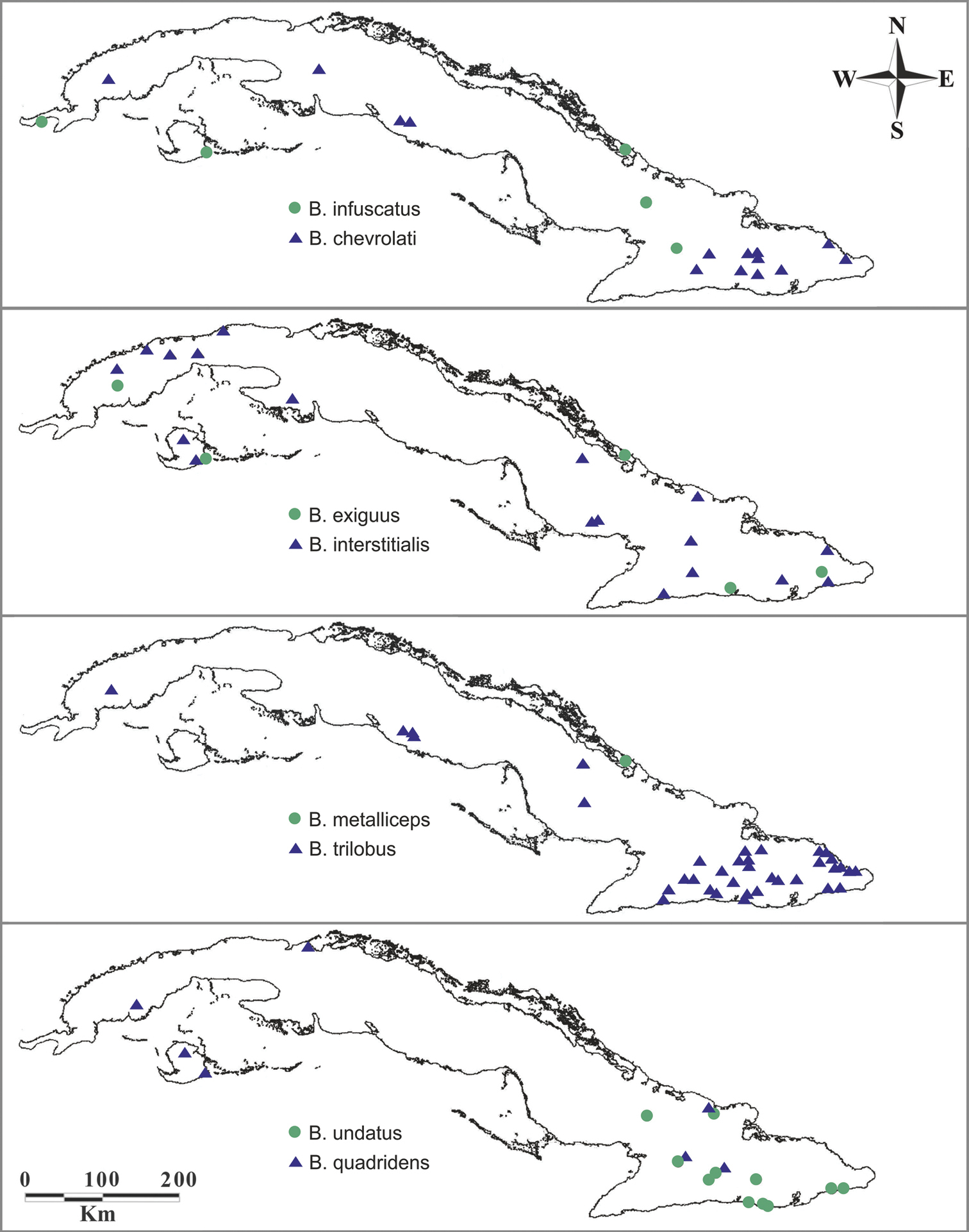
Known distribution of Berosus species in Cuba (includes our as well as historical records).

Localities of Cuban Berosus a Deler-Hernández collecting Berosus chevrolati and Berosus trilobus in a deep pool on side of a lowland river in Dos Caminos (June 2012) b locality of Berosus trilobus near Topes de Collantes (June 2010).
Habitus as in Figs 10a, b. Body length 6.3–7.2 mm. Head metallic black; pronotum pale with a pair of closely arranged elongate longitudinal black spots mesally, pronotal punctuation darkened; elytra pale with darkened striae and interval punctuation, plus with larger elongate dark spots on posterior half of intervals 1-4 and at midlength of intervals 8-10. Elytral apices with subapical spine in male (Fig. 10g), entire and rounded in female (Fig. 10h). Mesoventral process lowly laminar with large tooth directed posteriad (Fig. 10c). Abdominal ventrite 1 with median keel developed only between metacoxae. Emargination of abdominal ventrite 5 rectangular, with two broad and short medial teeth (Fig. 10i). Aedeagus (Figs 10d-f) with median lobe ca. as long as parameres, lateral margins of parameres subparallel except apically; median lobe narrow in dorsal view, slightly wider in lateral view.
USA (Texas), Mexico, Lesser Antilles (Hansen, 1999) and Cuba.
Cuban specimens were collected in permanent and temporary pools as well as from running waters with clear or turbid water, having or lacking aquatic vegetation.
| 1 | First abdominal ventrite carinate along its entire length or large part of it. Small to medium sized species (3.0-4.6 mm). Pronotum testaceous, with unpaired median black spot (Figs 1a, 9a) or without any dark spots (Fig. 2a) | 2 |
| – | First abdominal ventrite carinate only basally. Medium sized to large species (4.1-7.2 mm). Pronotum pale or testaceous with a pair of median black spots (Figs 3a, 4a, 6a, 7a, 10a), or pale without dark spots (Fig. 5a) | 4 |
| 2 | Head black. Pronotum and elytra with dark spots. Dorsal surface coarsely punctate. Mesoventral process subquadrate. First abdominal ventrite with median carina throughout the length. Median lobe of the aedeagus without subapical dorsal series of setae | 3 |
| – | Head testaceous; pronotum and elytra without dark spots (Fig. 2a). Meso-ventral process triangular (Figs 2b-c). First abdominal ventrite with median carina reaching the midlength. Aedeagus with median lobe bearing two series of long setae setae subapically on dorsal surface (Fig. 2e) | Berosus exiguus (Say) |
| 3 | Dark pronotal spot large and trilobate, narrow anteriorly and very wide posteriorly (Fig. 9a). Subapical area of each elytron forming a bump (Figs 9a-b). Apex of the median lobe beak-shaped in lateral view, basal projection of the median lobe long (Fig. 9e) | Berosus trilobus Chevrolat |
| – | Dark pronotal spot narrow, situated mesally, not widened posteriad (Fig. 1a). Subapical area of each elytron without a distinct bump (Figs 1a-b). Apex of the median lobe rounded in lateral view, basal projection of the median lobe short (Fig. 1e). | Berosus chevrolati Zaitzev |
| 4 | Head metallic black to black. First abdominal ventrite without lateral depressions, emargination of abdominal ventrite 5 rectangular, without distinct sexual dismorphism. Parameres separated from each other. Elytral apex with or without subapical spine | 5 |
| – | Head testaceous (except mesally in some cases, Figs 7a–b). First abdominal ventrite with lateral depressions; emargination of abdominal ventrite 5 deeply or shallowly circular, sexually dimorphic (Figs 7f–g). Parameres joined mesally into a common dish-like structure (Figs 8e–h). Elytral apex with subapical spine in both sexes (Figs 7c–d) | Berosus quadridens Chevrolat |
| 5 | Pronotal disc without spots (Fig. 5a) or with small submesal anterior spots (Fig. 6a), never with a pair of mesal elongate large dark spots throughout the pronotal length. Apical emargination of abdominal ventrite 5 without tooth (Fig. 5g) or with a single medial tooth (Fig. 6g). Median lobe longer that parameres | 6 |
| – | Pronotal disc with a pair of narrow elongate metallic central black spots. Apical emargination of abdominal ventrite 5 with two medial teeth. Median lobe shorter that parameres | 7 |
| 6 | Elytral striae distinctly darkened, elytral disc without numerous darker spots (Figs 5a–b). Apical emargination of abdominal ventrite 5 without median tooth (Fig. 5g). Median lobe of the aedeagus very long, spatulate apically in dorsal view, sinuate in lateral view (Figs 5d–f) | Berosus metalliceps Sharp |
| – | Elytral series not darkened, each elytron with several darker spots on the disc (Figs 6a-b). Apical emargination of abdominal ventrite 5 with a broad and short median tooth (Fig. 6g). Median lobe of the aedeagus slender and apex, pointed in dorsal view, slightly arcuate in lateral view (Figs 6d–f) | Berosus peregrinus (Herbst) |
| 7 | Body size less than 6.0 mm. Elytral apices entire (without subapical spines) in both sexes. Phallobase longer than a half of total length of the aedeagus | 8 |
| – | Body size more than 6.3 mm. Elytral apices sexually dimorphic, with subapical spine in males (Fig. 10g) and rounded in females (Fig. 10h). Phallobase shorter than a half of the total length of the aedeagus (Figs 10d–f) | Berosus undatus (Fabricius) |
| 8 | Pronotum without mesh-like microsculpture on interstices. Mesoventral process with very small tooth (Fig. 4c). Posterolateral angles of metaventral process triangular. Aedeagus narrow, lateral margins of parameres subparallel, base of each paramere with a conspicuous tooth (Figs 4d–f) | Berosus interstitialis |
| – | Pronotum with strong mesh-like microsculpture on interstices. Mesoventral process with larger tooth (Fig. 3c). Posterolateral angles of metaventral process with rounded laminae. Parameres sinuate along lateral margins, base of parameres without conspicuous teeth (Figs 3d–f) | Berosus infuscatus |
We thank Yoandri S. Megna (Universidad de Oriente, Santiago de Cuba) for his help during the field collecting and sharing of some specimens for this study, Andrew Short (Kansas University, Lawrence, U.S.A.) for his help with identification of the specimens, providing the literature, valuable discussions and numerous corrections of this manuscript, and Bruno Clarkson Mattos (Universidade do Brasil, Rio de Janeiro, Brasil) for numerous corrections of the text and comments on the morphology of male genitalia. We are deeply grateful to the authorities and personnel of the Cuban Ministry of Science, Technology and Environment (CITMA) and the Empresa Nacional para la Protección de la Flora y la Fauna (ENPFF) for providing access to protected areas under their control. The work of M. Fikáček was partly supported by the Ministry of Culture of the Czech Republic (DKRVO 2012 and DKRVO 2013/12, National Museum, 00023272), the visit of the first author to Prague was supported by the institutional resources of the Ministry of Education, Youth and Sport of the Czech Republic. Open access to this paper was supported by the Encyclopedia of Life (EOL) Open Access Support Project (EOASP).