





Citation: Yoshimura M, Fisher BL (2014) A revision of the ant genus Mystrium in the Malagasy region with description of six new species and remarks on Amblyopone and Stigmatomma (Hymenoptera, Formicidae, Amblyoponinae). ZooKeys 394: 1–99. doi: 10.3897/zookeys.394.6446
The genus Mystrium is revised for the Malagasy region. Six species, Mystrium barrybressleri sp. n., Mystrium labyrinth sp. n., Mystrium eques sp. n., Mystrium mirror sp. n., Mystrium shadow sp. n., and Mystrium janovitzi sp. n. are described as new. Two existing names, Mystrium fallax Forel and Mystrium stadelmanni Forel, are synonymized with Mystrium voeltzkowi Forel and Mystrium mysticum Roger, respectively. All recognized species, including species outside of the Malagasy region, are assigned to one of the three newly proposed species groups. The associations between existing names and males are reexamined, and males of eight of the ten Malagasy species are described or redescribed. The taxonomic history of Mystrium highlights the importance of using unique identifiers when designating type specimens and the use of deposited vouchers in phylogenetic and ecological studies. Keys to species for workers, queens, and males are provided. Furthermore, a neotype for Mystrium mysticum is designated, as well as lectotypes for Mystrium camillae Emery, Mystrium rogeri Forel, Mystrium fallax Forel, Mystrium oberthueri Forel, Mystrium stadelmanni Forel, and Mystrium voeltzkowi Forel. Stigmatomma gingivale (Brown) is reassigned to Amblyopone as comb. rev. and Amblyopone awa Xu & Chu, Amblyopone kangba Xu & Chu, Amblyopone meiliana Xu & Chu, and Amblyopone zomae Xu & Chu are transferred to the genus Stigmatomma as comb. n.
Major, minor, ergatoid queen, intercaste, intermediate, male, Madagascar, Comoros, Mayotte, mandible evolution, labral dentiform setae, genitalia, Amblyoponini, XMAS clade, Onychomyrmex, Apomyrma
The genus Mystrium Roger, 1862 was originally described from Madagascar, and ten species names were regarded as valid prior to this revision (Fig. 1). The Malagasy region has an incomparably abundant and rich Mystrium fauna with over half of the species endemic to the Malagasy region. After Roger established Mystrium with a single species, Mystrium mysticum Roger, 1862, Forel described five additional species in the Malagasy region between 1895 and 1899 (

Colonies of Mystrium. A Mystrium oberthueri B Mystrium voeltzkowi. A a cocoon and a worker B larvae, workers, and ergatoid queens. Photos by Alex Wild.
Recent research in Madagascar demonstrated that the previous taxonomic works were insufficient to diagnose the species boundaries of Mystrium in the Malagasy region. Specimens collected from a single colony showed remarkable morphological variations within each existing species, and these variations rendered most published species-level diagnostic characters insufficient to delimit species. A reassessment of species boundaries was also needed in light of recent ecological and phylogenetic studies. Ecological studies reported various reproductive systems across Mystrium species, including typical winged queens, short-winged queens (
In this study, we revise the species-level taxonomy for the genus Mystrium in the Malagasy region. Comprehensive morphological and distributional examinations using recently accumulated material enabled us to recognize ten species in this region; six species are new, and two existing names, Mystrium fallax and Mystrium stadelmanni, are synonymized with Mystrium voeltzkowi and Mystrium mysticum, respectively. We propose a new species-group system, and all recognized species, including extralimital species, are assigned to one of the three species groups. Both workers and queens of existing Malagasy species are redescribed with new diagnostic characters. Associations between existing names and males are reexamined, and the males of eight of the ten Malagasy species of Mystrium are described or redescribed. Identification keys to species for workers, queens, and males are provided. Furthermore, a neotype for Mystrium mysticum is designated, as well as lectotypes for Mystrium camillae, Mystrium rogeri, Mystrium fallax, Mystrium oberthueri, Mystrium stadelmanni, and Mystrium voeltzkowi. Though we do not include species outside of the Malagasy region in this revision, we provide taxonomic remarks on Mystrium camillae to better define species in the camillae group from the Malagasy region.
In addition to the species-level taxonomy of Mystrium, we also propose new genus-level diagnostic characters of Mystrium. In our previous work (
BMNH The Natural History Museum (formerly British Museum, Natural History), London, U.K.
CASC California Academy of Sciences, San Francisco, California, U.S.A.
MCZC Museum of Comparative Zoology, Cambridge, Massachusetts, U.S.A.
MHNG Muséum d’Histoire Naturelle de la Ville de Genève, Geneva, Switzerland
MNHN Muséum National d’Histoire Naturelle, Paris, France
MSNG Museo Civico di Storia Naturale “Giacomo Doria”, Genova, Italy
NHMB Naturhistorisches Museum, Basel, Switzerland
PBTZ Parc Botanique et Zoologique de Tsimbazaza, Antananarivo, Madagascar
ZMHB Museum für Naturkunde der Humboldt-Universität zu Berlin, Berlin, Germany
Materials. The material on which this work is based was collected during arthropod surveys in Madagascar and the nearby islands in the Southwest Indian Ocean conducted by B. Fisher and the Malagasy ant team from the Madagascar Biodiversity Center in Antananarivo, Madagascar. Their work in the region includes more than 6, 000 leaf litter samples, 4, 000 pitfall traps, 1, 000 Malaise trap collections, and 9, 000 additional hand collection events throughout Madagascar from 1992 through 2013 (see
We used a series of specimens collected from a single colony to determine the associations among queens, workers, and males and the extent of within-colony variation. When it was impossible to find all sexes and castes within the same colony sample, the associations based on morphological characters were confirmed by COI barcoding analysis (Fisher unpublished data).
Methods. Observations and dissections were carried out under stereoscopic microscopes (LEICA M125). Digital color images were created using a LEICA DFC425 digital camera. LEICA Application Suite software (ver. 3.8) was used for images, and a compound microscope (Leica DM4000M) and Nikon digital camera (DXm1200) and Helicon Focus version 4.10.2 software were used as well. The images were edited in Adobe Photoshop and Adobe Illustrator. Each imaged or dissected specimen is uniquely identified with a specimen-level unique identifier (e.g. CASENT0003099).
Measurements and indices. The following measurements and indices (illustrated in Fig. 2) are cited in the text. The values are presented in mm.
Head length (HL), maximum length of head in full-face view between lines drawn across anterior margin of clypeus and posterior margin of head (worker, queen, male; including ocelli in male and queen).
Head width (HW), maximum width of head in full-face view, including eyes (worker, queen, and male).
Head depth (HD), maximum depth of head in lateral view measured perpendicular to full-face view plane (worker).
Mandible length (ML), maximum length of dorsal surface of the mandible (worker and queen).
Scape length (SL), length of scape excluding radicle (worker, queen, and male).
Eye length (EL), maximum length of eye measured in lateral view (queen and male).
Weber’s length of mesosoma (WL), maximum diagonal distance in lateral view, from base of anterior slope of pronotum to metapleural lobe (worker, queen, and male).
Pronotal width (PnW), maximum width of pronotum in dorsal view (worker).
Mesonotal width (MnW), maximum width of mesonotum in dorsal view (queen, and male).
Propodeal width (PpW), maximum width of propodeum in dorsal view (worker).
Petiolar length (PtL), maximum length of petiolar node in dorsal view (worker and queen).
Petiolar width (PtW), maximum width of petiolar node in dorsal view (worker and queen).
Cephalic index (CI), HW/HL ×100 (worker, queen, and male).
Mandible index (MI), ML/HW ×100 (worker and queen).
Scape index (SI), SL/HW ×100 (worker, queen, and male).
Eye index (EI), EL/HL ×100 (queen and male).
Mesonotal index (MnI), MnW/HW ×100 (queen and male).
Petiolar index (PtI), PtW/PtL ×100 (worker and queen).
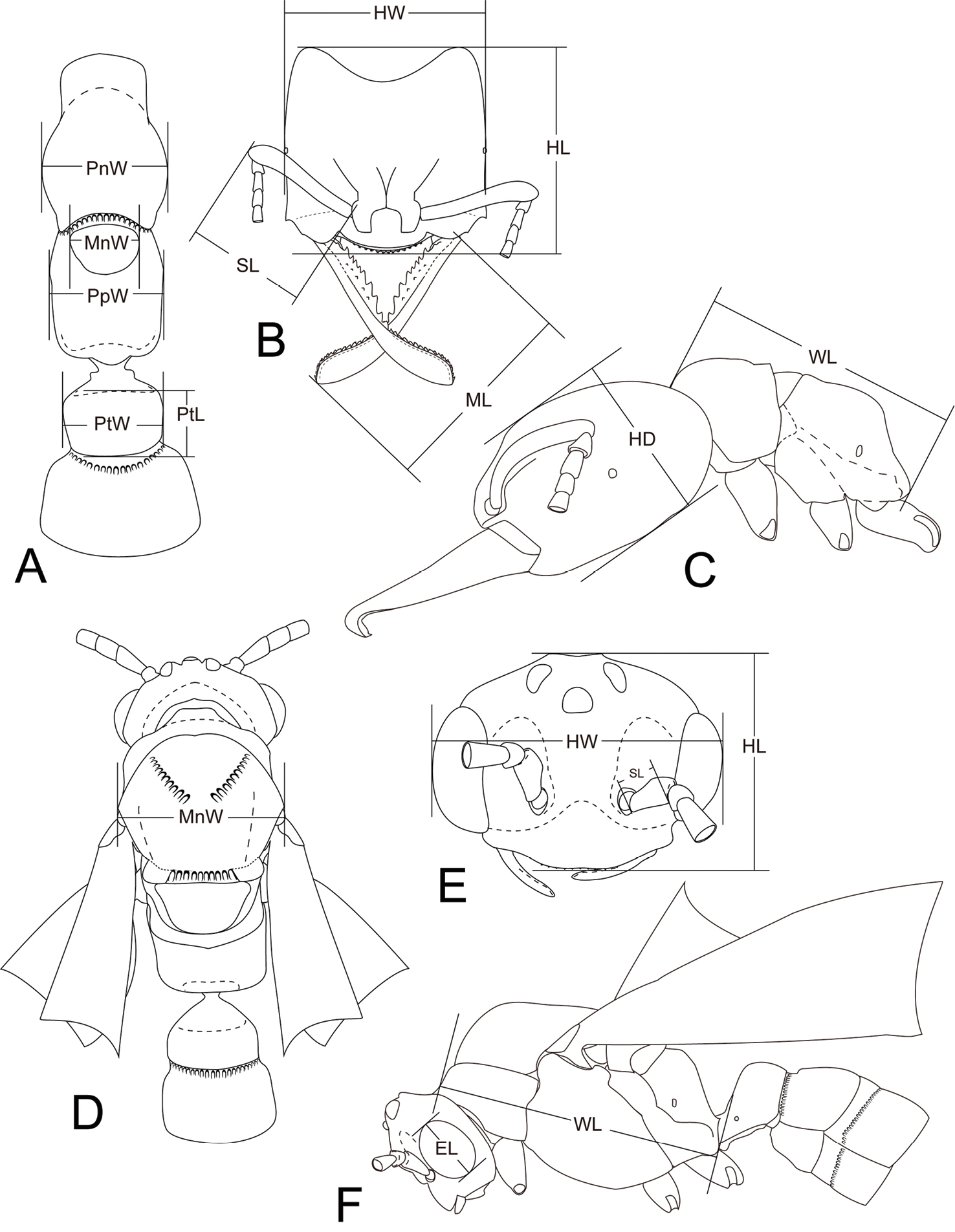
Measurements for Mystrium species. A, B, C worker D, E, F male. A, D body in dorsal view B, E head in full-face view C, F body in lateral view. Abbreviations are explained in Measurements and indices section.
Terminology. Morphological terminology for workers and ergatoid queens follows
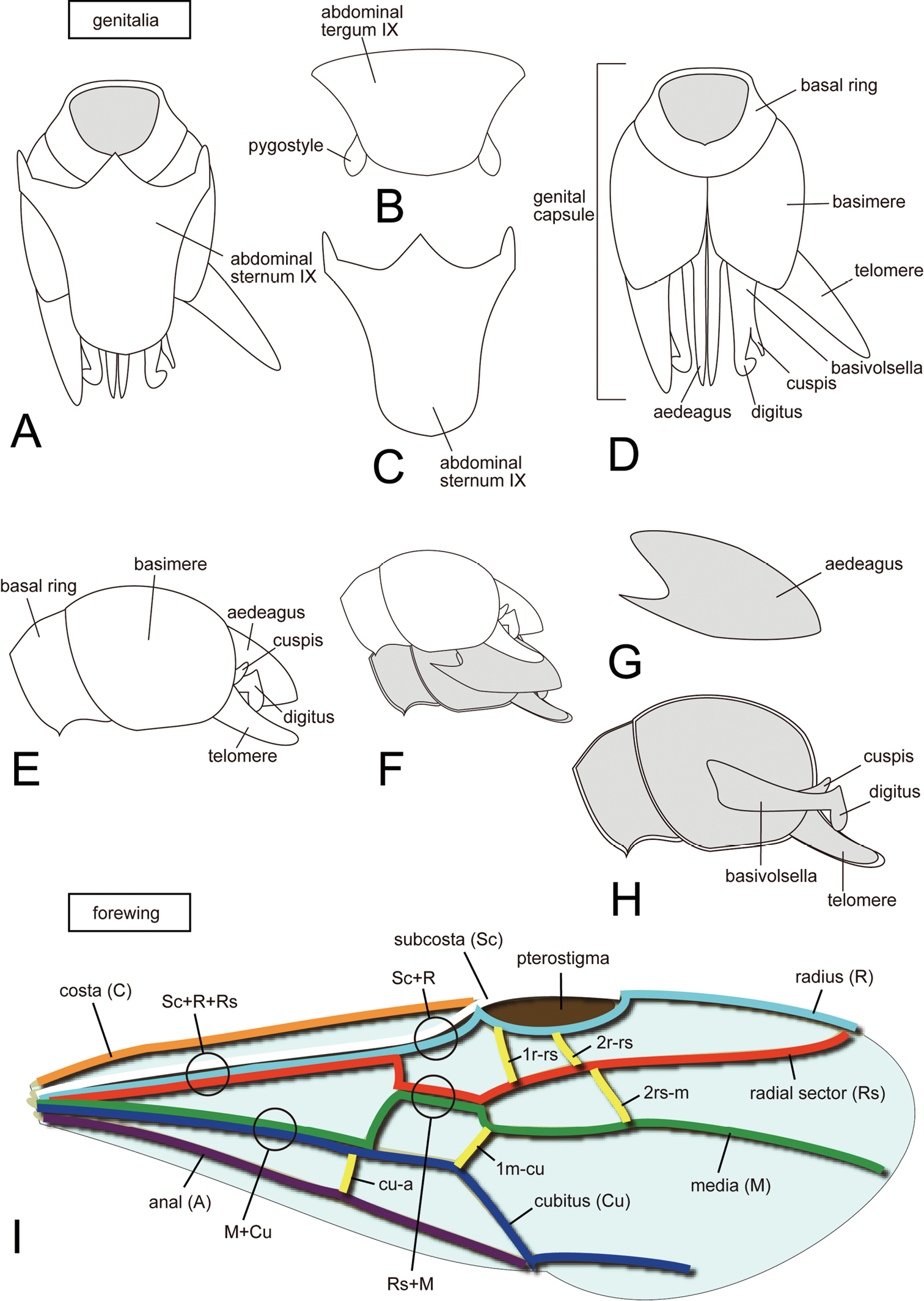
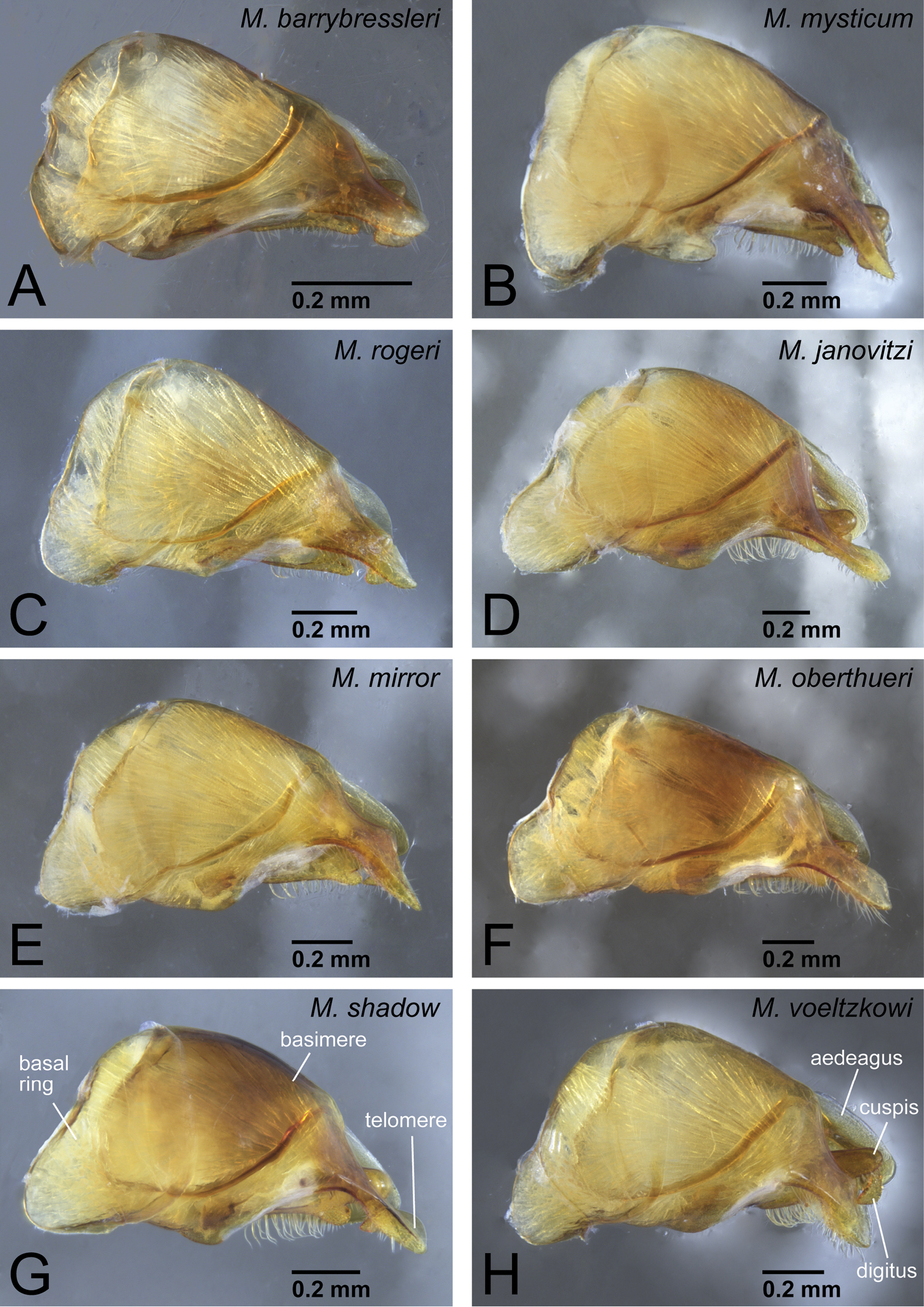
Terminology for male genitalia and forewing of ants. A, D, E, F, H genital capsule B abdominal tergum IX C abdominal sternum IX G aedeagus I forewing. A, C, D in ventral view B, I in dorsal view E–H in lateral view.
We replaced one genital term, telomere, with harpago in an earlier work (
Terminology for caste names. We classify females into three phenotypes: winged gyne, large wingless phenotype, and small phenotype with or without vestigial wings. Some species lack the winged gyne, and small phenotype acts as a reproductive cast; some species have intermediates between winged gyne and large phenotype. The presence or absence of each caste, and the role of each phenotype are consistent within species groups defined below, but vary among the species groups (Table 1). In this study, we use “queen” in reference only to the winged gyne; intercaste for intermediates between winged gyne and large wingless phenotype; major worker for the large wingless phenotype and minor worker for the small phenotype when the small phenotype does not function as a reproductive; worker for the large wingless phenotype and ergatoid queen for small phenotype when the small phenotype functions as a reproductive. The term female is used as a collective noun to indicate all female phenotypes. Males often display remarkable intra-specific variation in size and shape; however, no clear morphological division among those variations is recognized. Relationships between caste and species groups are given in Table 1.
Terminology for geographical regions. Terminology and definition for geographical regions follows
Distribution of castes and phenotypes across species groups. Caste and the function of phenotypes vary across species groups. Caste names are explained in the Terminology section. Morphological characters defining phenotypes are given in the description of each species group.
| female | male | ||||
|---|---|---|---|---|---|
| winged gyne | intermediates | large wingless phenotype | small phenotype | ||
| camillae species group | queen | - | major worker | minor worker | male |
| mysticum species group | queen | intercaste | major worker | minor worker | male |
| voeltzkowi species group | - | - | worker | ergatoid queen | male |
http://species-id.net/wiki/Mystrium
The characters uniquely observed in Mystrium within the subfamily Amblyoponinae are given in italics.
-
Compound eye present.
-
Posterior margin of head strongly expands posteriorly on each side.
-
Anterior margin of clypeus with specialized conical setae.
-
Spatulate setae present on clypeus mesal of mandible insertion (Fig. 4A).
-
Labrum lacking small dentiform setae arranged horizontally.
-
Palpal formula 4, 3.
-
Distinct extension present on the distal edge of second labial palpomere (Fig. 4B).
-
Mandible linear without a distinct basal angle.
-
Masticatory margin of mandible with two rows of projections: true mandibular teeth on dorsal row and a series of basal denticles on the ventral row; basal ventral denticles larger than true mandibular teeth except for apical teeth.
-
At midlength of mandible, row of basal denticles arranged on ventral edge of mandibular shaft, distant from mandibular teeth (Fig. 4C).
-
Mandible twisted inward so that apical tooth is located ventrally (Fig. 4D).
-
No basal large projection present on basal portion of mandibular inner margin.
-
Ridge present on apical portion of mandible, which is originally dividing ventral and inner surfaces of mandible, inserting dorsally to the apical tooth (Fig. 5A vs. 5B: Stigmatomma mystriops Brown, 1960, see comments).
-
Constriction between petiole and abdominal segment III present.
-
Cinctus 3, constriction between pre- and post-sclerites on abdominal segment IV, distinctly present.
-
One or two stout spines present on posterior portion of abdominal sternum VII (Fig. 5C).
-
Body surface with spatulate to squamose setae in some workers of all species.
The diagnostic characters uniquely observed in Mystrium within the subfamily Amblyoponinae are given in italics.
-
Frontal carinae present.
-
Anterior margin (=free margin) of clypeus with specialized conical setae.
-
Antenna consisting of 13 segments.
-
Mandible with single, blunt apical tooth.
-
Palpal formula 4, 3.
-
Notaulus distinct or absent.
-
Mesepimeron often lacking distinct posterodorsal lobe (epimeral lobe).
-
Mesotibia with one or two spurs in most cases, rarely indistinct.
-
Metatibia with two spurs.
-
Distinct constriction present between petiole and abdominal segment III in dorsal view.
-
Abdominal segment IV with tergosternal fusion.
-
Pretergite of abdominal segment IV distinctly differentiated from posttergite, with the cinctus between them.
-
Pygostyles absent.
-
Distal margin of abdominal sternum IX convex.
-
Separation between basimere and telomere distinct.
-
Basal projection on cuspis well developed.
-
Basoventral portion of the aedeagus in lateral view extended basally, distal margin of extension rounded (Fig. 5D).
-
Serrate denticles present on basal portion of ventral margin of aedeagus in lateral view.
-
Pterostigma well developed on forewing.
-
Radial sector on forewing fully present.
-
Radial sector on forewing reaches costal margin.
-
2r-rs on forewing connected with radial sector posterior to pterostigma.
-
2rs-m present on forewing.
-
Position of cu-a on forewing variable, close to or far from junction between media and cubitus.
-
Radius present on hindwing.
-
1rs-m present on hindwing.
-
Media on hindwing present apical to 1rs-m.

Generic diagnostic characters of Mystrium. A Mystrium rogeri (CASENT0001069) B Mystrium voeltzkowi (CASENT0317584) C Mystrium mysticum (CASENT0429965) D Mystrium janovitzi (CASENT0482696). A, B, C worker D ergatoid queen. A clypeus and mandible in oblique anterior view B mouthparts in oblique anterior view (left galea is omitted) C mandible in oblique lateral view D head in lateral view.
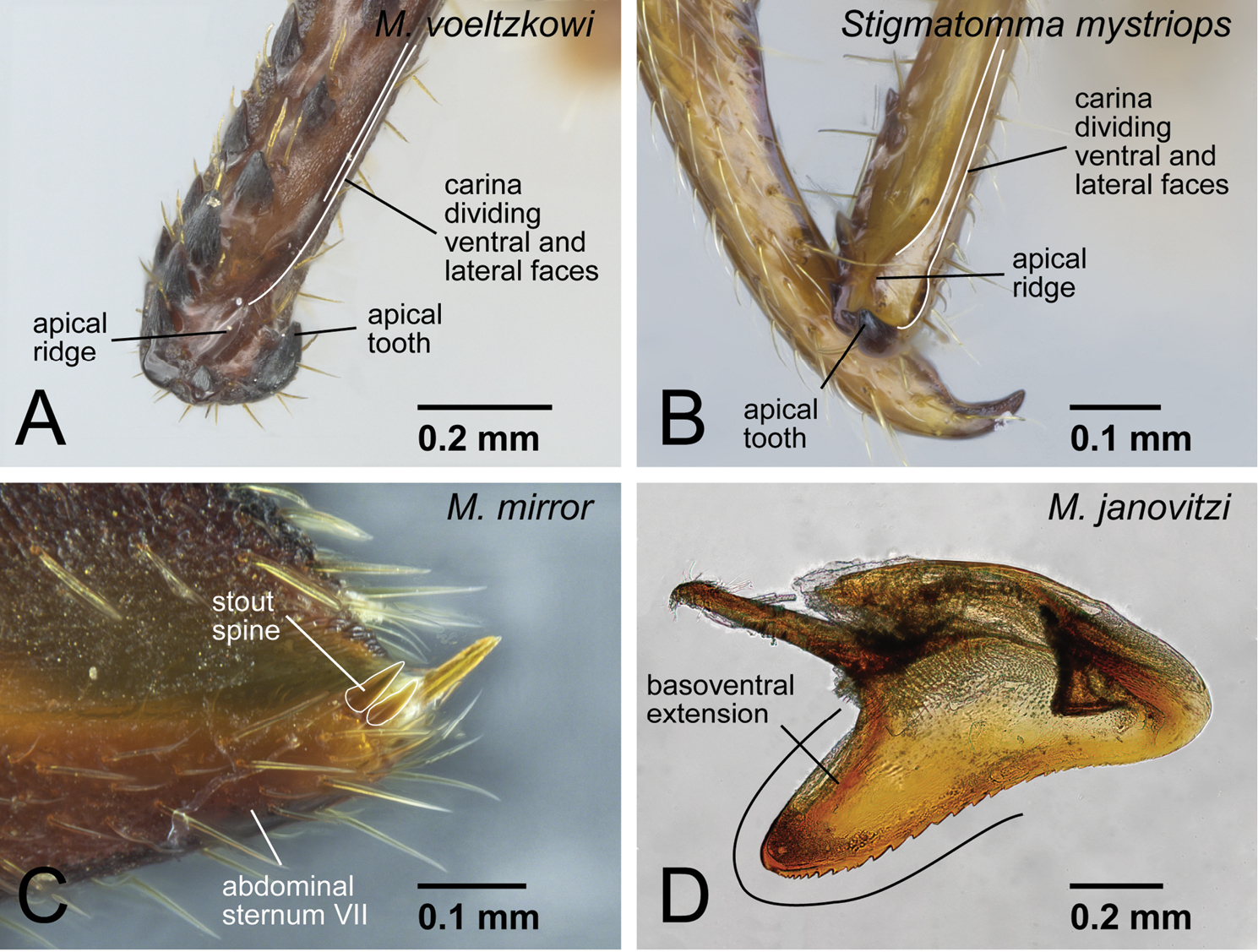
Generic diagnostic characters of Mystrium and the differences from Stigmatomma. A Mystrium voeltzkowi (CASENT0002075) B Stigmatomma mystriops (INB0003690672) C Mystrium mirror (CASENT0317582) D Mystrium janovitzi (CASENT0080644). A, B, C worker D male A, B mandible in oblique mesal view C abdominal segment VII in lateral view D aedeagus in lateral view.
The workers of the genus Mystrium can be distinguished easily from those of the other amblyoponine genera by their characteristic head shape and mandibles (Fig. 1). Mystrium has a wide head with posterior margin strongly concave, mandible linear and longer than head with rounded apex in full-face view, and two separated rows of “teeth” on the masticatory margin. In this study, we propose further detailed characters as the result of our comparative study. All Mystrium females share six characters unique to this genus which distinguish it from the other amblyoponine genera: spatulate setae on clypeus mesal of the mandible insertion (character 4: Fig. 4A), a distinct extension on the distal edge of the second segment on the labial palp (character 6: Fig. 4B), mandibular teeth distant from basal denticles (character 10: Fig. 4C), twisted mandible (character 11: Fig. 4D), a ridge on the apical portion of the mandible inserting dorsally to the apical tooth (character 13: 5A), and one or two lateral spines on abdominal sternum VII (character 16: 5C). Character 10, mandible teeth and basal denticles arranged in two rows (Fig. 4C), was used in
In addition to characters unique to Mystrium, we discuss two generic characters (characters 5 and 9) that are useful for distinguishing amblyoponine genera. Neither character 5 nor 9 distinguish Mystrium from the other XMAS genera; however, both of these characters do distinguish Mystrium from Amblyopone.
Character 5 is provisionally proposed here to separate Amblyopone from genera in the XMAS clade. In the XMAS clade, the basal part of the labrum is lacking small dentiform setae (Fig. 6A). However, in Amblyopone, small dentiform setae are present on the labrum (Fig. 6B). Mandible character 9 proposed in
The differences in mandibular characters between species in the XMAS clade and those in Amblyopone are associated with the parts of the mandible used for catching their prey. We assume that the masticatory margin is the main functional part of the mandible in XMAS clade species (Figs 6C, 6E), while the whole mandibular shaft is used for this purpose in Amblyopone (Figs 6D, 6F). The presence or absence of dentiform setae on the labrum (character 5: Figs 6B vs. 6A) supports our assumption. Amblyopone species probably “hold” prey using teeth along the whole shaft of the mandible (Fig. 6F), conical setae on the flat clypeus, and dentiform setae on the labrum (Fig. 6B); while species of Stigmatomma (in XMAS clade) “pinch” prey using their masticatory margin (distal part) of the mandible (Fig. 6E), and support it using anterior clypeal conical setae strongly extended anteriorly (Fig. 6A). When the mandibles of both groups elongated under these two use conditions, their morphology adapted differently. The mandibular shaft in Amblyopone became wider and stouter (Fig. 6F), while the basal denticles in XMAS clade became larger than true teeth and distinctly directed basally (Fig. 6E).
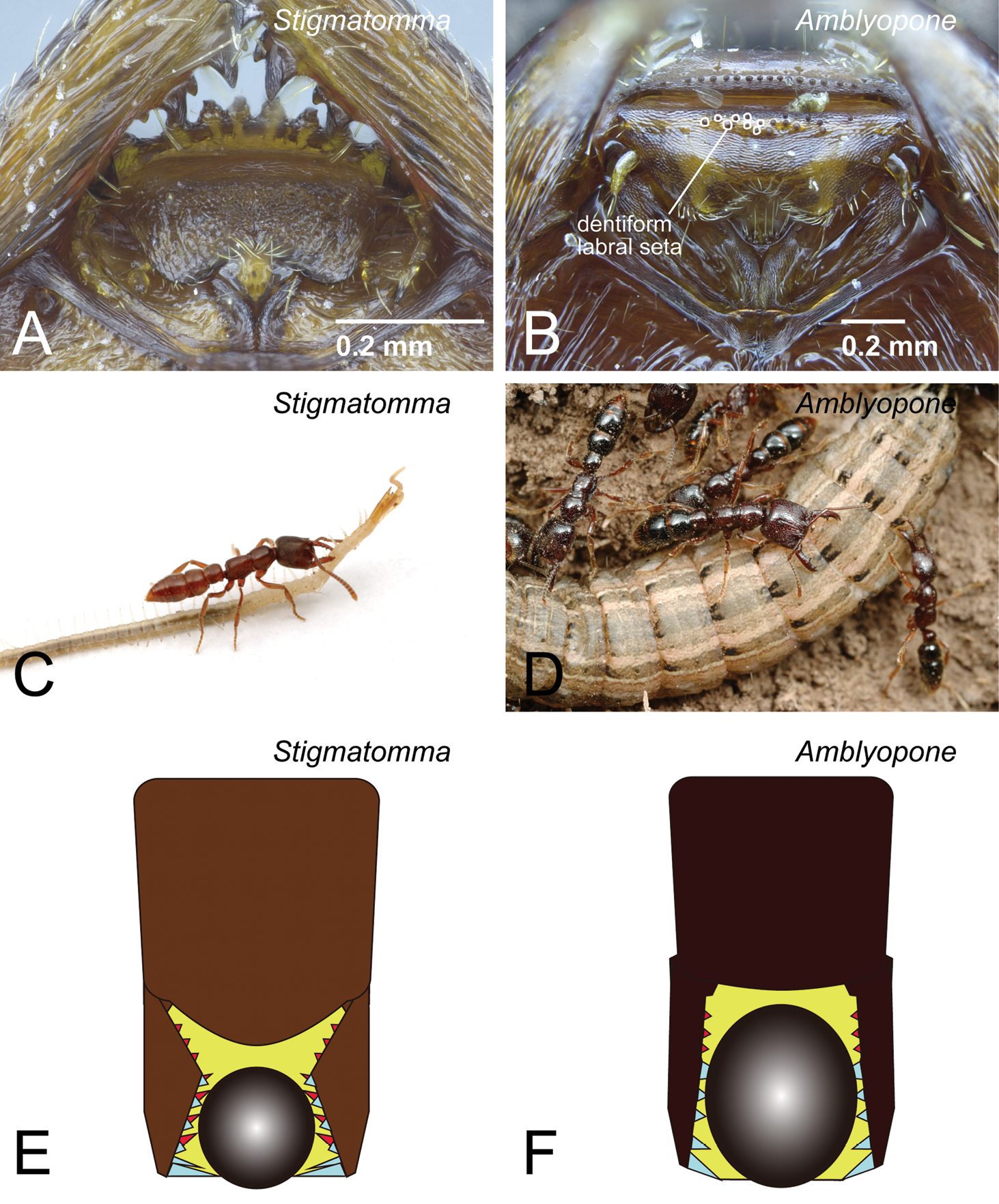
Differences in characters of the worker mandible and labrum between XMAS clade genera and Amblyopone. A, C, E XMAS clade genera (Stigmatomma) B, D, F Amblyopone. A Stigmatomma pallipes (CASENT0247077) B Amblyopone australis (CASENT0100424) C Stigmatomma oregonense D Amblyopone australis. A, B labrum C, D usage of the mandibles E, F pattern diagrams for mandibular use. A dentiform labral seta is absent B dentiform labral setae are present C, E the masticatory margin is the main functional part of the mandible in XMAS clade species D, F the whole mandibular shaft is the functional part of the mandible in Amblyopone. C, D Photos by Alex Wild.
Because of the presence of the dentiform labral setae, Stigmatomma gingivale is retransferred to Amblyopone as Amblyopone gingivalis Brown, 1960 comb. rev., although
On the other hand, on the basis of the mandible characters, Amblyopone awa, Amblyopone kangba, Amblyopone meiliana, and Amblyopone zomae are transferred to the genus Stigmatomma as Stigmatomma awa (Xu & Chu, 2012), comb. n., Stigmatomma kangba (Xu & Chu, 2012), comb. n., Stigmatomma meilianum (Xu & Chu, 2012), comb. n., and Stigmatomma zomae (Xu & Chu, 2012), comb. n., respectively. According to the original description (Xu and Chu 2012), the mandibular characters in figures 29, 34, and 52 show that Stigmatomma zomae, Stigmatomma meilianum, and Stigmatomma awa have typical forms observed in Stigmatomma, although we did not examine actual specimens of any of these four Chinese species.
The male diagnostic characters used in this revision follow those in
In this study, we divide the species of Mystrium into the three species groups listed below. The subdivisions of the genus are useful because of the difficulty of species-level identification.
camillae species group: Alate queen present (Table 1). Dorsal surface of pronotum covered with regular reticulation in worker. Second maxillary palpomere shorter than the third in the Malagasy species. Labrum in females gently convex but without median longitudinal carina. Two lateral spines present on abdominal sternum VII in females. Whole vertex sculptured and not smooth in worker. Petiole in dorsal view in females relatively wide (PtI>210 in queen; PtI>180 in worker). Known from the Afrotropical, Malagasy, Indomalaya, and Australian regions.
mysticum species group: Alate queen present (Table 1). Dorsal surface of pronotum covered with somewhat irregular striae, and central part of pronotal dorsum covered with fine striae in worker. Second maxillary palpomere longer than the third. Labrum in females with longitudinal carina and strongly convex on its central portion. Single lateral spine present on abdominal sternum VII in females. Ventral area of vertex in females not sculptured, smooth. Petiole in dorsal view in females relatively long (PtI<189 in queen; PtI<183 in worker). Known only from the Malagasy region.
voeltzkowi species group: Only ergatoid queen, which is relatively smaller than worker, present; alate queen absent (Table 1). Dorsal surface of pronotum covered with somewhat irregular longitudinal striae in worker. Second maxillary palpomere longer than the third. Labrum in females gently convex but without median longitudinal carina. Two lateral spines present on abdominal sternum VII in females. Whole vertex in females sculptured and not smooth. Petiole in dorsal view relatively long (PtI<195 in queen; PtI<180 in worker). Known only from the Malagasy region.
A list of Mystrium species is given below. Sexes and castes known in each species are given in brackets as: w, worker; aq, alate queen; eq, ergatoid queen; m, male. In addition to the species listed below, we recognize two undetermined morphospecies in the collection and provide notes for these taxa in the remarks of the camillae species group. The associations between the species names and the species codes used in previous papers are also discussed in the remarks for each species.
-
camillae species group
-
Mystrium barrybressleri sp. n. [w, aq, m] Madagascar
-
Mystrium camillae Emery, 1889 [w, aq, m] South-East Asia, Australia
-
=Mystrium javana Karavaiev, 1925 [w: minor]
-
=Mystrium oculatum Xu, 1998 [w, aq]
-
Mystrium labyrinth sp. n. [w, aq] Madagascar
-
Mystrium leonie Bihn & Verhaagh, 2007 [w] Indonesia
-
Mystrium maren Bihn & Verhaagh, 2007 [w] Indonesia
-
Mystrium silvestrii Santschi, 1914 [w] Africa
-
-
mysticum species group
-
Mystrium mysticum Roger, 1862 [w, aq, m] Madagascar, Comoros
-
=Mystrium stadelmanni Forel, 1895 syn. n.
-
Mystrium rogeri Forel, 1899 [w, aq, m] Madagascar, Comoros
-
-
voeltzkowi species group
-
Mystrium eques sp. n. [w, eq] Madagascar
-
Mystrium janovitzi sp. n. [w, eq, m] Madagascar
-
Mystrium mirror sp. n. [w, eq, m] Madagascar
-
Mystrium oberthueri Forel, 1897 [w, eq, m] Madagascar
-
Mystrium shadow sp. n. [w, eq, m] Madagascar
-
Mystrium voeltzkowi Forel, 1897 [w, eq, m] Madagascar, Mayotte
-
=Mystrium fallax Forel, 1897 [eq] syn. n.
-
| 1 | Wings present and well developed. Wing sclerites fully developed even with wings removed (Fig. 7A). Both anterior and lateral ocelli distinct (Fig. 7C) | 2 |
| – | Wings absent or vestigial. Wing sclerites undeveloped or incomplete (Fig. 7B). Lateral ocelli absent (Fig. 7D) | 5 |
| 2 | Labrum with distinct longitudinal ridge on its middle portion (Fig. 8A). Distal portion of abdominal sternum VII with single pair of stout spines (Fig. 8C). Body size relatively large (HW>1.9 mm) | 3 (mysticum species group, part) |
| – | Labrum without distinct longitudinal ridge on its middle portion (Fig. 8B). Distal portion of abdominal sternum VII with two pairs of stout spines (Fig. 8D). Body size relatively small (HW<1.7 mm) | 4 (camillae species group, part) |
| 3 | Setae (=hairs) on dorsal surface of head and pronotum simple (Fig. 9A). Anterior margin of clypeus convex (Fig. 9C) | Mystrium mysticum Roger (queen and intercaste) |
| – | Setae (=hairs) on dorsal surface of head and pronotum spatulate (Fig. 9B). Anterior margin of clypeus straight or weakly concave (Fig. 9D) | Mystrium rogeri Forel (queen and intercaste) |
| 4 | Petiole short in dorsal view, 0.5× length of abdominal segment III (Fig. 10A). Setae on pronotal dorsum narrow and sharp on its apex. First flagellomere (third segment of antenna) distinctly shorter than pedicel (second segment of antenna) (Fig. 10C). Body yellow to yellowish brown | Mystrium barrybressleri sp. n. (queen) |
| – | Petiole long in dorsal view, 0.7× length of abdominal segment III (Fig. 10B). Setae on pronotal dorsum spatulate and dull on its apex. First flagellomere as long as pedicel (Fig. 10D). Body blackish brown | Mystrium labyrinth sp. n. (queen) |
| 5 | Ventral half of vertex unsculptured, smooth (Fig. 11A). Labrum with distinct longitudinal ridge on its middle portion (as in Fig. 8A). Distal portion of abdominal sternum VII with single pair of stout spines (Fig. 8D). Anterior ocellus sometimes present | 6 (mysticum species group, worker and intercaste) |
| – | Ventral half of vertex sculptured (Fig. 11B). Labrum without distinct longitudinal ridge on its middle portion (as in Fig. 8B). Distal portion of abdominal sternum VII with two pairs of stout spines (Fig. 8D). Ocelli absent | 7 |
| 6 | Anterior margin of clypeus convex (as in Fig. 9C). Among same-sized individuals, combination of following characters usually observed: sculpture relatively shallow and weak on lateral surface of pronotum; difference in width of dorsal surface of mandible larger between mandibular shaft (narrowest part) and distal portion (widest part) (Fig. 12A); petiole in dorsal view relatively wider | Mystrium mysticum Roger (worker and intercaste) |
| – | Anterior margin of clypeus straight or weakly concave (as in Fig. 9D). Among same sized individuals, combination of following characters usually observed: sculpture relatively deep and strong on lateral surface of pronotum; difference in width of dorsal surface of mandible smaller between mandibular shaft (narrowest part) and distal portion (widest part) (Fig. 12B); petiole in dorsal view relatively narrower | Mystrium rogeri Forel (worker and intercaste) |
| 7 | Head dorsum and pronotum strongly to weakly and regularly reticulate (Fig. 13A). No deep longitudinal furrow on midline of pronotum. Second maxillary palpomere shorter than third (Fig. 13C) | 8 (camillae group, worker) |
| – | Head dorsum and pronotum finely to roughly sculptured longitudinally, but not regularly reticulate (Fig. 13B). Pronotum often with deep longitudinal furrow along midline. Second maxillary palpomere longer than third (Fig. 13D) | 9 (voeltzkowi group) |
| 8 | First flagellomere segment of antenna distinctly shorter than pedicel (as in Fig. 10C). Genal tooth relatively short, distinctly failing to reach to anterior margin of lateral lobe of clypeus (Fig. 14A). No carina present on dorsal edge of metapleural gland bulla. Body size relatively small (HW<1.50 mm). Petiole relatively wider in dorsal view (PtI>195). Body yellowish brown to reddish brown | Mystrium barrybressleri sp. n. (worker) |
| – | First flagellomere as long as pedicel (as in Fig 10D). Genal tooth relatively long, almost reaching anterior margin of lateral lobe of clypeus (Fig. 14B). Distinct carina present on dorsal edge of metapleural gland bulla. Body size relatively large (HW>1.70 mm). Petiole relatively narrower in dorsal view (PtI<185). Body reddish brown to black | Mystrium labyrinth sp. n. (worker) |
| 9 | Apical tooth strongly sclerotized, its apex darker than basal portion, strongly curving inward (Fig. 15A) | 10 (voeltzkowi group, worker) |
| – | Apical tooth moderately sclerotized, its apex same color as basal portion, straight and directed ventrally (Fig. 15B) | 15 (voeltzkowi group, ergatoid queen) |
| 10 | With head in posterior view, posterior face of vertex forming right or even acute angle with its dorsal face on median line, on lateral portion of vertex as steep as median portion (Fig. 16A) | 11 |
| – | With head in posterior view, posterior face of vertex forming blunt angle with its dorsal face on median line, lateral portion of vertex distinctly steeper than median portion (Fig. 16B) | 13 |
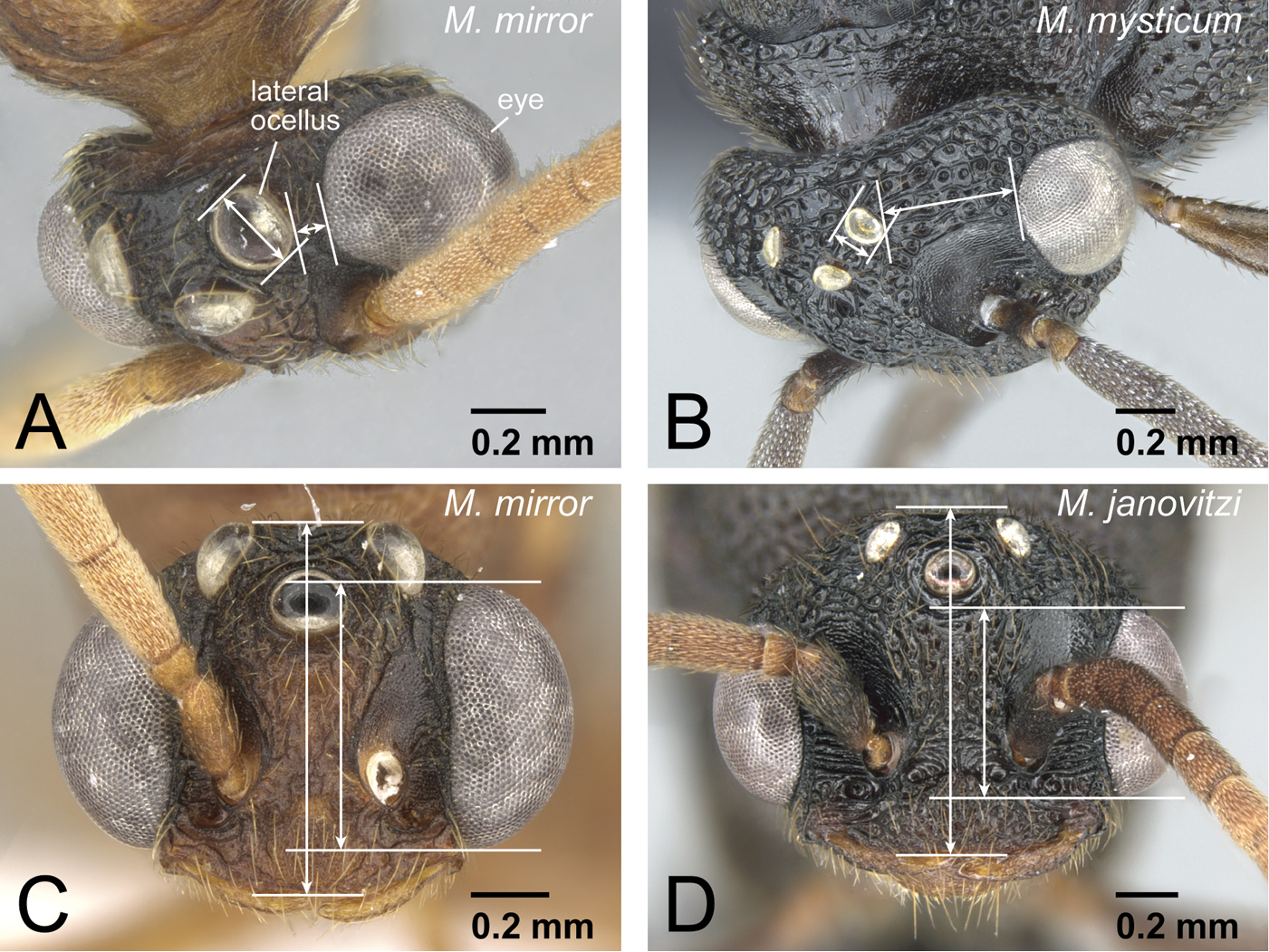
| 11 | Dorsal surface of pronotum without strong, deep, and thick longitudinal striae (Fig. 17A). Genal tooth undeveloped, distinctly fails to reach anterior margin of lateral lobe of clypeus in full-face view (Fig. 17C). Clypeal conical setae extremely small and short (Fig. 17C). Metanotal groove indistinct in lateral view. Mesonotum often not differentiated from propodeum in dorsal view | Mystrium janovitzi sp. n. (worker) |
| – | Dorsal surface of pronotum with strong, deep, and thick longitudinal striae (Fig. 17B). Genal tooth distinctly developed, almost reach to anterior margin of lateral lobe of clypeus in full-face view (Fig. 17D). Clypeal conical setae long (Fig. 17D). Metanotal groove shallow but distinct in lateral view. Mesonotum always clearly differentiated from propodeum in dorsal view | 12 |
| 12 | Anterior margin of clypeus straight (Fig. 18A). First flagellomere longer than pedicel (Fig. 18C). With mandible in dorsal view, width of visible surface on distal portion almost the same as on mandibular shaft (Fig. 18E) | Mystrium oberthueri Forel (worker) |
| – | Anterior margin of clypeus strongly convex (Fig. 18B). First flagellomere as long as pedicel (Fig. 18D). With mandible in dorsal view, visible surface on distal portion slightly wider than on mandibular shaft (Fig. 18F) | Mystrium eques sp. n. (worker) |
| 13 | Anterior margin of clypeus strongly convex with long conical setae, of which median pair usually larger than adjacent pair (Fig. 19A) | Mystrium shadow sp. n. (worker) |
| – | Anterior margin of clypeus almost straight, with short conical setae, of which median pair usually smaller than adjacent pair (Fig. 19B) | 14 |
| 14 | Genal tooth relatively short, reaching or slightly exceeding basal line of lateral clypeal lobe (Fig. 20A). Eye relatively larger than in individuals of Mystrium voeltzkowi of same body size (Fig. 20C). Metapleural gland bulla weakly developed, posterior margin of propodeal declivity almost straight (Fig. 20E) | Mystrium mirror sp. n. (worker) |
| – | Genal tooth relatively long, reaching to about half of lateral clypeal lobe (Fig. 20B). Eye relatively smaller than Mystrium mirror in individuals of same body size (Fig. 20D). Metapleural gland bulla strongly developed, posterior margin of propodeal declivity distinctly convex posteriorly on its ventral side (Fig. 20F) | Mystrium voeltzkowi Forel (worker) |
| 15 | With head in posterior view, posterior face of vertex forming right or even acute angle with its dorsal face on median line, lateral portion of vertex as steep as on median portion (Fig. 21A) | 16 |
| – | With head in posterior view, posterior face of vertex forming blunt angle with its dorsal face on median line, lateral portion of vertex distinctly steeper than median portion (Fig. 21B) | 18 |
| 16 | Anterior margin of clypeus strongly convex (Fig. 22A) | Mystrium eques sp. n. (ergatoid queen) |
| – | Anterior margin of clypeus almost straight (Fig. 22B) | 17 |
| 17 | Clypeal conical setae extremely short (Fig. 23A). First flagellomere as long as pedicel (Fig. 23C) | Mystrium janovitzi sp. n. (ergatoid queen) |
| – | Clypeal conical setae moderately long (Fig. 23B). First flagellomere longer than pedicel (Fig. 23D) | Mystrium oberthueri Forel (ergatoid queen) |
| 18 | Metapleural gland bulla extremely well developed, expanding above propodeal spiracle (Fig. 24A). Propodeal declivity in lateral view strongly convex posteriorly on its ventral two thirds | Mystrium voeltzkowi Forel (ergatoid queen) |
| – | Metapleural gland bulla weakly to moderately developed, not expanding above propodeal spiracle (Fig. 24B). Propodeal declivity in lateral view usually straight or convex posteriorly on its ventral third | 19 |
| 19 | Setae on pronotal dorsum almost simple, narrowing distally, with sharp apex (Fig. 25A). Anterolateral corner of head usually without spine, but sometimes with small spine. Vestigial wings often unclear and unrecognizable, or existing as quite small appendage (Fig. 25C) | Mystrium mirror sp. n. (ergatoid queen) |
| – | Setae on pronotal dorsum spatulate, widened distally and with sharp apex (Fig. 25B). Anterolateral corner of head with small spine. Vestigial wings clearly recognizable as appendages (Fig. 25D) | Mystrium shadow sp. n. (ergatoid queen) |

Female castes of Mystrium mysticum. A, C alate queen (CASENT0429962) B worker (CASENT0429955) D intercaste (CASENT0429959). A, B mesosoma in dorsal view C, D head in full-face view.

Female diagnostic characters of mysticum species group. A Mystrium rogeri (CASENT0001069) B Mystrium barrybressleri (CASENT0045009) C Mystrium mysticum (CASENT0003808) D Mystrium mirror (CASENT0317582). A–D worker. A, B mandible in oblique anterior view C, D abdominal segment VII in lateral view.

Queens of mysticum species group. A, C Mystrium mysticum (CASENT0429983) B, D Mystrium rogeri (CASENT0001070). A, B head to mesonotum in oblique lateral view C, D clypeus in oblique full-face view.

Queens of camillae species group. A, C Mystrium barrybressleri (CASENT0145109) B, D Mystrium labyrinth (CASENT0084950). A, B petiole and abdominal segment III in dorsal view C, D pedicel and flagellum in lateral view.

Vertex of Mystrium workers. A Mystrium mysticum (CASENT0429965) B Mystrium voeltzkowi (CASENT0002075).

Mandible of females of mysticum species group. A Mystrium mysticum (CASENT0429965) B Mystrium rogeri (CASENT0001069).

Worker diagnostic characters of camillae species group. A Mystrium barrybressleri (CASENT0045009) B Mystrium voeltzkowi (CASENT0002075) C Mystrium barrybressleri (CASENT0047970) D Mystrium voeltzkowi (CASENT0317584). A–D worker. A, B pronotum in dorsal view C, D mouthparts in oblique lateral view.
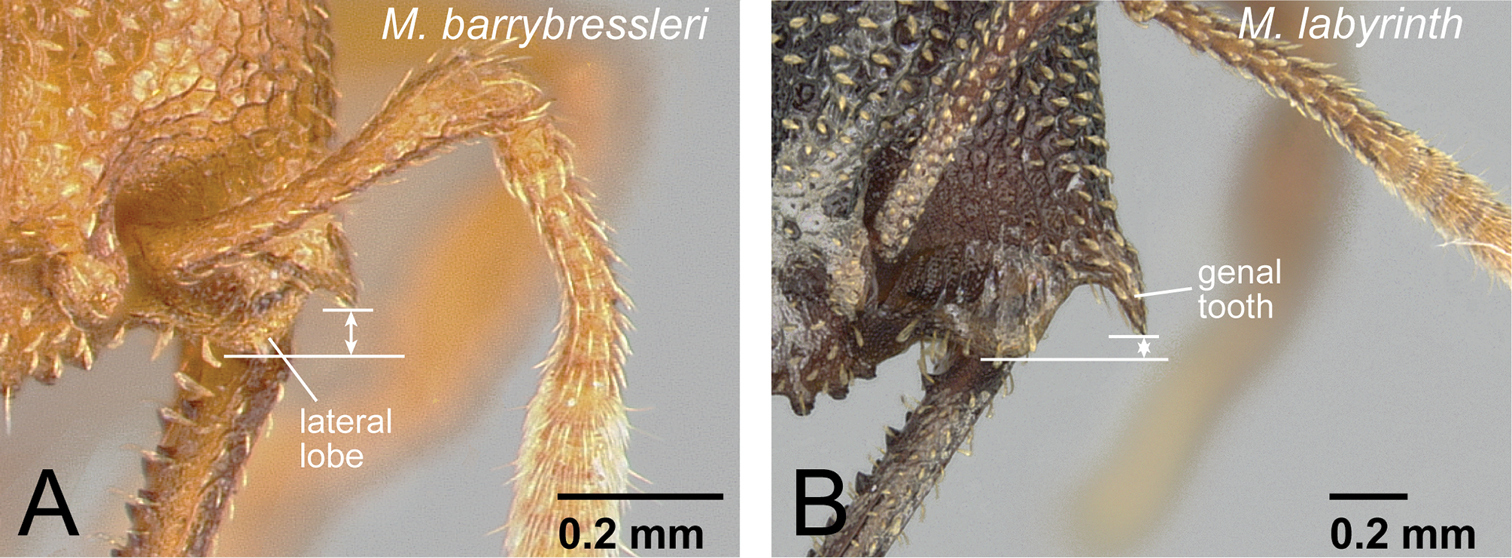
Head of workers in camillae species group. A Mystrium barrybressleri (CASENT0006081) B Mystrium labyrinth (CASENT0003281).

Distal portion of the mandible of Mystrium voeltzkowi in mesal view. A worker (CASENT0002075) B ergatoid queen (CASENT0002074).

Vertex of workers in voeltzkowi species group. A Mystrium oberthueri (CASENT0499569) B Mystrium voeltzkowi (CASENT0002075). A, B head in posterior view.

Differences in worker morphology between Mystrium janovitzi and Mystrium oberthueri. A Mystrium janovitzi (CASENT0006082) B Mystrium oberthueri (CASENT0499569) C Mystrium janovitzi (CASENT0482698: holotype) D Mystrium oberthueri (CASENT0496066). A, B mesosoma in dorsal view C, D clypeus and genal tooth in full-face view.

Differences in worker morphology between Mystrium oberthueri and Mystrium eques. A Mystrium oberthueri (CASENT0496066) B Mystrium eques (CASENT0418314: holotype) C, E Mystrium oberthueri (CASENT0499569) D, F Mystrium eques (CASENT0317399). A, B head in full-face view C, D pedicel and flagellum in lateral view E, F mandible in dorsal view.
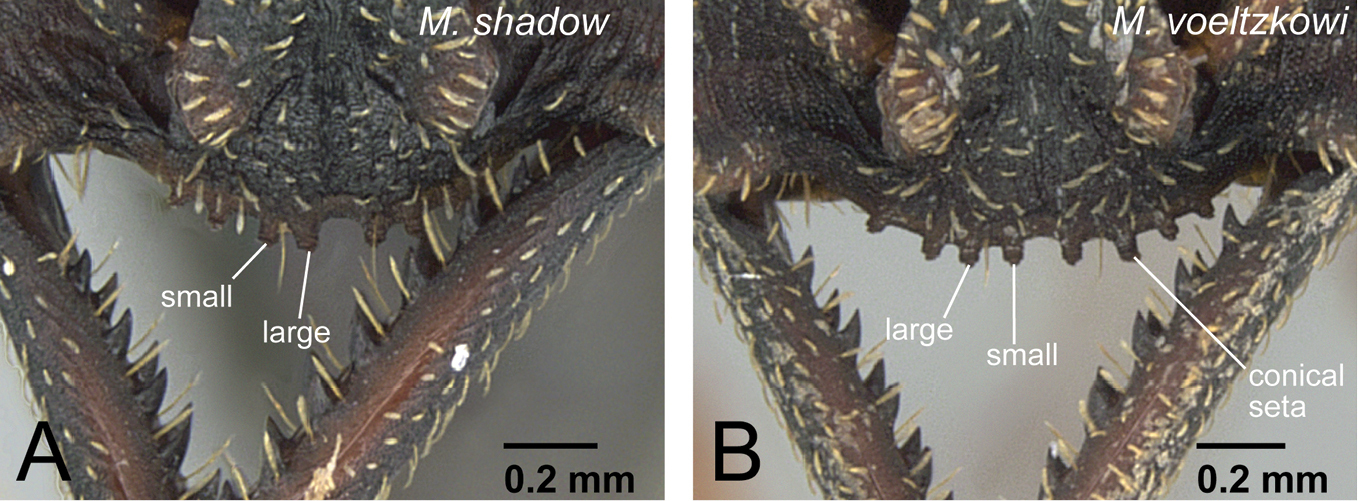
Clypeus of workers in voeltzkowi species group. A Mystrium shadow (CASENT0318933: holotype) B Mystrium voeltzkowi (CASENT0002075).
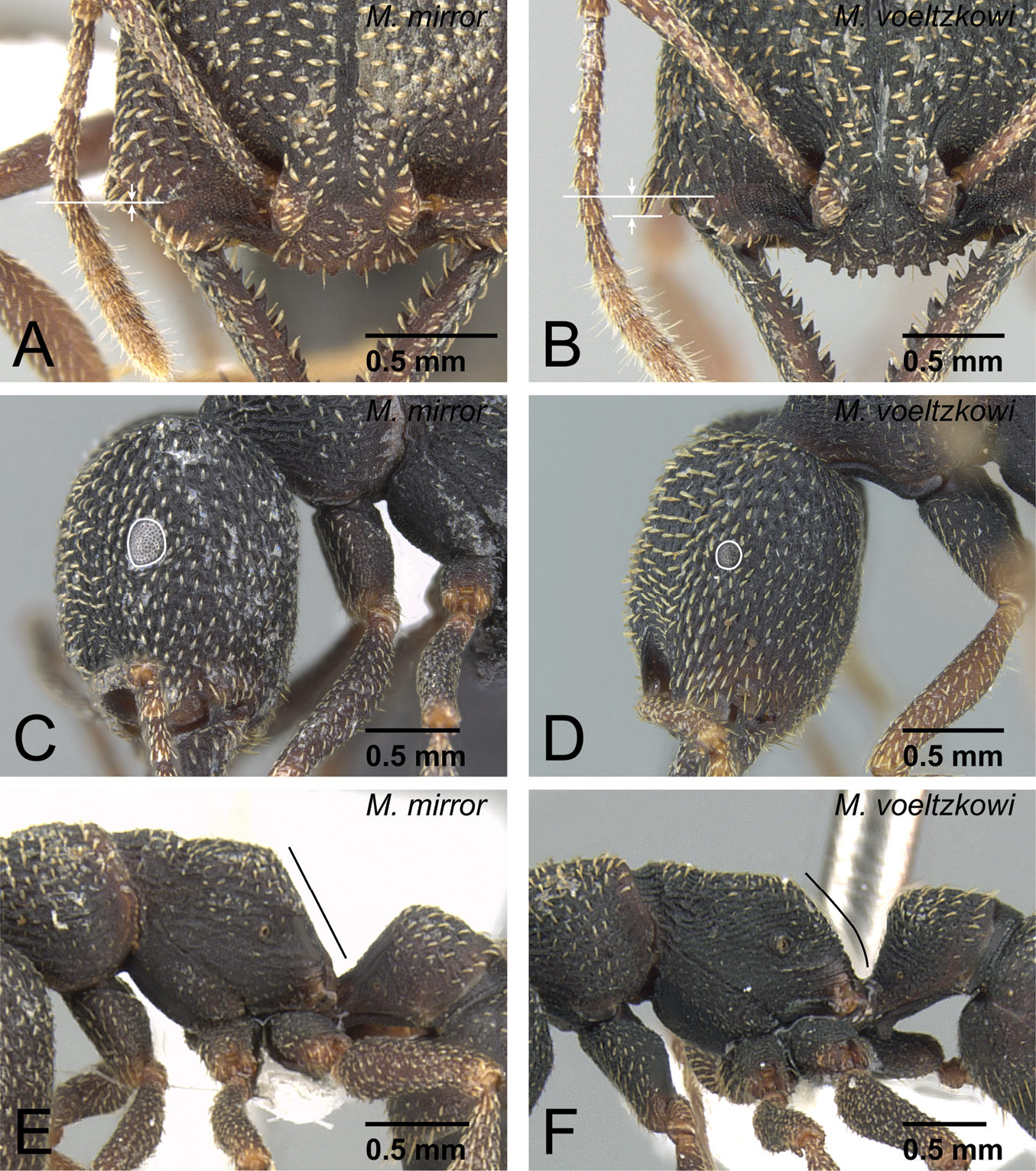
Differences in worker morphology between Mystrium mirror and Mystrium voeltzkowi. A, E Mystrium mirror (CASENT0429897) B, D, F Mystrium voeltzkowi (CASENT0002075) C Mystrium mirror (CASENT0002340). A, B head in full-face view C, D head in lateral view E, F mesosoma and petiole in lateral view.

Vertex of ergatoid queens in voeltzkowi species group. A Mystrium eques (CASENT0418311) B Mystrium shadow (CASENT0077647: paratype). A, B head in posterior view.

Clypeus of ergatoid queens in voeltzkowi species group. A Mystrium eques (CASENT0418317: paratype) B Mystrium oberthueri (CASENT0496064). A, B head in full-face view.

Differences in ergatoid queen characters between Mystrium janovitzi and Mystrium oberthueri. A Mystrium janovitzi (CASENT0482696) B Mystrium oberthueri (CASENT0496064) C Mystrium janovitzi (CASENT0003302) D Mystrium oberthueri (CASENT0095743). A, B head in full-face view C, D pedicel and flagellum in lateral view.

Propodeum of ergatoid queens of Mystrium voeltzkowi and Mystrium mirror in lateral view. A Mystrium voeltzkowi (CASENT0002092) B Mystrium mirror (CASENT0318940: paratype).
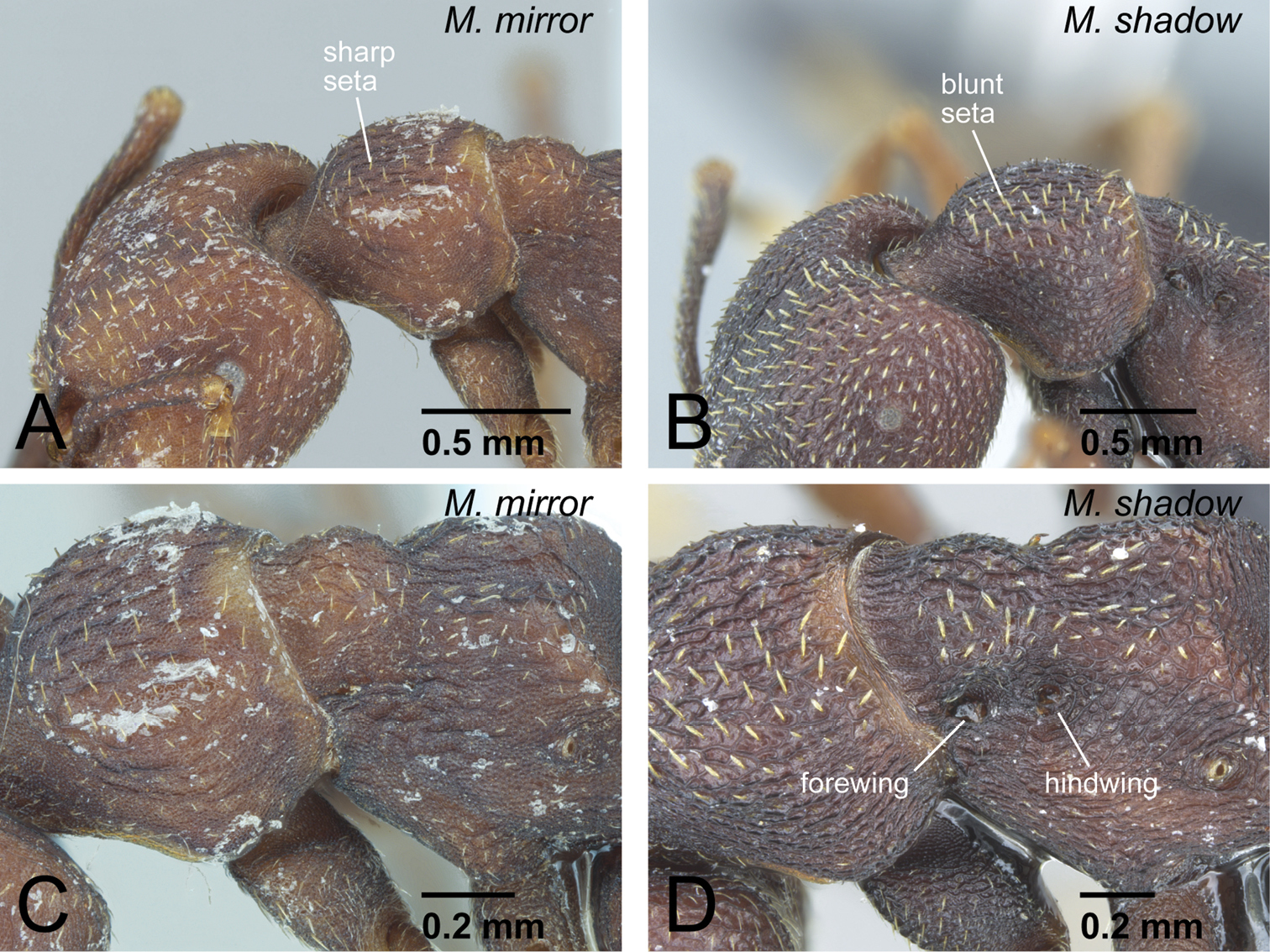
Differences in ergatoid queen characters between Mystrium mirror and Mystrium shadow. A, C Mystrium mirror (CASENT0429899) B, D Mystrium shadow (CASENT0077647: paratype). A, B head and pronotum in oblique dorsal view C, D mesosoma in oblique dorsal view.
Males of Mystrium labyrinth and Mystrium eques are unknown.
| 1 | Lateral ocellus relatively large and close to eye; distance between lateral ocellus and eye less than 1.2× maximum diameter of lateral ocellus (Fig. 26A). Eye large, occupying about 0.75× of head length (Fig. 26C). Body relatively brighter | 2 |
| – | Lateral ocellus relatively small and distant from eye; distance between lateral ocellus and eye more than 1.2× maximum diameter of lateral ocellus (Fig. 26B). Eye relatively smaller, occupying 0.5-0.6× of head length (Fig. 26D). Body relatively darker | 3 |
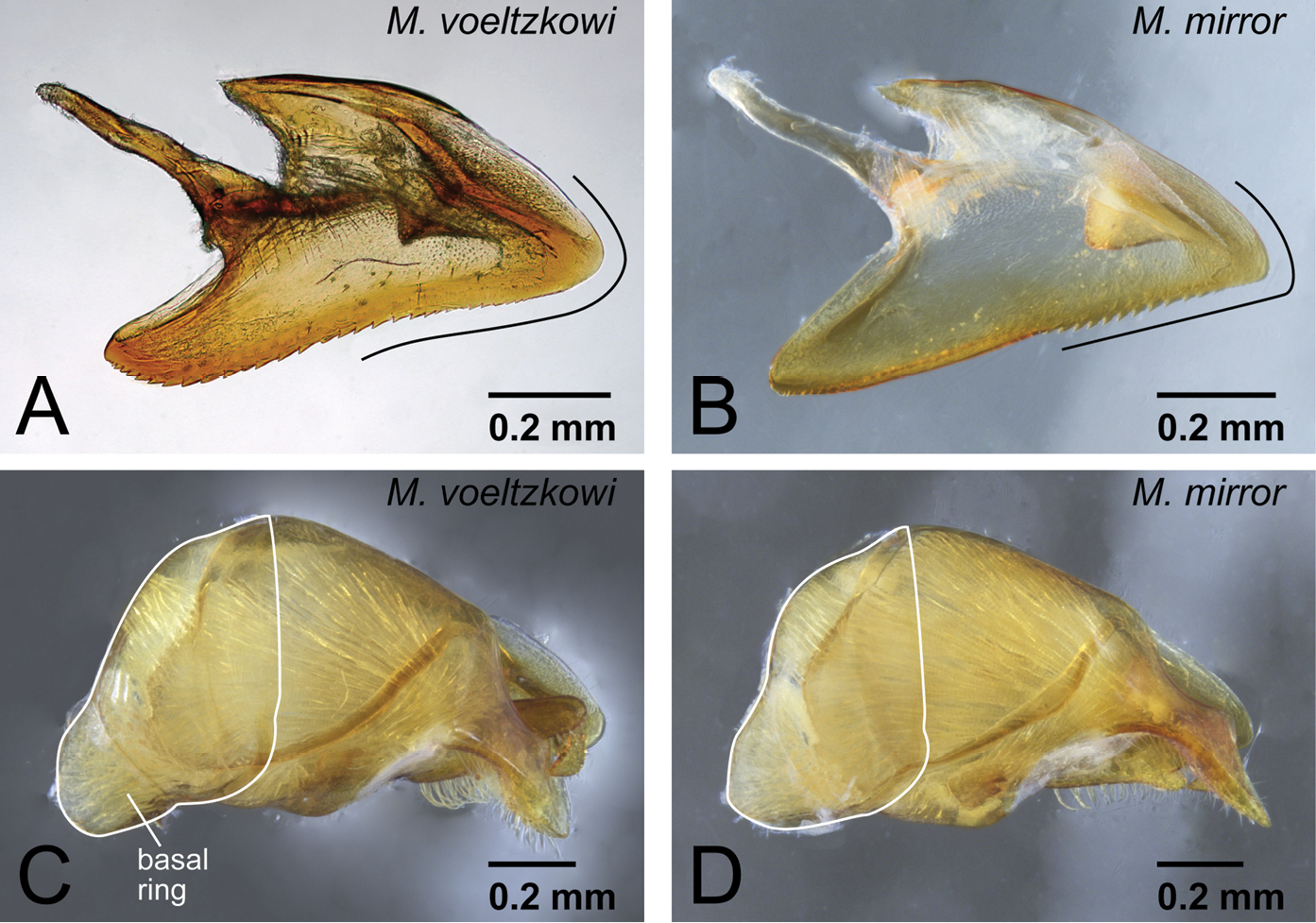
| 2 | Distal portion of aedeagus rounded (Fig. 27A). Basal ring gently extending basally (Fig. 27C) | Mystrium voeltzkowi Forel |
| – | Distal portion of aedeagus relatively sharp (Fig. 27B). Basal ring not extending basally (Fig. 27D) | Mystrium mirror sp. n. |
| 3 | Abdominal tergum VIII punctured (Fig. 28A). Petiolar dorsum covered with rough, deep punctures (Fig. 28C). Eye relatively small, occupying about half of head | 4 |
| – | Abdominal tergum VIII smooth and not punctured (Fig. 28B). Petiolar dorsum smooth, or covered with fine punctures, or at most shallow, irregular punctures (Fig. 28D). Eye relatively large, occupying about 0.6× of head length | 5 |
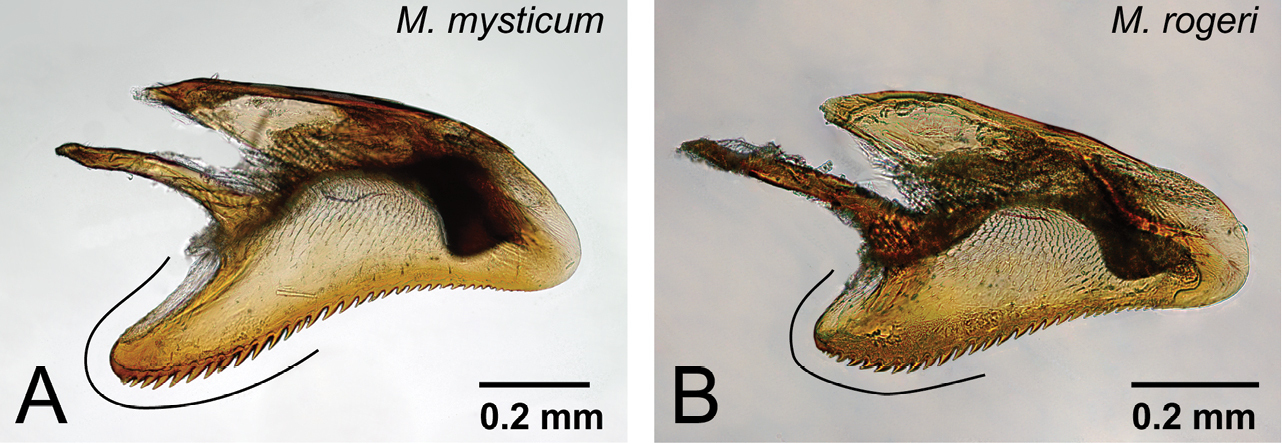
| 4 | Basoventral expansion of aedeagus long, its distal portion relatively narrow and sharp (Fig. 29A) | Mystrium mysticum Roger |
| – | Basoventral expansion of aedeagus short, its distal portion relatively wide and rounded (Fig. 29B) | Mystrium rogeri Forel |
| 5 | With head in full-face view, dorsal margin of head straight (Fig. 30A) | Mystrium oberthueri Forel |
| – | With head in full-face view, dorsal margin of head somewhat rounded (Fig. 30B) | 6 |
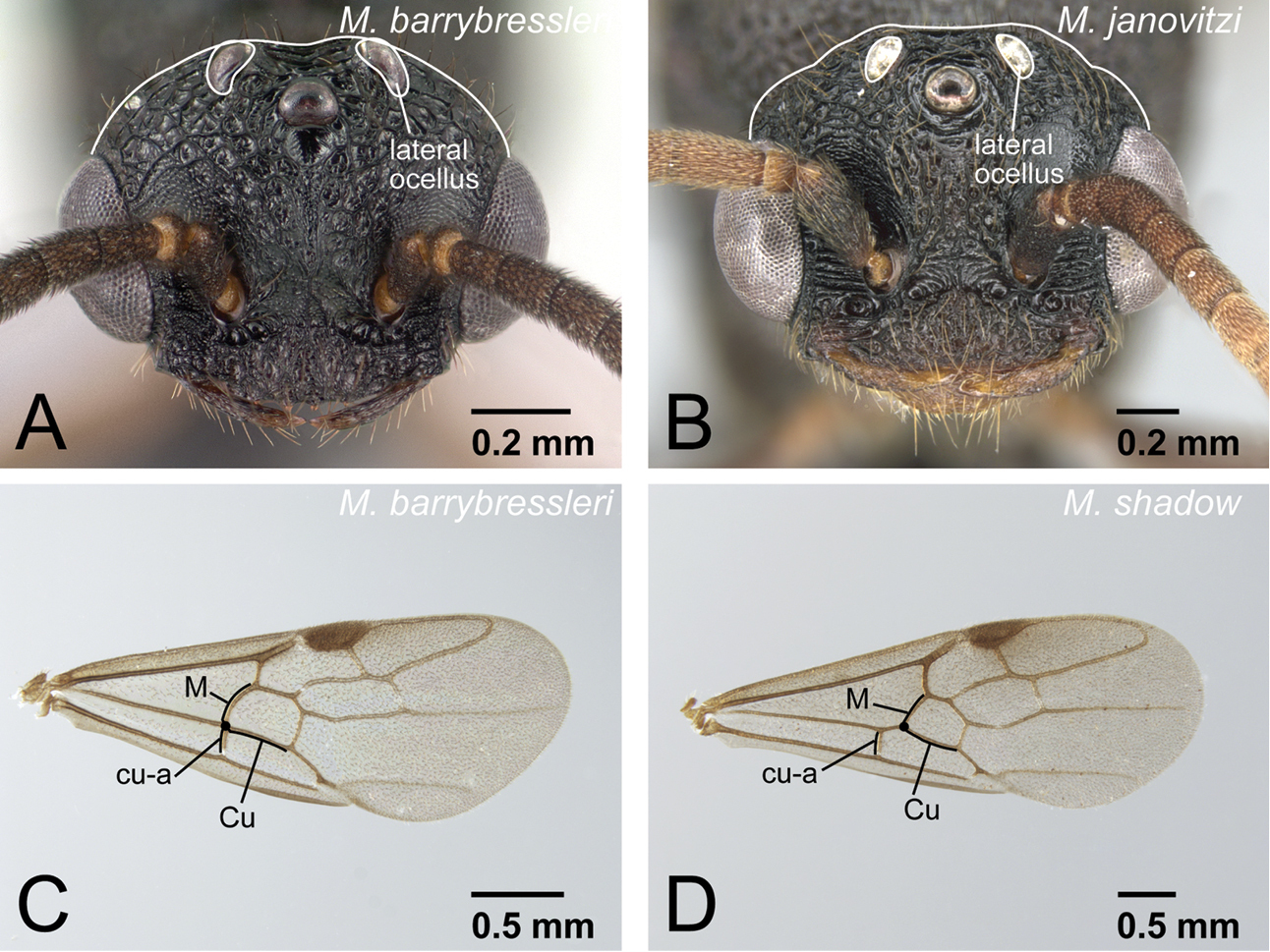
| 6 | Body size smaller (HW<1.20 mm). With head in full-face view, lateral ocelli clearly protruding from dorsal margin of head (Fig. 31A). On forewing, cu-a usually located at junction of Media (M) and Cubitus (Cu), sometimes slightly basal from junction (Fig. 31C). Petiole in dorsal view shorter, its length 0.5× of that of abdominal tergite III | Mystrium barrybressleri sp. n. |
| – | Body size larger (HW>1.50 mm). With head in full-face view, lateral ocelli reaching or failing to reach dorsal margin of head (Fig. 31B). On forewing, cu-a located far basal from junction of Media (M) and Cubitus (Cu) (Fig. 31D). Petiole in dorsal view longer, its length 0.6× or more of that of abdominal tergite III | 7 |
| 7 | Posterior face of vertex gently rising from neck, posterior and dorsal faces almost continuous without division (Fig. 32A). Basoventral expansion of aedeagus relatively short (Fig. 32C) | Mystrium shadow sp. n. |
| – | Posterior face of vertex steeply rising from neck, posterior and dorsal faces clearly divided (Fig. 32B). Basoventral expansion of aedeagus relatively long (Fig. 32D) | Mystrium janovitzi sp. n. |
Head of Mystrium males. A, C Mystrium mirror (CASENT0318941) B Mystrium mysticum (CASENT0429983) D Mystrium janovitzi (CASENT0007820). A, B in oblique dorsal view C, D full-face view.
Differences in male characters between Mystrium voeltzkowi and Mystrium mirror. A, C Mystrium voeltzkowi (CASENT0317583) B, D Mystrium mirror (CASENT0317444). A, B aedeagus in lateral view C, D genital capsule in lateral view.

Abdomen of Mystrium males. A Mystrium mysticum (CASENT0318027) B Mystrium janovitzi (CASENT0007820) C Mystrium mysticum (CASENT0429983) D Mystrium shadow (CASENT0303561). A, B abdominal segment VIII in oblique lateral view C, D petiole in dorsal view.
Differences in male genital characters between Mystrium mysticum and Mystrium rogeri. A Mystrium mysticum (CASENT0218102) B Mystrium rogeri (CASENT0001152). A, B aedeagus in lateral view.

Head of Mystrium males in full-face view. A Mystrium oberthueri (CASENT0179499) B Mystrium shadow (CASENT0107647).
Mystrium males. A Mystrium barrybressleri (CASENT0135866) B Mystrium janovitzi (CASENT0007820) C Mystrium barrybressleri (CASENT0078803) D Mystrium shadow (CASENT0107674). A, B head in full-face view C, D right forewing in dorsal view.
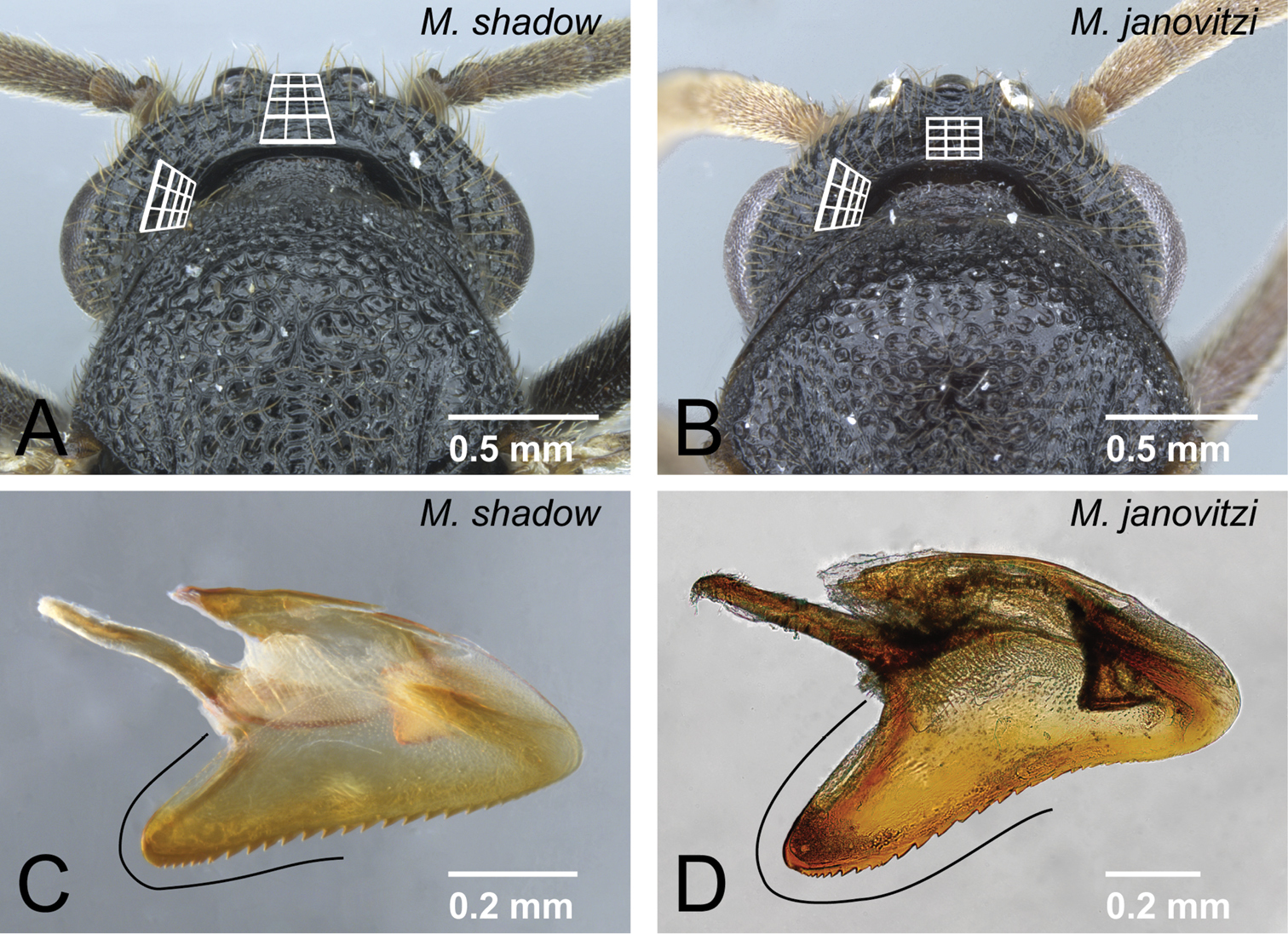
Differences in male characters between Mystrium shadow and Mystrium janovitzi. A Mystrium shadow (CASENT0303561) B Mystrium janovitzi (CASENT0080292) C Mystrium shadow (CASENT0107647) D Mystrium janovitzi (CASENT0080644). A, B head in posterior view C, D aedeagus in lateral view.
Due to the remarkable intra-specific morphological variation in each species, most specific characters are distinct only when individuals belonging to the same caste and in the same body size range are compared. The definition of each caste is given in each species-group description. Characters unique to each species group in the Malagasy region are given in italics.
The camillae species group has two species in the Malagasy region, Mystrium labyrinth sp. n. and Mystrium barrybressleri sp. n., in addition to the species known from outside of the Malagasy region: Mystrium camillae Emery, 1889, Mystrium leonie Bihn & Verhaagh, 2007, Mystrium maren Bihn & Verhaagh, 2007 from Asia and Mystrium silvestrii Santschi, 1914 from Africa. In addition to the four described extralimital species, we recognize two more undescribed morphospecies, one from Indonesia (Mystrium id01: CASENT0102169) and one from Mozambique (Mystrium mz01: CASENT0318957) in the species group. In this study we include minimal discussion of the species outside the Malagasy region because the material currently available is insufficient for an in-depth analysis. The camillae species group is based on the distinctly reticulate head and pronotal dorsum (Fig. 13A). Although several exceptions have been observed outside of the Malagasy region, the second maxillary palpomere, which is shorter than the third, differentiates this species group from the other Mystrium species in the Malagasy region (Fig. 13C). All known queens have wings, developed eyes and complete ocelli as is usual among ant queens. Rarely, the wings are shrunken and incomplete, as observed in an individual of Mystrium camillae collected in Indonesia (CASENT0009854), but the wing sclerites, e.g. the division of mesoscutum, mesoscutellum, and metanotum, are still fully developed (as in Fig. 7A).
Two phenotypes were observed in conspecific workers of Mystrium barrybressleri, Mystrium camillae, and Mystrium mz01 from Mozambique (Table 1): the mandible is stouter and the apical tooth is feebly sclerotized and curved inward in one phenotype (as in Fig. 15A), while the mandible is narrower and the apical tooth is moderately sclerotized and directed ventrally in the other (as in Fig. 15B). The differences between these two worker phenotypes in terms of mandible shape, body size, setae, and body color, are similar to those between workers and ergatoid queens in the voeltzkowi species group (see the voeltzkowi group description below for detail); however, these differences are relatively less pronounced. The small workers are also lack the wing buds, functional spermatheca, and developed ovaries, which were found in ergatoid queens in the voeltzkowi species group (
Although Mystrium camillae is not found in the Malagasy region, we designate the lectotype for Mystrium camillae and give some remarks below to discuss the diagnostic characters of Mystrium barrybressleri more clearly.
http://zoobank.org/0FFDA298-4426-4C05-88F2-A8712FE01526
http://species-id.net/wiki/Mystrium_barrybressleri
Figs 8B, 10A, 10C, 13A, 13C, 14A, 31A, 31C, 33A–C, 34A–C, 35A–B, 36A, 36C, 36E, 37A, 38A, 39A, 40A, 41A, 42AWorker: CASENT0129838, BLF15450, MADAGASCAR, Toliara, Forêt Ivohibe 55.0 km N Tolagnaro (-24.569°, 47.204°), 200 m alt., 2–4.xii.2006, B.L.Fisher et al. leg. [CASC].
2 workers: CASENT0129839 [BMNH], CASENT0129840 [MHNG], same data as holotype.
Description. Measurements: holotype. HL 1.26, HW 1.14, SL 0.69, ML 1.08, HD 0.72, WL 1.18, PnW 0.63, PpW 0.50, PtW 0.56, PtL 0.28, CI 90.5, SI 60.7, MI 94.5, PpI 79.7, PtI 198.2.
HL 0.79–1.52, HW 0.71–1.49, SL 0.46–0.89, ML 0.67–1.41, HD 0.47–0.91, WL 0.85–1.51, PnW 0.46–0.78, PpW 0.36–0.60, PtW 0.39–0.68, PtL 0.19–0.30, CI 89.7–100.4, SI 56.0–69.9, MI 86.4–97.5, PpI 77.1–87.5, PtI 199.0–227.8 (10 specimens measured).
Posterolateral corner of head strongly to moderately expanding posteriorly. Posterior face of vertex forming slightly blunt angle with dorsal face on median line of head, so that declivity of vertex on lateral part slightly steeper than on median part. Ventral half of vertex sculptured. Eye small, only with several ommatidia. Anterior margin of clypeus convex with long, conical setae, of which median pair larger than adjacent pair. Genal tooth of head moderately developed, reaching about half of lateral lobe of clypeus. Masticatory margin of mandible slightly visible in full-face view, and dorsal surface on distal portion of mandible slightly wider than that on mandibular shaft. Second maxillary palpomere shorter than third. First flagellomere (third antennal segment) about 0.5× length of pedicel (second antennal segment). Central part of pronotal dorsum strongly to weakly, and regularly reticulate. Lateral surface of pronotum strongly and regularly reticulate on its posterior portion. Mesonotum distinct in dorsal view, its length slightly shorter than that of propodeum. Metanotal groove shallowly impressed and mesonotum higher than pronotum in lateral view. No carina present on dorsal edge of metapleural gland bulla. Petiole extremely wide in dorsal view (PtI>195).
Body color yellowish brown to reddish brown.
Description. Measurements: HL 1.43, HW 1.52, SL 0.88, ML 1.10, HD 0.99, WL 2.05, MnW 1.23, PtW 1.12, PtL 0.46, CI 106.8, SI 58.0, MI 72.1, MnI 80.8, PtI 243.1 (one specimen measured).
Wings present and well developed. Wing sclerites fully developed even if wings have been removed. Posterolateral corner of head moderately expanding posteriorly; expansion weaker than that of workers. Posterior surface of vertex forming slightly blunt angle with its dorsal surface on median line of head, so that declivity of vertex on lateral part slightly steeper than on median part. Ventral half of vertex weakly sculptured, almost smooth. Eye well developed. Both anterior and lateral ocelli clearly present, median portion of lateral ocelli and posterior portion of anterior ocellus edged by blackish pigment. Anterior margin of clypeus convex with long conical setae, of which median pair larger than adjacent pair. Genal tooth of head moderately developed, reaching about half of lateral lobe of clypeus. Masticatory margin of mandible visible in full-face view, and width of dorsal surface on distal portion slightly wider than that on mandibular shaft. Spoon-shaped seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna short, about 0.5× length of pedicel. Setae on pronotum almost simple, narrowing distally with strongly sharpened apex. Propodeal declivity in lateral view straight, making right angle with its dorsal margin. Petiole wide and short in dorsal view, 0.25× length of abdominal segment III.
Body color yellow to yellowish brown.
Description. Measurements: HL 0.72–0.86, HW 0.91–1.17, SL 0.16–0.22, EL 0.39–0.45, WL 1.28–1.78, MnW 0.81–1.22, CI 126.8–137.5, SI 16.7–18.9, EI 52.5–55.7, MnI 88.8–108.2 (5 specimens measured).
Eye moderately large, occupying 0.6× head length. Ocelli protruding from dorsal margin of head in full-face view. Dorsal margin of head in full-face view rounded. Both anterior and lateral ocelli small. Lateral ocellus small and distant from eye: distance between these more than 2× maximum diameter of lateral ocellus. Posterior half of vertex clearly differentiated from its dorsal half, its dorsal face distinctly shorter than its posterior face. Palpal formula 4, 3. First segment of maxillary palp flattened and distinctly wider than second segment. Second maxillary palpomere shorter than third. Notauli weakly impressed on mesoscutum, but often unclear or absent. Petiole in dorsal view thin, its length 0.5× that of abdominal tergite III. Petiolar dorsum covered with fine punctures. Abdominal tergum VIII without deep punctures, almost smooth.
Distal portion of abdominal sternum IX smooth and not punctured. Basal ring quite short, not extending basally. Telomere slightly extending distally more than digitus. Basoventral expansion of aedeagus well developed basoventrally, distinctly longer than dorsal extension. Ventral margin of aedeagus gently curved ventrally in lateral view. Aedeagus moderately narrowing distally on its distal half and its distal portion narrowly rounded.
On forewing, cu-a usually located at junction of Media (M) and Cubitus (Cu), sometimes slightly basal from junction.
Body color reddish brown to blackish brown.

Workers of camillae species group. A, C Mystrium barrybressleri sp. n. (CASENT0129838) B Mystrium barrybressleri sp. n. (CASENT0129840) D Mystrium camillae (CASENT0102123) E Mystrium labyrinth sp. n. (CASENT0003281). A, C major worker (holotype) B minor worker (paratype) D major worker (lectotype) E minor worker (holotype). A, B, E head to abdomen in lateral view C, D mesosoma in oblique dorsal view C the lateral portion of the propodeum is gently and weakly convex D the lateral portion of the propodeum is steeply and strongly convex.

Head of workers of camillae species group. A Mystrium barrybressleri sp. n. (CASENT0129838) B Mystrium barrybressleri sp. n. (CASENT0129840) C Mystrium barrybressleri sp. n. (CASENT0045009) D Mystrium camillae (CASENT0102123) E Mystrium labyrinth sp. n. (CASENT0003281). A major worker (holotype) B minor worker (paratype) C major worker D major worker (lectotype) E minor worker (holotype) A, B, E in full-face view C, D clypeus in oblique anterior view C a pair of spatulate setae on the anteromedial portion of clypeus is long D a pair of spatulate setae on the anteromedial portion of clypeus is short.

Workers of camillae species group in dorsal view. A Mystrium barrybressleri sp. n. (CASENT0129838) B Mystrium barrybressleri sp. n. (CASENT0129840) C Mystrium labyrinth sp. n. (CASENT0003281). A major worker (holotype) B minor worker (paratype) C minor worker (holotype).

Queens of camillae species group. A, C, E Mystrium barrybressleri sp. n. (CASENT0145109) B, D, F Mystrium labyrinth sp. n. (CASENT0084950). A, B head to abdomen in lateral view C, D head in full-face view E, F head to abdomen in dorsal view.

Males of Mystrium in lateral view. A Mystrium barrybressleri sp. n. (CASENT0135866) B Mystrium mysticum (CASENT0318027) C Mystrium rogeri (CASENT0001083) D Mystrium janovitzi sp. n. (CASENT0007820) E Mystrium mirror sp. n. (CASENT0318941) F Mystrium oberthueri (CASENT0179499) G Mystrium shadow sp. n. (CASENT0107647) H Mystrium voeltzkowi (CASENT0101673).

Head of Mystrium males in full-face view. A Mystrium barrybressleri sp. n. (CASENT0135866) B Mystrium mysticum (CASENT0318027) C Mystrium rogeri (CASENT0001083) D Mystrium janovitzi sp. n. (CASENT0007820) E Mystrium mirror sp. n. (CASENT0318941) F Mystrium oberthueri (CASENT0179499) G Mystrium shadow sp. n. (CASENT0107647) H Mystrium voeltzkowi (CASENT0101673).

Males of Mystrium in dorsal view. A Mystrium barrybressleri sp. n. (CASENT0135866) B Mystrium mysticum (CASENT0318027) C Mystrium rogeri (CASENT0001083) D Mystrium janovitzi sp. n. (CASENT0007820) E Mystrium mirror sp. n. (CASENT0318941) F Mystrium oberthueri (CASENT0179499) G Mystrium shadow sp. n. (CASENT0107647) H Mystrium voeltzkowi (CASENT0101673).
Genital capsule of Mystrium males in lateral view. A Mystrium barrybressleri sp. n. (CASENT0078803) B Mystrium mysticum (CASENT0218102) C Mystrium rogeri (CASENT0001152) D Mystrium janovitzi sp. n. (CASENT0080644) E Mystrium mirror sp. n. (CASENT0317444) F Mystrium oberthueri (CASENT0317422) G Mystrium shadow sp. n. (CASENT0107647) H Mystrium voeltzkowi (CASENT0317583).

Aedeagus of Mystrium males in lateral view. A Mystrium barrybressleri sp. n. (CASENT0078803) B Mystrium mysticum (CASENT0218102) C Mystrium rogeri (CASENT0001152) D Mystrium janovitzi sp. n. (CASENT0080644) E Mystrium mirror sp. n. (CASENT0317444) F Mystrium oberthueri (CASENT0317422) G Mystrium shadow sp. n. (CASENT0107647) H Mystrium voeltzkowi (CASENT0317583).
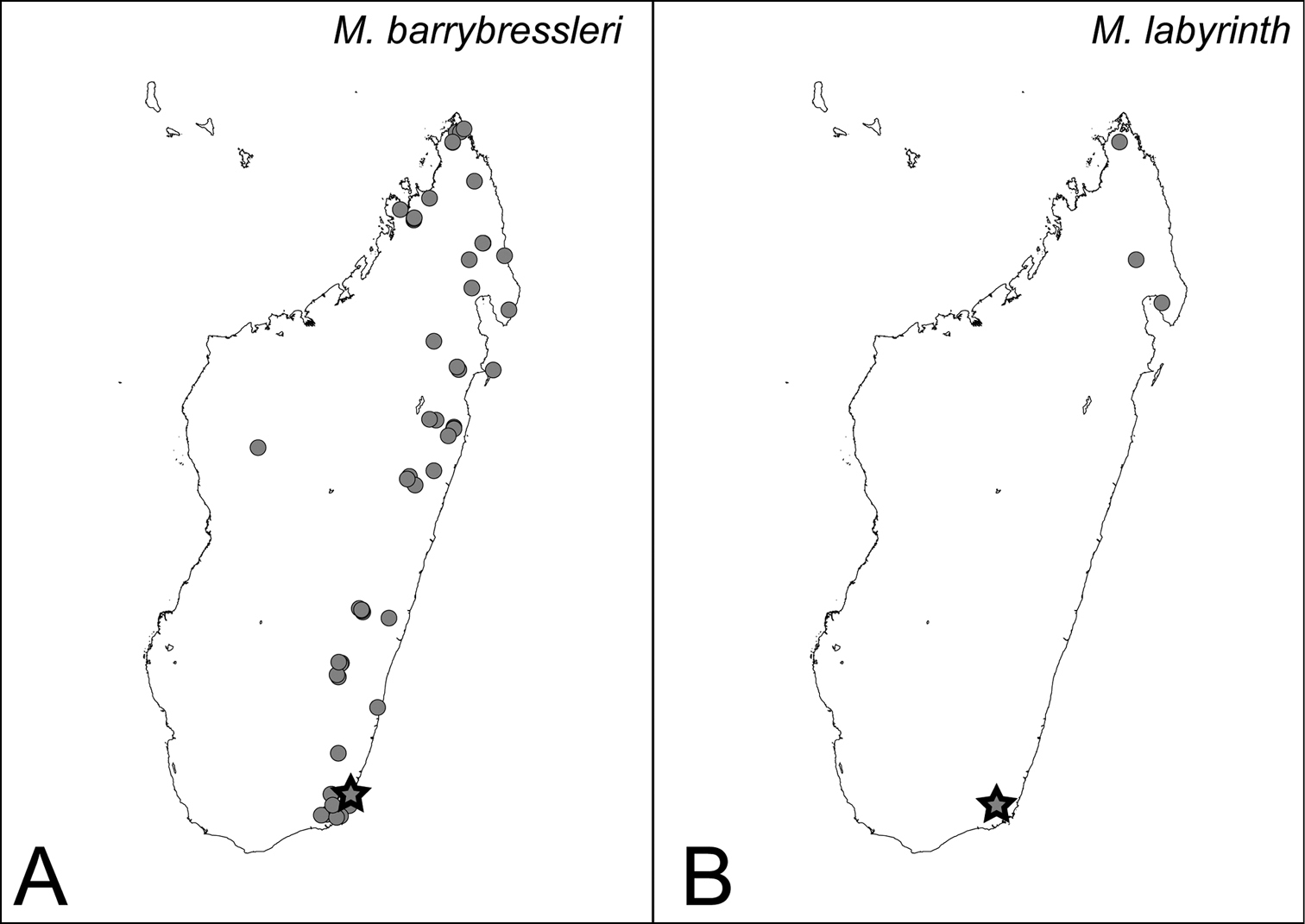
Distribution maps for camillae species group. A Mystrium barrybressleri sp. n. B Mystrium labyrinth sp. n. Star symbols represent the type locality.
The specific epithet is the patronym of Dr. Barry Lee Bressler, retired physicist, former adjunct professor of physics at Virginia Polytechnic Institute and State University, and amateur naturalist, in recognition of his interest in myrmecology and his support for research on ants.
MADAGASCAR: as in Figure 42A.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Diego-Suarez. Sakalava Beach [vegetated beach dunes] (-12.26278°, 49.3975°), across sandy trail in dwarf littoral forest, 10 m alt.; Montaigne Francais (-12.325°, 49.33333°), along forested limestone ridge, 150 m alt.; 7 km N Joffreville [camp 2 of Fisher] (-12.33333°, 49.25°), in dry forest, 360 m alt.; Parc National Montagne d’Ambre [1st campsite] (-12.51444°, 49.18139°), rainforest, 960 m alt.; [Petit Lac road] (-12.52028°, 49.17917°), rainforest, 1125 m alt.; Antsiranana. Forêt de Binara, 9.4 km 235° SW Daraina (-13.26333°, 49.6°), montane rainforest, 1100 m alt.; 9.1 km 233° SW Daraina (-13.26333°, 49.60333°), rainforest, 800 m alt.; Ampasindava, Forêt d’Ambilanivy, 3.9 km 181° S Ambaliha (-13.79861°, 48.16167°), rainforest, 600 m alt.; R.S. Manongarivo, 10.8 km 229° SW Antanambao (-13.96167°, 48.43333°), rainforest, 400 m alt.; Manongarivo, 12.8 km 228° SW Antanambao (-13.97667°, 48.42333°), rainforest, 780 m alt.; 14.5 km 220° SW Antanambao (-13.99833°, 48.42833°), montane rainforest, 1175 m alt.; Parc National de Marojejy, Manantenina River, 27.6 km 35° NE Andapa, 9.6 km 327° NNW Manantenina (-14.435°, 49.76°), rainforest, 775 m alt.; 28.0 km 38° NE Andapa, 8.2 km 333° NNW Manantenina (-14.43667°, 49.775°), rainforest, 450 m alt.; SAVA Region, district of Sambava, Marojejy National Park, 5 km W of Manantenina village, 1st campsite (Mantella) (-14.43817°, 49.774°), low altitude rainforest, 487 m alt.; Forêt Ambanitaza, 26.1 km 347° Antalaha (-14.67933°, 50.18367°), rainforest, 240 m alt.; 6.5 km SSW Befingotra, Rés. Anjanaharibe-Sud (-14.75°, 49.5°), rainforest, 875 m alt.; Fotodriana, Cap Masoala (-15.69694°, 50.27028°), lowland rainforest, 25 m alt.; Toamasina. Montagne d’Akirindro 7.6 km 341° NNW Ambinanitelo (-15.28833°, 49.54833°), rainforest, 600 m alt.; Réserve Spéciale Ambatovaky, Sandrangato river (-16.7633°, 49.26692°), rainforest, 520 m alt.; (-16.81753°, 49.29498°), rainforest, 360 m alt.; Ile Sainte Marie, Forêt Ambohidena, 22.8 km 44° Ambodifotatra (-16.82433°, 49.96417°), littoral rainforest, 20 m alt.; Parc National de Zahamena, Tetezambatana forest, near junction of Nosivola and Manakambahiny Rivers (-17.74298°, 48.72936°), rainforest, 860 m alt.; Onibe River (-17.75908°, 48.85468°), rainforest, 780 m alt.; Reserve Betampona, Camp Vohitsivalana, 37.1 km 338° Toamasina (-17.88667°, 49.2025°), rainforest, 520 m alt.; Réserve Naturelle Betampona, 34.1 km 332° Toamasina (-17.916135°, 49.20185°), rainforest, 550 m alt.; 34.08 km 332° Toamasina (-17.91977°, 49.20039°), rainforest, 525 m alt.; Camp Rendrirendry 34.1 km 332° Toamasina (-17.924°, 49.19967°), rainforest, 390 m alt.; F.C. Sandranantitra (-18.04833°, 49.09167°), rainforest, 450 m alt.; F.C. Andriantantely (-18.695°, 48.81333°), rainforest, 530 m alt.; Analamay (-18.80623°, 48.33707°), montane rainforest, 1068 m alt.; Ambatovy, 12.4 km NE Moramanga (-18.84773°, 48.29568°), montane rainforest, 1000 m alt.; Torotorofotsy (-18.87082°, 48.34737°), montane rainforest, marsh edge, 1070 m alt.; 7 km SE Andasibe National Park Headquarters (-18.96278°, 48.45267°), tropical forest, 1050 m alt.; Mahajanga. Réserve Spéciale Marotandrano, Marotandrano 48.3 km S Mandritsara (-16.28322°, 48.81443°), transition humid forest, 865 m alt.; Toliara. Réserve Spéciale d’Ambohijanahary, Forêt d’Ankazotsihitafototra, 35.2 km 312° NW Ambaravaranala (-18.26667°, 45.40667°), montane rainforest, 1050 m alt.; 11 km NW Enakara, Rés. Andohahela (-24.56667°, 46.83333°), rainforest, 800 m alt.; 10 km NW Enakara, Rés. Andohahela (-24.56667°, 46.81667°), rainforest, 430 m alt.; Parc National Andohahela, Col de Tanatana, 33.3 km NW Tolagnaro (-24.7585°, 46.85367°), rainforest, 275 m alt.; 2.7 km WNW 302° Ste. Luce (-24.77167°, 47.17167°), littoral rainforest, 20 m alt.; 29.5 km WNW Tolanaro, Vasiha Mt. (-24.92306°, 46.74083°), rainforest, 700 m alt.; Tsimelahy - Parcel II, Andohahela National Park, transition forest (-24.93683°, 46.62667°), transition forest, 180 m alt.; Mandena, 8.4 km NNE 30° Tolagnaro (-24.95167°, 47.00167°), littoral rainforest, 20 m alt.; Manatantely, 8.9 km NW Tolagnaro (-24.9815°, 46.92567°), rainforest, 100 m alt. Fianarantsoa. Vohiparara broken bridge, Fianarantsoa Prov. (-21.22617°, 47.36983°), high altitude rainforest, 1110 m alt.; 7 km W Ranomafana National Park (-21.25°, 47.41667°), montane rainforest, 900 m alt.; radio tower, Ranomafana National Park, Fianarantsoa Prov. (-21.25083°, 47.40717°), forest edge, mixed tropical forest, open area, 1130 m alt.; Belle Vue trail, Ranomafana National Park, Fianarantsoa Prov. (-21.2665°, 47.42017°), mixed tropical forest, 1020 m alt.; Parc National de Ranomafana, Vatoharanana River, 4.1 km 231° SW Ranomafana (-21.29°, 47.43333°), montane rainforest, 1100 m alt.; 7.6 km 122° Kianjavato, Forêt Classée Vatovavy (-21.4°, 47.94°), rainforest, 175 m alt.; 36 km S Ambalavao, Rés. Andringitra (-22.2°, 46.96667°), montane rainforest, 820 m alt.; 45 km S. Ambalavao (-22.21667°, 47.01667°), rainforest, 785 m alt.; 43 km S Ambalavao, Rés. Andringitra (-22.23333°, 47°), rainforest, 825 m alt.; 9.0 km NE Ivohibe (-22.42667°, 46.93833°), rainforest, 900 m alt.; R.S. Ivohibe, 7.5 km ENE Ivohibe (-22.47°, 46.96°), rainforest, 900 m alt.; Manombo Special Reserve, 32 km SE of Farafangana (-23.02183°, 47.72°), lowland rainforest, 36 m alt.; Parc National Befotaka-Midongy, Papango 27.7 km S Midongy-Sud, Mount Papango (-23.83517°, 46.96367°), rainforest, 940 m alt.
The workers of Mystrium barrybressleri can be distinguished from other Mystrium workers and queens in the Malagasy region by a combination of the following characters: mesosoma lacking wing-sclerites; first flagellomere (third segment of antenna) shorter than pedicel (second segment of antenna) (as in Fig. 10C); and second maxillary palpomere shorter than third (Fig. 13C). The wider petiole in dorsal view (Figs 35A, B) separates Mystrium barrybressleri workers from the other Mystrium workers in the Malagasy region. A combination of smaller body size, small lateral ocelli distant from the eye (as in Fig. 26B), rounded posterior margin of head with protruding lateral ocelli in full-face view (Fig. 31A), cu-a on the forewing located at junction of Media (M) and Cubitus (Cu) (Fig. 31C), wide and short petiole in dorsal view, and fine punctation on abdominal tergite III separates Mystrium barrybressleri males from other known Mystrium males in the Malagasy region.
The workers of Mystrium barrybressleri are most similar to those of Mystrium camillae Emery. Mystrium camillae appears to be a complex of several species (see discussion in Mystrium camillae) but we did not have sufficient material to fully evaluate the castes and worker variation throughout the range of the species complex. Mystrium barrybressleri workers can be distinguished by the shape of the propodeum. In workers, the lateral portion of the propodeum is steeply and strongly convex in Mystrium “camillae” (Fig. 33D), while the convexity is gentle and weak in Mystrium barrybressleri (Fig. 33C). Also, in Mystrium barrybressleri, minor workers have setae much narrower and simpler than those of major workers (Fig. 35A: major vs. 35B: minor), while in Mystrium “camillae” minor and major workers have similar setae.
There are additional differences with Mystrium camillae (sensu stricto, as defined by the syntype series from Myanmar). The pair of spatulate setae on the anteromedial portion of clypeus is distinctly longer in Mystrium barrybressleri (Fig. 34C) than those in the syntype series of Mystrium camillae (Fig. 34D). The queen of Mystrium barrybressleri can be distinguished by the presence of simple setae on the pronotum, while in Mystrium camillae sensu stricto, the setae are clavate. Additional diagnostic characters can be presented once the species limits are further refined for Mystrium camillae and related species.
Mystrium barrybressleri is found from the rainforests and montane rainforests, and the distribution pattern of this species is similar to that of a forest species Tetramorium andrei Forel (
http://species-id.net/wiki/Mystrium_camillae
Figs 33D, 34D[here designated]. Worker: CASENT0102123, MYANMAR, Bhamo, iv.1886, Fea leg. [MSNG: examined].
The lectotype for Mystrium camillae is designated here. We confirm that five workers collected from vi.1885 to iv.1886 are part of the syntype series: two workers in MSNG [CASENT0102123, CASENT0102124]; one worker in MNHN [CASENT0101450]; and two workers in MHNG [CASENT0101809, CASENT0101784]. Although
According to the revision of Mystrium in the Indo-Australian region (
We have found that clarifying species boundaries in the genus Mystrium is much more complicated than would be expected from previous studies. The differences among some sibling species similar to Mystrium camillae could be recognizable only in a particular sex, caste, or phenotype, as in the case of Mystrium camillae and Mystrium barrybressleri. A reexamination of the species boundaries of Mystrium camillae based on a detailed comparative study using comprehensive colony samples from each local region will be necessary.
http://zoobank.org/0E7BCDE0-AF81-4120-9FC0-C1287C4BED46
http://species-id.net/wiki/Mystrium_labyrinth
Figs 10B, 10D, 14B, 33E, 34E, 35C, 36B, 36D, 36F, 42BWorker: CASENT0003281, BLF00976, MADAGASCAR, Toliara, 5 km N Isaka-Invondro (-24.75°, 46.8°), 350 m alt., 12.xi.1992, B.L.Fisher leg. [CASC].
Description. Measurements: holotype. HL 1.73, HW 1.76, SL 1.08, ML 1.77, HD 1.03, WL 1.96, PnW 1.01, PpW 0.84, PtW 0.90, PtL 0.51, CI 101.7, SI 61.2, MI 100.5, PpI 83.0, PtI 176.1.
HL 1.67–1.79, HW 1.72–1.84, SL 1.05–1.09, ML 1.70–1.76, HD 1.03–1.09, WL 1.90–2.02, PnW 1.02–1.08, PpW 0.81–0.87, PtW 0.82–0.95, PtL 0.45–0.52, CI 102.0–102.8, SI 58.0–61.1, MI 92.2–99.4, PpI 79.0–82.7, PtI 180.4–183.8 (3 specimens measured).
Posterolateral corner of head moderately expanding posteriorly. Posterior face of vertex forming almost right angle with its dorsal face on median line of head, so that declivity of vertex on lateral part as steep as that on median part. Ventral half of vertex sculptured. Eye relatively larger than that of Mystrium barrybressleri. Anterior margin of clypeus convex with long conical setae, of which median pair larger than adjacent pair. Genal tooth of head relatively long, as long as lateral lobe of clypeus. Masticatory margin of mandible in full-face view slightly visible on its basal half, invisible on its distal half. Width of dorsal surface of mandible almost identical from mandibular shaft to distal portion. Second maxillary palpomere shorter than third. First flagellomere (third antennal segment) as long as pedicel (second antennal segment). Central part of pronotal dorsum and lateral surface of pronotum strongly and regularly reticulate. Mesonotum often not differentiated and indistinct from propodeum in dorsal view, its length as long as that of propodeum. Metanotal groove indistinct in lateral view, but mesonotum slightly higher than pronotum. Short, but distinct ridge present on dorsal edge of metapleural gland bulla. Petiole wide in dorsal view, but narrower than that of Mystrium barrybressleri (PtI<185).
Body color reddish brown to black.
Description. Measurements: HL 1.65, HW 1.69, SL 0.99, ML 1.48, HD 1.10, WL 2.33, MnW 1.27, PtW 1.08, PtL 0.51, CI 102.5, SI 58.5, MI 87.6, MnI 74.8, PtI 212.2 (one specimens measured).
Wings present, well developed. Wing sclerites fully developed even if wings have dropped off. Posterolateral corner of head moderately expanding posteriorly; expansion weaker than that of workers. Posterior face of vertex forming almost right angle with its dorsal face on median line of head, so that declivity of vertex on lateral part as steep as that on median part. Ventral half of vertex sculptured. Eye well developed. Both anterior and lateral ocelli clearly present, median portion of lateral ocelli and posterior portion of anterior ocellus edged by blackish pigment. Anterior margin of clypeus convex, with long conical setae, of which median pair larger than adjacent pair. Genal tooth of head distinctly developed, reaching slightly posterior of lateral lobe of clypeus. Masticatory margin of mandible almost invisible in full-face view, and dorsal surface on distal portion as wide as that on mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna as long as pedicel. Setae on pronotum distinctly spatulate, widened distally. Propodeal declivity in lateral view slightly convex on dorsal part of metanotal gland bulla, making blunt angle with its dorsal margin. Petiole relatively long in dorsal view, 0.5× length of abdominal segment III.
Body color blackish brown.
Male unknown.
This species name is the English word labyrinth, inspired by the strong reticulation covering the body surface of the new species. The species epithet is a noun, and thus invariant.
MADAGASCAR: as in Figure 42B.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Diego-Suarez. Parc National Montagne d’Ambre (-12.51444°, 49.18139°), rainforest, 960 m alt.; Antsiranana. 6.5 km SSW Befingotra, Rés. Anjanaharibe-Sud (-14.75°, 49.5°), rainforest, 875 m alt.; Toamasina. 6.9 km NE Ambanizana, Ambohitsitondroina (-15.56667°, 50°), rainforest, 825 m alt.; Toliara. 5 km N Isaka-Invondro (-24.75°, 46.8°), rainforest, 350 m alt.
Mystrium labyrinth females can be distinguished easily from the other Mystrium females in the Malagasy region by a combination of the following characters: the pronotal dorsum covered with strong regular reticulation (as in Fig. 13A); the second maxillary palpomere shorter than the third (as in Fig. 13C); the long genal tooth of head nearly reaching anterior end of the lateral lobe of the clypeus (Fig. 14B); and the first flagellomere (third antennal segment) as long as the pedicel (second antennal segment) (Fig. 10D). The worker of Mystrium labyrinth is similar to that of Mystrium silvestrii; however, Mystrium labyrinth can be distinguished from Mystrium silvestrii by an anterior clypeal margin with long, well-developed conical setae (weak in Mystrium silvestrii and Mystrium mz01), and a moderately long petiole (Fig. 35C) (short and wide in Mystrium silvestrii as in Mystrium barrybressleri).
The queen described here is not associated with workers from the same colony (collected alone from -12.51444°, 49.18139°); however, the similarities between workers and queens are sufficient to determine their relationship. The male of this species is unknown.
The mysticum species group is endemic to the Malagasy region, and consists of two species, Mystrium mysticum Roger, 1862 and Mystrium rogeri Forel, 1899. Unique characters in this group are: a longitudinal carina on the central portion of the labrum in worker and queen (Fig. 8A); a single lateral spine on abdominal sternum VII in worker and queen (Fig. 8C); and unsculptured ventral area on the vertex in worker (Fig. 11A). The separation of the queens is usually straightforward in this species group except for intercastes, because queens have wings and completely developed wing-sclerites. The intercastes, which are the intermediate individuals between worker and queen having the lateral and mid ocelli and the wing sclerites, are sometimes found and all characters of this caste could be varied from absent to incompletely developed in variable degrees. These intercastes could be keyed out as either workers or queens; however, the description of the intercastes was included in the queen section.
The two phenotypes mentioned for the camillae species group are present in the workers of mysticum species group, similarly to the voeltzkowi species group (Table 1). One has a somewhat curved apical tooth (as in Fig. 15A), larger body size, and darker body color; and the other has a straight apical tooth, smaller body size, somewhat narrower body setae, and brighter body color. However, the division of the two phenotypes is weaker than that in the camillae species group, and no other distinct differences could be found between the two phenotypes except for the four characters mentioned above. The “small bright color” phenotype does not seem to be functional in reproduction as in the ergatoid queens of the voeltzkowi species group. In this study, we regard these two phenotypes as morphological variation observed in the worker caste, and we reference the larger one as a major worker and the smaller one as a minor worker when necessary.
http://species-id.net/wiki/Mystrium_mysticum
Figs 4C, 7, 8C, 9A, 9C, 11A, 12A, 26B, 28A, 28C, 29A, 37B, 38B, 39B, 40B, 41B, 43A, 43C, 44A, 44C, 45A, 45C, 46A, 46C, 47A, 47C, 48A, 48C, 49A[here designated]. Alate queen: CASENT0429914, BLF4770, MADAGASCAR, Toliara, Parc National de Kirindy Mite, 16.3 km 127° SE Belo sur Mer (-20.79528°, 44.147°), 80 m alt., 6-10.xii.2001, Fisher-Griswold Arthropod Team leg. [CASC].
[here designated]. Worker: CASENT0104561, MADAGASCAR, Est de I’Imeriua, Sikora leg. [ZMHB: examined].
Description. Measurements: lectotype of Mystrium stadelmanni. HL 2.05, HW 1.93, SL 1.16, ML 2.09, HD 1.26, WL 1.99, PnW 0.94, PpW 0.84, PtW 0.80, PtL 0.53, CI 94.3, SI 59.9, MI 108.2, PpI 89.5, PtI 153.0.
HL 1.44–2.27, HW 1.42–2.18, SL 0.86–1.35, ML 1.32–2.31, HD 0.98–1.39, WL 1.63–2.44, PnW 0.77–1.2, PpW 0.69–1.09, PtW 0.6–0.94, PtL 0.4–0.6, CI 96.0–102.0, SI 56–64, MI 91–108, PpI 86–95, PtI 146–168 (10 specimens measured).
Posterolateral corner of head strongly expanding posteriorly. Posterior face of vertex forming slightly blunt angle with its dorsal face on median line of head, so that declivity of vertex on lateral part slightly steeper than on median part. Ventral half of vertex smooth and not sculptured. Eye small to moderately small. Anterior margin of clypeus convex and with short conical setae. Genal tooth of head undeveloped: anterolateral corner of head angulate. Masticatory margin of mandible widely visible in full-face view, difference in width of dorsal surface of mandible relatively large between mandibular shaft and distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) as long as pedicel (second antennal segment). Shallow and fine longitudinal striae irregularly impressed on central part of pronotal dorsum, sometimes with shallow reticulation around them. On lateral surface of pronotum, shallow, fine longitudinal striae impressed, wide and shallow punctures usually arranged on central horizontal line. Mesonotum differentiated from propodeum in dorsal view, length as long as that of propodeum in large individuals, shorter than propodeum in small individuals. Metanotal groove shallowly impressed in lateral view, mesonotum as high as pronotum in large individuals, higher than pronotum in small individuals. Metanotum weakly developed in largest individuals. Metapleural gland bulla developed, so that propodeal declivity in lateral view weakly convex posteriorly on its ventral portion. Petiole in dorsal view moderately wide, and relatively wider than that of Mystrium rogeri.
Body color brown to black.
Description. Measurements: neotype. HL 2.44, HW 2.51, SL 1.49, ML 2.24, HD 1.54, WL 3.25, MnW 1.73, PtW 1.11, PtL 0.66, CI 102.7, SI 59.3, MI 89.2, MnI 69.1, PtI 168.2.
HL 2.14–2.73, HW 2.08–2.84, SL 1.26–1.60, ML 1.85–2.56, HD 1.32–1.72, WL 2.60–3.48, MnW 1.27–1.92, PtW 0.89–1.33, PtL 0.55–0.77, CI 97.2–104.2, SI 56.3–60.7, MI 87.2–98.2, MnI 61.0–71.3, PtI 160.2–188.8 (10 specimens measured).
Wings usually present and well developed; lacking in intercaste. Wing sclerites fully developed even if wings have dropped off in queen; developed to undeveloped in variable degrees in intercaste. Posterolateral corner of head strongly expanding posteriorly; expansion even stronger than that of workers. Posterior face of vertex forming slightly blunt angle with dorsal face on median line of head, so that declivity of vertex on lateral part slightly steeper than on median part. Ventral half of vertex smooth and not sculptured. Eye well developed. Both anterior and lateral ocelli clearly present and developed in most cases, lateral ones rarely smaller; ocelli varied from absent to developed in intercaste. Anterior margin of clypeus convex with small conical setae. Anterolateral portion of head angulate or with short spine. Masticatory margin of mandible widely visible in full-face view, and dorsal surface on distal portion distinctly wider that on mandibular shaft, difference in width much larger than that in Mystrium rogeri. Mandibular teeth (a row of dentition on dorsal side) often lacking on mid-portion of mandibular shaft. A spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna as long as pedicel. Setae on pronotum almost simple, narrowing distally with strongly sharpened apex. Propodeal declivity in lateral view almost straight and forming right to slight obtuse angle with dorsal margin, ventral portion with small convexity by metapleural gland. Petiole relatively long in dorsal view, about 0.6× length of abdominal segment III.
Body color black.
Description. Measurements: HL 1.13–1.33, HW 1.52–1.81, SL 0.22–0.28, EL 0.62–0.68, WL 2.26–2.86, MnW 1.46–1.89, CI 129.5–136.0, SI 14.3–17.0, EI 49.0–54.8, MnI 96.3–104.4 (7 specimens measured).
Eye relatively small, occupying about 0.5× length of head. Ocelli relatively distant from dorsal margin of head or just failing to reach dorsal margin in full-face view. Dorsal margin of head in full-face view rounded. Both anterior and lateral ocelli small. Distance between lateral ocellus and eye relatively long, about 3× longer than diameter of lateral ocellus. Posterior half of vertex clearly differentiated from dorsal half, its dorsal face almost as long as its posterior face. Palpal formula 4, 3. First segment of maxillary palp flattened and distinctly wider than second segment. Second maxillary palpomere longer than third. Notauli clearly impressed on mesoscutum. Petiole in dorsal view thin, its length about 0.65× that of abdominal tergite III. Petiolar dorsum covered with rough, deep punctures. Abdominal tergum VIII roughly and deeply punctured.
Abdominal sternum IX punctured on its distal portion. Basal ring short, not extending basally. Telomere extending slightly further distally than digitus. Basoventral expansion of aedeagus well developed basoventrally, distinctly longer than dorsal extension. Ventral margin of aedeagus strongly curved ventrally in lateral view. Aedeagus weakly narrowing distally, its distal portion widely rounded.
On forewing, cu-a located at junction of Media (M) and Cubitus (Cu).
Body color reddish brown to black.

Workers of mysticum species group in lateral view. A, C Mystrium mysticum B, C Mystrium rogeri. A major worker (CASENT0429955) B major worker (CASENT0001069) C minor worker (CASENT0429953) D minor worker (CASENT0001154).

Head of the workers of mysticum species group in full-face view. A, C Mystrium mysticum B, D Mystrium rogeri. A major worker (CASENT0429955) B major worker (CASENT0001069) C minor worker (CASENT0429953) D minor worker (CASENT0001154).

Workers of mysticum species group in dorsal view. A, C Mystrium mysticum B, D Mystrium rogeri. A major worker (CASENT0429955) B major worker (CASENT0001069) C minor worker (CASENT0429953) D minor worker (CASENT0001154).

Queens and intercastes of mysticum species group in lateral view. A, C Mystrium mysticum B, D Mystrium rogeri. A alate queen (CASENT0429962) B alate queen (CASENT0001079) C intercaste (CASENT0429959) D intercaste (CASENT0497221).

Head of the queens and intercastes of mysticum species group in full-face view. A, C Mystrium mysticum B, D Mystrium rogeri. A alate queen (CASENT0429962) B alate queen (CASENT0001079) C intercaste (CASENT0429959) D intercaste (CASENT0497221).

Queens and intercastes of mysticum species group in dorsal view. A, C Mystrium mysticum B, D Mystrium rogeri. A alate queen (CASENT0429962) B alate queen (CASENT0001079) C intercaste (CASENT0429959) D intercaste (CASENT0497221).
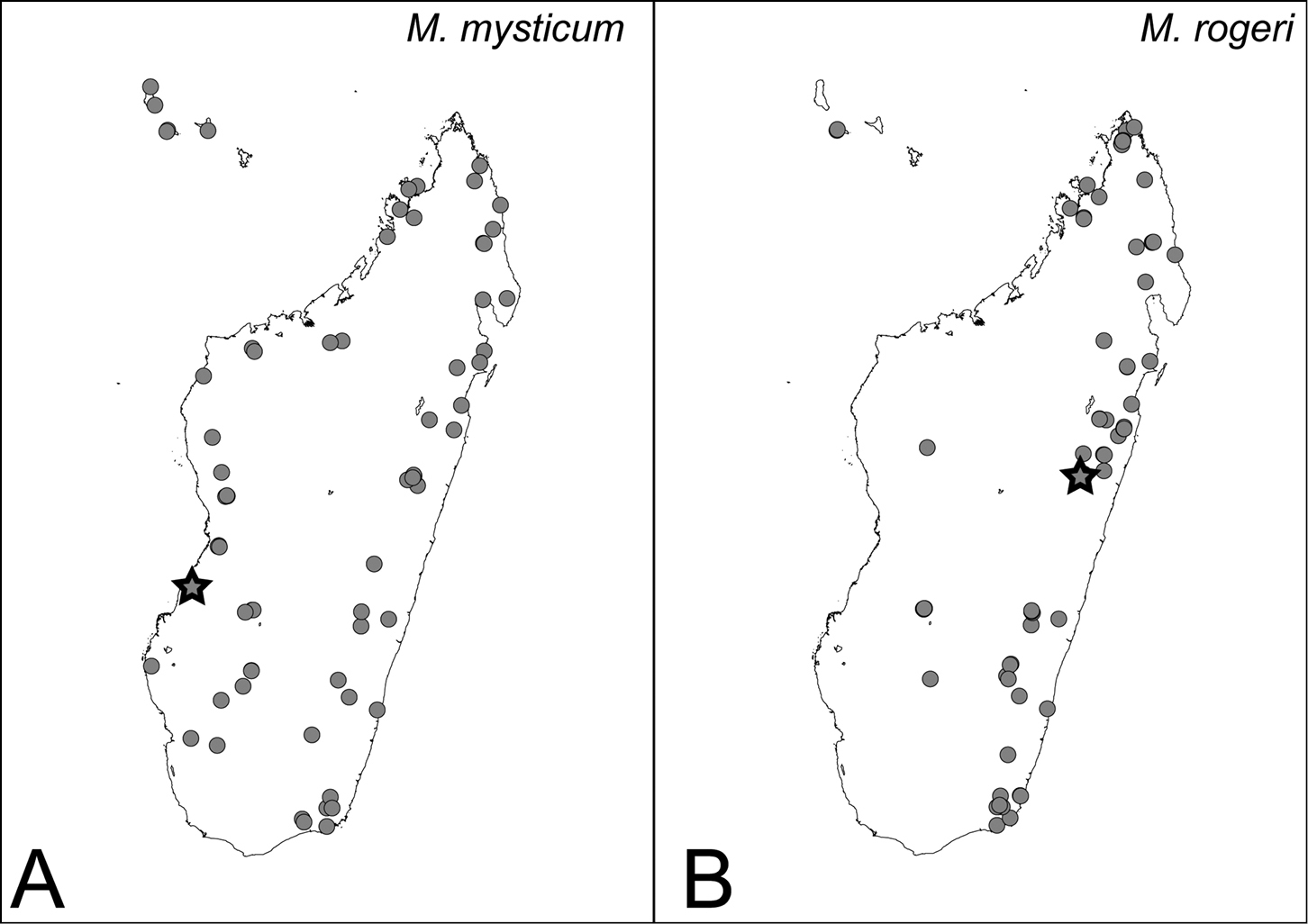
Distribution maps for mysticum species group. A Mystrium mysticum B Mystrium rogeri. Star symbols represent the type locality: the locality for Mystrium mysticum is that of the neotype, and for Mystrium rogeri is approximate, which is assumed from the original description.
MADAGASCAR and COMOROS: as in Figure 49A.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Antsiranana. Forêt d’Ampondrabe, 26.3 km 10° NNE Daraina (-12.97°, 49.7°), tropical dry forest, 175 m alt.; Forêt de Binara, 9.1 km 233° SW Daraina (-13.26333°, 49.60333°), rainforest, 650-800 m alt.; 9.4 km 235° SW Daraina (-13.26333°, 49.6°), montane rainforest, 1100 m alt.; Nosy Faly (-13.3624°, 48.49101°), open secondary vegetation, 15 m alt.; (-13.36435°, 48.49137°), open secondary vegetation, 40 m alt.; Nosy Be, Réserve Naturelle Intégrale de Lokobe, 6.3 km 112° ESE Hellville (-13.41933°, 48.33117°), rainforest, 30 m alt.; Ambondrobe, 41.1 km 175° Vohemar (-13.71533°, 50.10167°), littoral rainforest, 10 m alt.; Ampasindava, Forêt d’Ambilanivy, 3.9 km 181° S Ambaliha (-13.79861°, 48.16167°), rainforest, 600 m alt. R.S. Manongarivo, 10.8 km 229° SW Antanambao (-13.96167°, 48.43333°), rainforest, 400 m alt.; Makirovana forest (-14.17066°, 49.95409°), rainforest, 225 m alt.; Forêt d’Anabohazo, 21.6 km 247° WSW Maromandia (-14.30889°, 47.91433°), tropical dry forest, 120 m alt.; Parc National de Marojejy, Manantenina River, 28.0 km 38° NE Andapa, 8.2 km 333° NNW Manantenina (-14.43667°, 49.775°), rainforest, 450 m alt.; Marojejy R.N.I. #12 (-14.44533°, 49.78564°), rainforest, 375 m alt.; R. Onive (-15.47°, 50.225°), 150 m alt. Toamasina. Nosy Mangabe, 7.43 km S Maroantsetra (-15.4973°, 49.76223°), littoral rainforest, 3 m alt.; Parc National Mananara-Nord, 7.1 km 261° Antanambe (-16.455°, 49.7875°), rainforest, 225 m alt.; Res. Ambodiriana, 4.8 km 306°Manompana, along Manompana river (-16.67233°, 49.70117°), rainforest, 125 m alt.; Manakambahiny, near Vavatenina Forest (-17.46667°, 49.35°); Parc National de Zahamena (-17.73359°, 48.72625°), rainforest, 950 m alt.; Réserve Nationale Intégrale Betampona, Betampona 35.1 km NW Toamasina (-17.91801°, 49.20074°), rainforest, 500 m alt.; Corridor Forestier Analamay-Mantadia, Tsaravoniana (-18.75737°, 48.42302°), rainforest, 1018 m alt.; Tsaravoniana (-18.76124°, 48.42134°), rainforest, 939 m alt.; Ambatoharanana (-18.8024°, 48.40612°), rainforest, 950 m alt.; (-18.80388°, 48.40506°), rainforest, 1013 m alt.; Bevolota 17.1 km N Andasibe (-18.77071°, 48.43164°), montane rainforest, 995 m alt.; Ambatovy, 12.4 km NE Moramanga (-18.83937°, 48.30842°), montane rainforest, 1080 m alt.; Perinet (-18.95°, 48.5°), 800 m alt.; Mahajanga. Réserve d’Ankoririka, 10.6 km 13° NE de Tsaramandroso (-16.26722°, 47.04861°), tropical dry forest, 210 m alt.; Parc National d’Ankarafantsika, Ampijoroa Station Forestière, 5.4 km 331° NW Andranofasika (-16.29889°, 46.813°), tropical dry forest, 70 m alt.; Parc National de Namoroka, 16.9 km 317° NW Vilanandro (-16.40667°, 45.31°), tropical dry forest, 100 m alt.; 9.8 km 300° WNW Vilanandro (-16.46667°, 45.35°), tropical dry forest, 140 m alt.; Réserve Spéciale de Bemarivo, 23.8 km 223° SW Besalampy (-16.925°, 44.36833°), tropical dry forest, 30 m alt.; Réserve forestière Beanka, 54.3 km E Maintirano (-18.06009°, 44.54086°), tropical dry forest on tsingy, 262 m alt.; Parc National Tsingy de Bemaraha, 10.6 km ESE 123° Antsalova (-18.70944°, 44.71817°), tropical dry forest on tsingy, 150 m alt.; 2.5 km 62° ENE Bekopaka, Ankidrodroa River (-19.13222°, 44.81467°), tropical dry forest on tsingy, 100 m alt.; 3.4 km 93° E Bekopaka, Tombeau Vazimba (-19.14194°, 44.828°), tropical dry forest, 50 m alt.; Bekopaka (-19.14667°, 44.796°), village, 100 m alt. Toliara. Forêt de Kirindy, 15.5 km 64° ENE Marofandilia (-20.045°, 44.66222°), tropical dry forest, 100 m alt.; 48 km ENE Morondava, Kirindy Forest (-20.06667°, 44.65°), tropical dry forest, 30 m alt.; (-20.07444°, 44.67611°), tropical dry forest, 100 m alt.; Parc National de Kirindy Mite, 16.3 km 127° SE Belo sur Mer (-20.79528°, 44.147°), tropical dry forest, 80 m alt.; Makay Mts. (-21.21836°, 45.3106°), gallery forest on sandy soil, 510 m alt.; (-21.21985°, 45.32396°), gallery forest on sandy soil, 500 m alt.; (-21.22284°, 45.32477°), gallery forest on sandy soil, 490 m alt.; (-21.22336°, 45.32628°), gallery forest on sandy soil, 480 m alt.; (-21.25978°, 45.17158°), gallery forest with palm and pandanus, 430 m alt.; Forêt de Beroboka, 5.9 km 131° SE Ankidranoka (-22.23306°, 43.36633°), tropical dry forest, 80 m alt.; 29 km NNW Ranohira, Isalo N.P. (-22.31614°, 45.29625°), gallery forest, 490 m alt.; Parc National de Zombitse, 19.8 km 84° E Sakaraha (-22.84333°, 44.71°), tropical dry forest, 770 m alt.; Réserve Spéciale Kalambatritra, Ampanihy (-23.4635°, 46.4631°), montane rainforest, 1270 m alt.; Forêt de Mite, 20.7 km 29° WNW Tongobory (-23.52417°, 44.12133°), gallery forest, 75 m alt.; Beza-Mahafaly, 27 km E Betioky (-23.65°, 44.63333°), tropical dry forest, 135 m alt.; 10 km NW Enakara, Rés. Andohahela (-24.56667°, 46.81667°), rainforest, 425 m alt.; Parc National Andohahela, Col de Tanatana, 33.3 km NW Tolagnaro (-24.7585°, 46.85367°), rainforest, 275 m alt.; Parc National d’Andohahela, Col du Sedro, 3.8 km 113° ESE Mahamavo, 37.6 km 341° NNW Tolagnaro (-24.76389°, 46.75167°), montane rainforest, 900 m alt.; Réserve Privé Berenty, Forêt de Bealoka, Mandraré River, 14.6 km 329° NNW Amboasary (-24.95694°, 46.2715°), gallery forest, 35 m alt.; Forêt de Malaza, Mandraré River, 8.6 km 314° NW Amboasary (-25.00778°, 46.306°), gallery forest, 40 m alt.; Grand Lavasoa, 25.9 km W Tolagnaro (-25.08767°, 46.749°), rainforest, 450 m alt. Fianarantsoa. SE of Fandriana, Korikory, (-20.38444°, 47.66722°), 1670 m alt.; 7 km W Ranomafana National Park (-21.25°, 47.41667°), montane rainforest, 900 m alt.; 8 km E Kianjavato, Vatovavy Forest (-21.38861°, 47.94361°), lowland forest, 145 m alt.; Andrambovato along river Tatamaly (-21.50967°, 47.40762°), cultivated land (tavy), 984 m alt.; Parc National d’Isalo, Sahanafa River, 29.2 km 351° N Ranohira (-22.31333°, 45.29167°), gallery forest, 500 m alt.; 30 km NNW Ranohira, Isalo N.P. (-22.31722°, 45.29333°), gallery forest, 455 m alt.; R.S. Ivohibe 8.0 km E Ivohibe (-22.48333°, 46.96833°), montane rainforest, 1200 m alt.; Forêt d’Analalava, 29.6 km 280° W Ranohira (-22.59167°, 45.12833°), tropical dry forest, 700 m alt.; Forêt de Vevembe, 66.6 km 293° Farafangana (-22.791°, 47.18183°), rainforest, transition to montane forest, 600 m alt.; Réserve Speciale Manombo 24.5 km 228° Farafangana (-23.01583°, 47.719°), rainforest, 30 m alt.
COMOROS. Grande Comore. Grillé (-11.47578°, 43.34669°), montane rainforest, 995 m alt.; Karthala (-11.82699°, 43.4295°), montane rainforest, 1000 m alt. Mohéli. Ouallah (-12.29696°, 43.67392°), rainforest, 680 m alt.; (-12.32717°, 43.65952°), coastal scrub, 10 m alt. Anjouan. (-12.30537°, 44.45031°), along roadside, mango, banana, 500 m alt.
Mystrium mysticum females can be distinguished easily from other Mystrium females by a combination of a distinct longitudinal carina on the central portion of the labrum (Fig. 8A) or a single lateral spine on abdominal sternum VII (Fig. 8C), and the convexity of the anterior margin of the clypeus (Fig. 9C). In addition to these diagnostic characters, a combination of unsculptured and smooth areas on the ventral portion of the vertex (Fig. 11A) and the convexity of the anterior margin of the clypeus (as in Fig. 9C) separates Mystrium mysticum from the other Mystrium species in workers; in larger-sized alate queens (HW>2.00), simple setae on the pronotal dorsum (Fig. 9A) separate Mystrium mysticum from the queens of other species. For males, the small eye and small ocelli that are separated from each other by more than 3× the maximum diameter of lateral ocellus (Fig. 26B), petiolar dorsum (Fig. 28C) and abdominal tergum VIII (Fig. 28A) with rough and deep punctures, and well-developed basoventral expansion of aedeagus (Fig. 29A) separate Mystrium mysticum from the other Mystrium males in the Malagasy region.
Several diagnostic characters for Mystrium mysticum have been given in previous papers, (e.g.
The redescription of the male of Mystrium mysticum in this study may be the first actual description of the male of this species. Previous male descriptions for Mystrium mysticum have been given in
Clarification of the male-worker association in Mystrium mysticum in this study marks a great advance; however, some problems remain, especially in the level of confidence in diagnosing males. External characters, e.g. strong and rough punctures on the petiolar dorsum (Fig. 28C) and abdominal tergite VIII (Fig. 28A), distinguish the mysticum species group from the other Malagasy species. However, a specific character is neither possible nor practical to distinguish Mystrium mysticum males from Mystrium rogeri males. In fact, although we proposed the aedeagal character to separate Mystrium mysticum and Mystrium rogeri (Fig. 29), this difference cannot be confirmed without dissection. Unexpected remarkable variation could exist in the genital character as well as the other external characters. Further and more thorough taxonomic examinations of males are necessary to find diagnostic characters able to separate these two species.
The neotype of Mystrium mysticum was designated in this study because we concluded that the original type specimens of this “mysterious species” are lost. Our study revealed that no one examined any of the original type specimens after the publication of the original description by
A queen was chosen as the neotype because the original syntypes were two queens. The queen caste also offers a greater number of characters to distinguish this taxon from others in this species group. The location of the neotype was not based on a proximity to the original syntype series, but on the availably of a large collection series that included workers, males and queens.
Mystrium stadelmanni Forel, 1895 is synonymized with Mystrium mysticum in this study. Mystrium stadelmanni is described as a species having a longer petiole, narrower mesosoma, and less constricted abdomen compared with Mystrium mysticum; however, our observations reveal that the boundary of this species is doubtful. None of the diagnostic characters above represent more than intraspecific variation observed in a series of workers from a single colony of Mystrium mysticum. The fact that syntypes of Mystrium stadelmanni consist of two minor workers of Mystrium mysticum and one minor worker of Mystrium rogeri certifies that Forel’s determination (
Updating species boundaries and providing easier identification tools for Mystrium mysticum will help reorganize and connect existing ecological and phylogenetic information to proper species names in Mystrium. This taxonomic revision permits us to update identifications in recent ecological publications (
http://species-id.net/wiki/Mystrium_rogeri
Figs 4A, 8A, 9B, 9D, 12B, 29B, 37C, 38C, 39C, 40C, 41C, 43B, 43D, 44B, 44D, 45B, 45D, 46B, 46D, 47B, 47D, 48B, 48D, 49B[here designated]. Worker: CASENT0101670, MADAGASCAR, Amparafaravantsiv Moramangoufer [MHNG: examined].
Description. Measurements: lectotype. HL 1.87, HW 1.85, SL 1.13, ML 1.72, HD 1.24, WL 2.09, PnW 1.14, PpW 0.98, PtW 0.92, PtL 0.53, CI 98.7, SI 61.4, MI 93.3, PpI 86.5, PtI 174.6.
HL 1.19–2.27, HW 1.12–2.28, SL 0.70–1.37, ML 1.18–2.38, HD 0.83–1.47, WL 0.18–2.52, PnW 0.67–1.23, PpW 0.57–1.05, PtW 0.51–1.04, PtL 0.33–0.65, CI 93.9–101.4, SI 59.2–64.6, MI 90.9–105.7, PpI 79.3–90.1, PtI 153.1–182.6 (10 specimens measured).
Posterolateral corner of head strongly expanding posteriorly. Posterior face of vertex forming slightly blunt angle with dorsal face on median line of head, so that declivity of vertex on lateral part slightly steeper than on median part. Ventral half of vertex smooth and not sculptured. Eye small to moderately small. Anterior margin of clypeus straight or weakly concave and with small conical setae. Genal tooth of head undeveloped, but anterolateral corner of head angulate. Masticatory margin of mandible narrowly visible in full-face view, and difference in width of dorsal surface of mandible relatively small between mandibular shaft and distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) as long as pedicel (second antennal segment). Shallow and fine longitudinal striae irregularly impressed on central part of pronotal dorsum, and sometimes shallowly reticulated around those striae. On lateral surface of pronotum, deep, fine longitudinal striae impressed and wide and deep punctures arranged on central horizontal line. Mesonotum differentiated from propodeum in dorsal view, length as long as that of propodeum in large individuals, shorter than propodeum in small individuals. Metanotal groove shallowly impressed, mesonotum in lateral view as high as pronotum in large individuals, higher than pronotum in small individuals. Metanotum weakly developed in largest individuals. Metapleural gland bulla developed, so that propodeal declivity in lateral view weakly convex posteriorly on ventral portion. Petiole in dorsal view moderately narrow, relatively narrower than that of Mystrium mysticum.
Body color brown to black.
Description. Measurements: HL 1.94–2.31, HW 1.99–2.40, SL 1.20–1.42, ML 1.87–2.48, HD 1.29–1.51, WL 2.78–3.31, MnW 1.42–1.83, PtW 1.03–1.32, PtL 0.61–0.78, CI 99.8–104.2, SI 58.1–62.8, MI 90.4–103.1, MnI 68.1–76.1, PtI 155.3–185.1 (10 specimens measured).
Wings usually present and well developed; but lacking in intercaste. Wing sclerites fully developed even if wings have dropped off in queen; developed to undeveloped in variable degrees in intercaste. Posterolateral corner of head strongly expanding posteriorly, expansion not differentiated from that of workers. Posterior face of vertex forming slightly blunt angle with dorsal face on median line of head, so that declivity of vertex on lateral part slightly steeper than on median part. Ventral half of vertex smooth and not sculptured. Eye well developed. Both anterior and lateral ocelli clearly present in queen; ocelli varied from absent to developed in intercaste. Anterior margin of clypeus straight or weakly concave with small conical setae. Anterolateral portion of head weakly to strongly angulate or with short spine. Masticatory margin of mandible narrowly, slightly visible in full-face view, dorsal surface on distal portion moderately wider than on mandibular shaft, difference in width much smaller than that in Mystrium mysticum. Teeth always present on whole mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna as long as pedicel. Setae on pronotum spatulate, widened distally with sharp or blunt apex. Propodeal declivity in lateral view almost straight and forming right to slightly obtuse angle with its dorsal margin, of which ventral portion with small convexity by metapleural gland. Petiole relatively long in dorsal view, about 0.6× length of abdominal segment III.
Body color black.
Description. Measurements: HL 1.10–1.41, HW 1.43–1.85, SL 0.25–0.34, EL 0.59–0.76, WL 2.27–3.03, MnW 1.46–1.96, CI 127.9–132.0, SI 16.2–20.0, EI 49.9–54.1, MnI 101.4-107.6 (6 specimens measured).
Eye relatively small, occupying about half of head length. Ocelli relatively distant from dorsal margin of head in full-face view, or just failing to reach dorsal margin. Dorsal margin of head in full-face view rounded. Both anterior and lateral ocelli small. Distance between lateral ocellus and eye long, 3× longer than diameter of lateral ocellus. Posterior half of vertex clearly differentiated from dorsal half, dorsal face almost as long as posterior face. Palpal formula 4, 3. First segment of maxillary palp flattened and distinctly wider than second segment. Second maxillary palpomere longer than third. Notauli clearly impressed on mesoscutum. Petiole in dorsal view thin, its length about 0.5-0.65× that of abdominal tergite III. Petiolar dorsum covered with rough, deep punctures. Abdominal tergum VIII roughly and deeply punctured.
Abdominal sternum IX punctured on its distal portion. Basal ring short, not extending basally. Telomere extending slightly further distally than digitus. Basoventral expansion of aedeagus less developed basoventrally, as long as dorsal extension. Ventral margin of aedeagus gently curved ventrally in lateral view. Aedeagus weakly narrowing distally, its distal portion widely rounded.
On forewing, cu-a located at junction of Media (M) and Cubitus (Cu).
Body color reddish brown to black.
MADAGASCAR and COMOROS: as in Figure 49B.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Diego-Suarez. Sakalava Beach [vegetated beach dunes] (-12.26278°, 49.3975°), across sandy trail in dwarf littoral forest, 10 m alt.; 7 km N Joffreville [camp 2 of Fisher] (-12.33333°, 49.25°), in dry forest, 360 m alt.; Parc National Montagne d’Ambre [lemur trail] (-12.51667°, 49.18333°), rainforest, 975 m alt.; Parc National Montagne d’Ambre [Petit Lac road] (-12.52028°, 49.17917°), rainforest, 1125 m alt.; Antsiranana. Parc National Montagne d’Ambre, Antomboka (-12.51269°, 49.17807°), montane rainforest, 970 m alt.; Parc National Montagne d’Ambre (-12.51389°, 49.17784°), montane rainforest, 984 m alt.; Pic Bades (-12.5186°, 49.18625°), montane rainforest, 900 m alt.; (-12.52306°, 49.17901°), montane rainforest, 1100 m alt.; Roussettes (-12.52574°, 49.17238°), montane rainforest, 1025 m alt.; Roussettes Camp 7 km SW Park entrance (-12.51444°, 49.18139°), rainforest, 960 m alt.; Mahasarika (-12.53176°, 49.17662°), montane rainforest, 1135 m alt.; (-12.53417°, 49.17607°), montane rainforest, 1325 m alt.; 3.6 km 235° SW Joffreville (-12.53444°, 49.1795°), montane rainforest, 925 m alt.; Petit lac (-12.53664°, 49.17412°), montane rainforest, 1130 m alt.; Lac Maudit (-12.58502°, 49.15147°), montane rainforest, 1250 m alt.; 12.2 km 211° SSW Joffreville (-12.59639°, 49.1595°), montane rainforest, 1300 m alt.; Forêt de Binara, 9.1 km 233° SW Daraina (-13.26333°, 49.60333°), rainforest, 800 m alt.; (-13.26333°, 49.6°), montane rainforest, 1100 m alt.; Nosy Faly (-13.3624°, 48.49101°), open secondary vegetation, 15 m alt.; Galoko chain, Mont Galoko (-13.58487°, 48.71818°), rainforest, 520 m alt.; Ampasindava, Forêt d’Ambilanivy, 3.9 km 181° S Ambaliha (-13.79861°, 48.16167°), rainforest, 600 m alt.; R.S. Manongarivo, 12.8 km 228° SW Antanambao (-13.97667°, 48.42333°), rainforest, 780 m alt.; 14.5 km 220° SW Antanambao (-13.99833°, 48.42833°), montane rainforest, 1175 m alt.; Parc National de Marojejy, Manantenina River, 27.6 km 35° NE Andapa, 9.6 km 327° NNW Manantenina (-14.435°, 49.76°), rainforest, 775 m alt.; 28.0 km 38° NE Andapa, 8.2 km 333° NNW Manantenina (-14.43667°, 49.775°), rainforest, 450 m alt.; Antranohofa, 26.6 km 31° NNE Andapa, 10.7 km 318° NW Manantenina (-14.44333°, 49.74333°), montane rainforest, 1325 m alt.; Betaolana Forest, along Bekona River (-14.52996°, 49.44039°), rainforest, 880 m alt.; Forêt Ambanitaza, 26.1 km 347° Antalaha (-14.67933°, 50.18367°), rainforest, 240 m alt.; Toamasina. Montagne d’Anjanaharibe, 18.0 km 21° NNE Ambinanitelo (-15.18833°, 49.615°), rainforest, 470 m alt.; Res. Ambodiriana, 4.8 km 306°Manompana, along Manompana river (-16.67233°, 49.70117°), rainforest, 125 m alt.; Réserve Spéciale Ambatovaky, Sandrangato river (-16.7633°, 49.26692°), rainforest, 520 m alt.; (-16.7702°, 49.26638°), rainforest, 470 m alt.; (-16.77274°, 49.26551°), rainforest, 450 m alt.; Manakambahiny, near Vavatenina Forest (-17.46667°, 49.35°), alt.; Parc National de Zahamena (-17.73359°, 48.72625°), rainforest, 950 m alt.; Tetezambatana forest, near junction of Nosivola and Manakambahiny Rivers (-17.74298°, 48.72936°), rainforest, 860 m alt.; Onibe River (-17.75908°, 48.85468°), rainforest, 780 m alt.; Reserve Betampona, Camp Vohitsivalana, 37.1 km 338° Toamasina (-17.88667°, 49.2025°), rainforest, 520 m alt.; Réserve Nationale Intégrale Betampona, Betampona 35.1 km NW Toamasina (-17.91801°, 49.20074°), rainforest, 500 m alt.; Réserve Naturelle Betampona, 34.08 km 332° Toamasina (-17.91977°, 49.20039°), rainforest, 525 m alt.; Reserve Betampona, Camp Rendrirendry 34.1 km 332° Toamasina (-17.924°, 49.19967°), rainforest, 390 m alt.; F.C. Sandranantitra (-18.04833°, 49.09167°), rainforest, 450 m alt.; Station forestière Analamazaotra, Analamazaotra 1.3 km S Andasibe (-18.38466°, 48.41271°), montane rainforest, 980 m alt.; Ankerana (-18.40062°, 48.81311°), rainforest, 865 m alt.; Ankerana (-18.4017°, 48.80605°), montane forest, 1035 m alt.; (-18.4061°, 48.82029°), rainforest, 725 m alt.; (-18.40829°, 48.82107°), rainforest, 750 m alt.; (-18.4104°, 48.8189°), rainforest, 855 m alt.; F.C. Andriantantely (-18.695°, 48.81333°), rainforest, 530 m alt.; Corridor Forestier Analamay-Mantadia, Tsaravoniana (-18.75641°, 48.42195°), rainforest, 1036 m alt.; (-18.75737°, 48.42302°), rainforest, 1018 m alt.; (-18.76124°, 48.42134°), rainforest, 939 m alt.; (-18.76465°, 48.41938°), rainforest, 1039 m alt.; Ambohibolakely (-18.76087°, 48.37128°), rainforest, 1044 m alt.; (-18.76131°, 48.36437°), rainforest, 983 m alt.; (-18.77898°, 48.36375°), rainforest, 918 m alt.; Ambohibolakely (-18.77908°, 48.36628°), rainforest, 1014 m alt.; Ambatoharanana (-18.79956°, 48.4028°), rainforest, 1058 m alt.; (-18.80388°, 48.40506°), rainforest, 1013 m alt.; (-18.80398°, 48.40358°), rainforest, 1064 m alt.; (-18.80424°, 48.40081°), rainforest, 968 m alt.; (-18.80438°, 48.40735°), rainforest, 960 m alt.; Bevolota 17.1 km N Andasibe (-18.77071°, 48.43164°), montane rainforest, 995 m alt.; Analamay (-18.80623°, 48.33707°), montane rainforest, 1068 m alt.; Parc National d´ Andasibe-Mantadia, Forêt de Mantadia, 25.7 km 248° Moramanga (-18.81402°, 48.43028°), rainforest, 1040 m alt.; Ambatovy, 12.4 km NE Moramanga (-18.83937°, 48.30842°), montane rainforest, 1080 m alt.; Forêt Ambatovy, 14.3 km 57° Moramanga (-18.85083°, 48.32°), montane rainforest, 1075 m alt.; Torotorofotsy (-18.87082°, 48.34737°), montane rainforest, marsh edge, 1070 m alt.; Toliara. Réserve Spéciale d’Ambohijanahary, Forêt d’Ankazotsihitafototra, 35.2 km 312° NW Ambaravaranala (-18.26667°, 45.40667°), montane rainforest, 1050 m alt.; Makay Mts. (-21.20978°, 45.34184°), Gallery forest on sandy soil, 525 m alt.; (-21.21013°, 45.35462°), Gallery forest with bamboo, 540 m alt.; (-21.22336°, 45.32628°), Gallery forest on sandy soil, 480 m alt.; (-21.227°, 45.33222°), gallery forest on sandy soil, 475 m alt.; Forêt Ivohibe 55.6 km N Tolagnaro (-24.56167°, 47.20017°), rainforest, 650 m alt.; 11 km NW Enkara, Rés Andohahela (-24.56667°, 46.81667°), rainforest, 950 m alt.; Forêt Ivohibe 55.0 km N Tolagnaro (-24.569°, 47.204°), rainforest, 200 m alt.; 6 km SSW Eminiminy, Res. Andohahela (-24.73333°, 46.8°), wet forest, 330 m alt.; Rés. Andohahela, 6 km SSW Eminiminy (-24.73333°, 46.8°), rainforest, 330 m alt.; Parc National Andohahela, Col de Tanatana, 33.3 km NW Tolagnaro (-24.7585°, 46.85367°), rainforest, 275 m alt.; Manampanihy River, 5.4 km 113° ESE Mahamavo, 36.7 km 343° NNW Tolagnaro (-24.76389°, 46.76683°), rainforest, 650 m alt.; Col du Sedro, 3.8 km 113° ESE Mahamavo, 37.6 km 341° NNW Tolagnaro (-24.76389°, 46.75167°), montane rainforest, 900 m alt.; Forêt Mandena 8.5 km N Tolagnaro (-24.95267°, 47.0025°), littoral rainforest, 20 m alt.; Grand Lavasoa, 25.9 km W Tolagnaro (-25.08767°, 46.749°), rainforest, 450 m alt.; Toamasina. Res. Ambodiriana, 4.8 km 306° Manompana, along Manompana river (-16.67233°, 49.70117°), rainforest, 125 m alt.; Réserve Spéciale Ambatovaky, Sandrangato river (-16.7633°, 49.26692°), rainforest, 520 m alt.; (-16.7702°, 49.26638°), rainforest, 470 m alt.; (-16.77274°, 49.26551°), rainforest, 450 m alt.; Manakambahiny, near Vavatenina Forest (-17.46667°, 49.35°), alt.; Parc National de Zahamena (-17.73359°, 48.72625°), rainforest, 950 m alt.; Tetezambatana forest, near junction of Nosivola and Manakambahiny Rivers (-17.74298°, 48.72936°), rainforest, 860 m alt.; Onibe River (-17.75908°, 48.85468°), rainforest, 780 m alt.; Reserve Betampona, Camp Vohitsivalana, 37.1 km 338° Toamasina (-17.88667°, 49.2025°), rainforest, 520 m alt.; Réserve Nationale Intégrale Betampona, Betampona 35.1 km NW Toamasina (-17.91801°, 49.20074°), rainforest, 500 m alt.; Réserve Naturelle Betampona, 34.08 km 332° Toamasina (-17.91977°, 49.20039°), rainforest, 525 m alt.; Reserve Betampona, Camp Rendrirendry 34.1 km 332° Toamasina (-17.924°, 49.19967°), rainforest, 390 m alt.; F.C. Sandranantitra (-18.04833°, 49.09167°), rainforest, 450 m alt.; Station forestière Analamazaotra, Analamazaotra 1.3 km S Andasibe (-18.38466°, 48.41271°), montane rainforest, 980 m alt.; Ankerana (-18.40062°, 48.81311°), rainforest, 865 m alt.; Ankerana (-18.4017°, 48.80605°), montane forest, 1035 m alt.; (-18.4061°, 48.82029°), rainforest, 725 m alt.; (-18.40829°, 48.82107°), rainforest, 750 m alt.; (-18.4104°, 48.8189°), rainforest, 855 m alt.; F.C. Andriantantely (-18.695°, 48.81333°), rainforest, 530 m alt.; Corridor Forestier Analamay-Mantadia, Tsaravoniana (-18.75641°, 48.42195°), rainforest, 1036 m alt.; (-18.75737°, 48.42302°), rainforest, 1018 m alt.; (-18.76124°, 48.42134°), rainforest, 939 m alt.; (-18.76465°, 48.41938°), rainforest, 1039 m alt.; Ambohibolakely (-18.76087°, 48.37128°), rainforest, 1044 m alt.; (-18.76131°, 48.36437°), rainforest, 983 m alt.; (-18.77898°, 48.36375°), rainforest, 918 m alt.; Ambohibolakely (-18.77908°, 48.36628°), rainforest, 1014 m alt.; Ambatoharanana (-18.79956°, 48.4028°), rainforest, 1058 m alt.; (-18.80388°, 48.40506°), rainforest, 1013 m alt.; (-18.80398°, 48.40358°), rainforest, 1064 m alt.; (-18.80424°, 48.40081°), rainforest, 968 m alt.; (-18.80438°, 48.40735°), rainforest, 960 m alt.; Bevolota 17.1 km N Andasibe (-18.77071°, 48.43164°), montane rainforest, 995 m alt.; Analamay (-18.80623°, 48.33707°), montane rainforest, 1068 m alt.; Parc National d´ Andasibe-Mantadia, Forêt de Mantadia, 25.7 km 248° Moramanga (-18.81402°, 48.43028°), rainforest, 1040 m alt.; Ambatovy, 12.4 km NE Moramanga (-18.83937°, 48.30842°), montane rainforest, 1080 m alt.; Forêt Ambatovy, 14.3 km 57° Moramanga (-18.85083°, 48.32°), montane rainforest, 1075 m alt.; Torotorofotsy (-18.87082°, 48.34737°), montane rainforest, marsh edge, 1070 m alt.; Toliara. Réserve Spéciale d’Ambohijanahary, Forêt d’Ankazotsihitafototra, 35.2 km 312° NW Ambaravaranala (-18.26667°, 45.40667°), montane rainforest, 1050 m alt.; Makay Mts. (-21.20978°, 45.34184°), gallery forest on sandy soil, 525 m alt.; (-21.21013°, 45.35462°), gallery forest with bamboo, 540 m alt.; (-21.22336°, 45.32628°), gallery forest on sandy soil, 480 m alt.; (-21.227°, 45.33222°), gallery forest on sandy soil, 475 m alt.; Forêt Ivohibe 55.6 km N Tolagnaro (-24.56167°, 47.20017°), rainforest, 650 m alt.; 11 km NW Enkara, Rés Andohahela (-24.56667°, 46.81667°), rainforest, 950 m alt.; Forêt Ivohibe 55.0 km N Tolagnaro (-24.569°, 47.204°), rainforest, 200 m alt.; 6 km SSW Eminiminy, Res. Andohahela (-24.73333°, 46.8°), wet forest, 330 m alt.; Rés. Andohahela, 6 km SSW Eminiminy (-24.73333°, 46.8°), rainforest, 330 m alt.; Parc National Andohahela, Col de Tanatana, 33.3 km NW Tolagnaro (-24.7585°, 46.85367°), rainforest, 275 m alt.; Manampanihy River, 5.4 km 113° ESE Mahamavo, 36.7 km 343° NNW Tolagnaro (-24.76389°, 46.76683°), rainforest, 650 m alt.; Col du Sedro, 3.8 km 113° ESE Mahamavo, 37.6 km 341° NNW Tolagnaro (-24.76389°, 46.75167°), montane rainforest, 900 m alt.; Forêt Mandena 8.5 km N Tolagnaro (-24.95267°, 47.0025°), littoral rainforest, 20 m alt.; Grand Lavasoa, 25.9 km W Tolagnaro (-25.08767°, 46.749°), rainforest, 450 m alt. Mahajanga. Réserve Spéciale Marotandrano, Marotandrano 48.3 km S Mandritsara (-16.28322°, 48.81443°), transition humid forest, 865 m alt. Fianarantsoa. Ranomafana Nat. Park, Miaranony Forest (-21.25°, 47.41667°), montane forest, 700 m alt.; 7 km W Ranomafana (-21.26667°, 47.41667°), montane rainforest, 1000 m alt.; Parc National de Ranomafana, Vatoharanana River, 4.1 km 231° SW Ranomafana (-21.29°, 47.43333°), montane rainforest, 1100 m alt.; 7.6 km 122° Kianjavato, Forêt Classée Vatovavy (-21.4°, 47.94°), rainforest, 175 m alt.; Andrambovato along river Tatamaly (-21.50967°, 47.40762°), cultivated land (tavy), 984 m alt.; (-21.51089°, 47.40987°), rainforest, 994 m alt.; 2 km W Andrambovato, along river Tatamaly (-21.51167°, 47.41°), montane rainforest, 1075 m alt.; 45 km S. Ambalavao (-22.21667°, 47.01667°), rainforest, 785 m alt.; 43 km S Ambalavao, Rés. Andringitra (-22.23333°, 47°), rainforest, 825 m alt.; 9.0 km NE Ivohibe (-22.42667°, 46.93833°), montane rainforest, 900 m alt.; Parc National d’Isalo, 9.1 km 354° N Ranohira (-22.48167°, 45.46167°), gallery forest, 725 m alt.; R.S. Ivohibe 8.0 km E Ivohibe (-22.48333°, 46.96833°), montane rainforest, 1200 m alt.; Forêt de Vevembe, 66.6 km 293° Farafangana (-22.791°, 47.18183°), rainforest, transition to montane forest, 600 m alt.; Réserve Speciale Manombo 24.5 km 228° Farafangana (-23.01583°, 47.719°), rainforest, 30 m alt.; Parc National Befotaka-Midongy, Papango 27.7 km S Midongy-Sud, Mount Papango (-23.83517°, 46.96367°), rainforest, 940 m alt.
COMOROS. Mohéli, Ouallah (-12.32717°, 43.65952°), coastal scrub, 10 m alt.
The females of Mystrium rogeri can be distinguished easily from other Mystrium females by the combination of a distinct longitudinal carina on the central portion of the labrum (Fig. 8A) or a single lateral spine on abdominal sternum VII (Fig. 8C), and the straight anterior margin of the clypeus (Fig. 9D). In addition to these diagnostic characters, a combination of unsculptured and smooth areas on the ventral portion of the vertex (as in Fig. 11A) and the straight anterior clypeal margin separates Mystrium rogeri from the other Mystrium species workers; and a larger-sized alate queen (HW>1.99) with spatulate setae on the pronotal dorsum (Fig. 9B) separate Mystrium rogeri from the queens of other species. For males, the small eye and small ocelli (as in Fig. 26B) that are separated from each other by a distance of more than 3× the maximum diameter of lateral ocellus, petiolar dorsum (as in Fig. 28C) and abdominal tergum VIII (as in Fig. 28A) with rough and deep punctures, and less developed basoventral expansion of aedeagus (Fig. 29B) separate Mystrium rogeri from the other Mystrium males in the Malagasy region.
We propose a new set of diagnostic characters in this study to correctly identify Mystrium rogeri. The diagnostic characters listed above are constant in females regardless of their body size. Because of the remarkable morphological variation in each species, most diagnostic characters for Mystrium rogeri proposed in previous papers (e.g.
As we discussed in Mystrium mysticum, further studies are necessary to better separate the males of Mystrium rogeri and Mystrium mysticum. Here we propose only an aedeagal character to separate these very similar males (Fig. 29).
The lectotype for Mystrium rogeri is designated. Mystrium rogeri was described by Forel in 1899 using at least two workers and one male collected in Madagascar, and we confirm one worker and one male in his collection. Although we could not find any information to identify which type specimens were used in his original description (
Mystrium rogeri is one of the most common Mystrium species used in recent ecological and phylogenetic studies (
The voeltzkowi species group is endemic to the Malagasy region, and consists of six species: Mystrium eques sp. n., Mystrium janovitzi sp. n., Mystrium mirror sp. n., Mystrium oberthueri Forel, 1897, Mystrium shadow sp. n., and Mystrium voeltzkowi Forel, 1897. The complete lack of alate queens and the presence of ergatoid queens that are relatively smaller than conspecific workers are unique characters in this group.
In this study, we define ergatoid queens as the individuals having a mandible with an apical tooth that is moderately sclerotized and directed ventrally (Fig. 15B; Table 1). On the other hand, the workers are defined as individuals having a mandible with an apical tooth that is hardly sclerotized and curved inward (Fig. 15A). This definition for the ergatoid queen in the voeltzkowi species group is assumed on the basis of previous ecological reports by Molet et al. (
http://zoobank.org/0FF584D9-44A0-44C5-AF5E-BA17BF4ACE7B
http://species-id.net/wiki/Mystrium_eques
Figs 18B, 18D, 18F, 21A, 22A, 50A, 51A, 52A, 53A, 54A, 55A, 56AWorker: CASENT0418314, BLF04691, MADAGASCAR, Toamasina, Tampolo, Masoala Peninsula, 40.4 km 154° SSE Maroantsetra (-15.73°, 49.96°), 30 m alt., 28.xi.–3.xii.2001, B.L.Fisher & H.J.Ratsirarson leg. [CASC].
13 workers: CASENT0317390 [CASC], CASENT0317393 [CASC], CASENT0317394 [CASC], CASENT0317395 [CASC], CASENT0317396 [MSNG], CASENT0317397 [MHNG], CASENT0317398 [BMNH], CASENT0317399 [MCZC], CASENT0317443 [CASC], CASENT0418312 [MNHN], CASENT0418313 [CASC], CASENT0418316 [NHMB], CASENT0418318 [ZMHB]; 5 ergatoid queens: CASENT0418311 [CASC], CASENT0418315 [BMNH], CASENT0418317 [MHNG], CASENT0317391 [MCZC], CASENT0317392 [NHMB], with same data as holotype.
Description. Measurements: holotype. HL 1.78, HW 1.65, SL 1.23, ML 1.87, HD 1.08, WL 2.01, PnW 0.94, PpW 0.83, PtW 0.85, PtL 0.57, CI 92.6, SI 74.4, MI 113.8, PpI 88.6, PtI 150.4.
HL 1.72–2.11, HW 1.59–2, SL 1.26–1.42, ML 1.81–2.18, HD 1.08–1.33, WL 2.07–2.34, PnW 0.92–1.1, PpW 0.83–1.02, PtW 0.83–1.03, PtL 0.54–0.67, CI 91.4–97.3, SI 70.4–78.9, MI 107.4–114.2, PpI 84.7–92.7, PtI 149.5–161.2 (10 specimens measured).
Posterolateral corner of head strongly to moderately expanding posteriorly. Posterior face of vertex forming almost a right angle with dorsal face on median line of head, so that declivity of vertex on lateral part as steep as that on median part. Whole region of vertex finely striated. Eye relatively smaller than that of Mystrium oberthueri. Anterior margin of clypeus strongly convex with long conical setae. Genal tooth of head relatively long, as long as lateral lobe of clypeus. Masticatory margin of mandible almost invisible in full-face view. Width of dorsal surface of mandible on distal portion slightly wider than that on mandibular shaft. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) as long as pedicel (second antennal segment). Strong, deep and thick longitudinal striae regularly impressed on whole central part of pronotal dorsum. Strong, deep, and thick longitudinal striae impressed on lateral surface of pronotum. Mesonotum differentiated from propodeum in dorsal view, length slightly shorter than that of propodeum. Metanotal groove shallowly and gently impressed, and mesonotum higher than pronotum in lateral view. Metapleural gland bulla moderately developed, and propodeal declivity in lateral view almost straight. Petiole widened on posterior 1/4 and gently narrowed anteriorly in dorsal view, anterior margin straight to gently rounded and sometimes edged by thin striae.
Body color blackish brown. Appendages brighter, and four distal segments of antennal club yellowish.
Description. Measurements: HL 1.48–1.62, HW 1.41–1.57, SL 1.10–1.25, ML 1.38–1.56, HD 0.99–1.09, WL 1.95–2.10, PnW 0.85–0.95, PpW 0.83–0.94, PtW 0.87–0.97, PtL 0.52–0.56, CI 94.0–97.2, SI 77.7–83.9, MI 93.2–107.0, PpI 93.8–101.6, PtI 160.5–172.6 (7 specimens measured).
Wings vestigial and forming small but distinct appendages. Wing sclerites undeveloped. Posterolateral corner of head strongly expanding posteriorly, expansion relatively weaker than that of workers. Posterior face of vertex forming almost a right angle with dorsal face on median line of head, so that declivity of vertex on lateral part as steep as on median part. Ventral half of vertex sculptured. Eye small but distinct. Ocelli absent. Anterior margin of clypeus distinctly convex with long to short conical setae. Anterolateral portion of head with short spine. Masticatory margin of mandible almost invisible in full-face view. Dorsal surface of mandible on distal portion slightly wider than that on mandibular shaft. Spatulate or narrow spoon-shaped seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna about 1.0–1.2× length of pedicel. Setae on pronotum distinctly spatulate, widened distally with sharp or blunt apex. Metapleural gland bulla moderately developed, not expanding to dorsum of propodeal spiracle, so that propodeal declivity in lateral view weakly convex and rounded posteriorly on its ventral 1/3. Petiole relatively long in dorsal view, about 1.0–0.8× length of abdominal segment III.
Body color blackish to reddish brown.
unknown.

Workers of voeltzkowi species group in lateral view. A Mystrium eques sp. n. (CASENT0418314: holotype) B Mystrium janovitzi sp. n. (CASENT0482698: holotype) C Mystrium mirror sp. n. (CASENT0429897: holotype) D Mystrium oberthueri (CASENT0496066) E Mystrium shadow sp. n. (CASENT0318933: holotype) F Mystrium voeltzkowi (CASENT0002075).

Head of the workers of voeltzkowi species group in full-face view. A Mystrium eques sp. n. (CASENT0418314: holotype) B Mystrium janovitzi sp. n. (CASENT0482698: holotype) C Mystrium mirror sp. n. (CASENT0429897: holotype) D Mystrium oberthueri (CASENT0496066) E Mystrium shadow sp. n. (CASENT0318933: holotype) F Mystrium voeltzkowi (CASENT0002075).

Workers of voeltzkowi species group in dorsal view. A Mystrium eques sp. n. (CASENT0418314: holotype) B Mystrium janovitzi sp. n. (CASENT0482698: holotype) C Mystrium mirror sp. n. (CASENT0429897: holotype) D Mystrium oberthueri (CASENT0496066) E Mystrium shadow sp. n. (CASENT0318933: holotype) F Mystrium voeltzkowi (CASENT0002075).

Ergatoid queens of voeltzkowi species group in lateral view. A Mystrium eques sp. n. (CASENT0418317: paratype) B Mystrium janovitzi sp. n. (CASENT0482696) C Mystrium mirror sp. n. (CASENT0318940: paratype) D Mystrium oberthueri (CASENT0496064) E Mystrium shadow sp. n. (CASENT0077647: paratype) F Mystrium voeltzkowi (CASENT0002092).

Head of the ergatoid queens of voeltzkowi species group in full-face view. A Mystrium eques sp. n. (CASENT0418317: paratype) B Mystrium janovitzi sp. n. (CASENT0482696) C Mystrium mirror sp. n. (CASENT0318940: paratype) D Mystrium oberthueri (CASENT0496064) E Mystrium shadow sp. n. (CASENT0077647: paratype) F Mystrium voeltzkowi (CASENT0002092).

Ergatoid queens of voeltzkowi species group in dorsal view. A Mystrium eques sp. n. (CASENT0418317: paratype) B Mystrium janovitzi sp. n. (CASENT0482696) C Mystrium mirror sp. n. (CASENT0318940: paratype) D Mystrium oberthueri (CASENT0496064) E Mystrium shadow sp. n. (CASENT0077647: paratype) F Mystrium voeltzkowi (CASENT0002092).
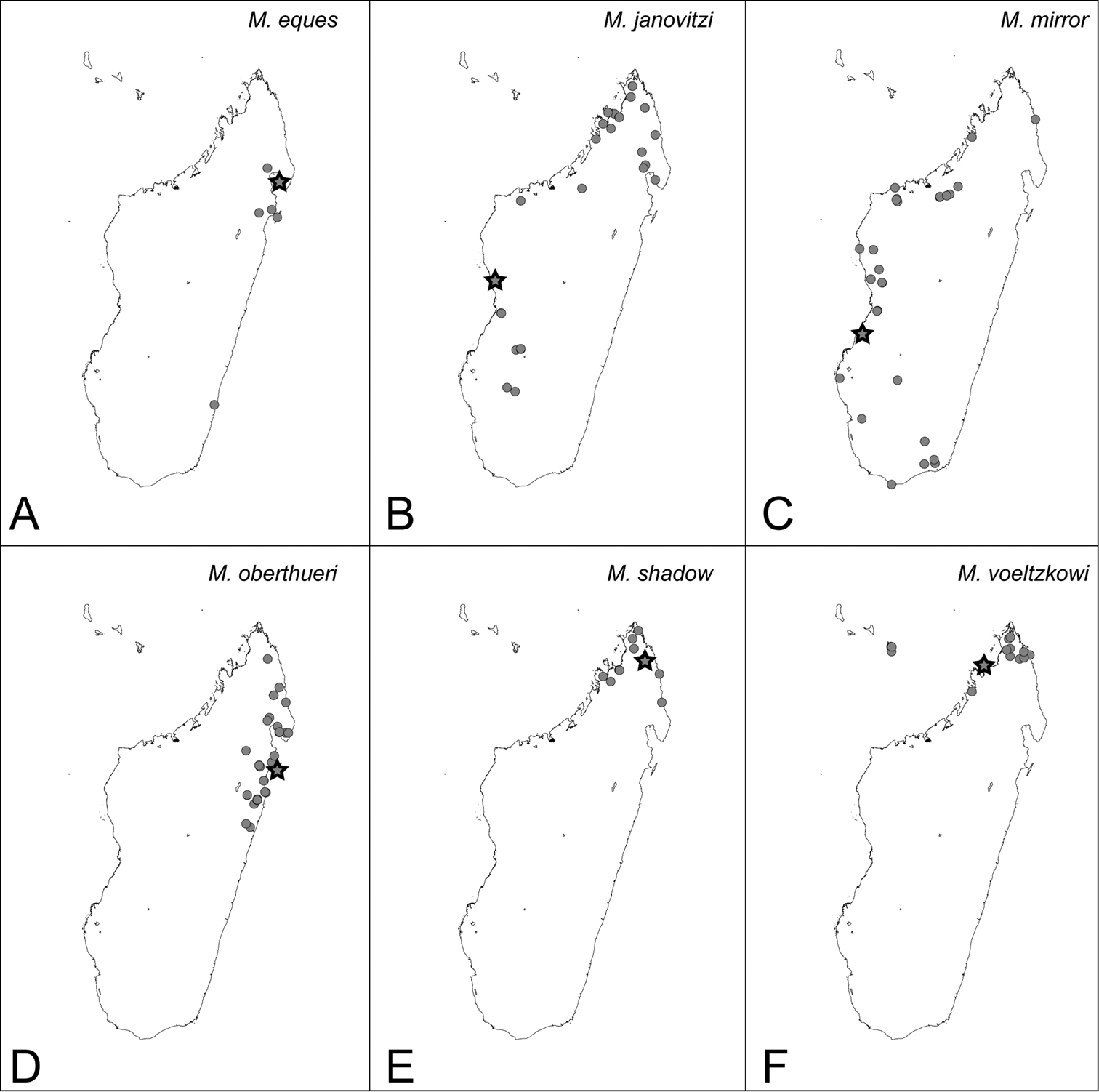
Distribution maps for voeltzkowi species group. A Mystrium eques sp. n. B Mystrium janovitzi sp. n. C Mystrium mirror sp. n. D Mystrium oberthueri E Mystrium shadow sp. n. F Mystrium voeltzkowi. Star symbols represent the type locality: the localities for Mystrium oberthueri and Mystrium voeltzkowi are approximate, which are assumed from the original descriptions.
This species name is the Latin word eques, inspired by the distinctly developed clypeus (shield) on the new species. The species epithet is a noun and invariant.
MADAGASCAR: as in Figure 56A.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Toamasina. Montagne d’Akirindro 7.6 km 341° NNW Ambinanitelo (-15.28833°, 49.54833°), rainforest, 600 m alt.; 5.3 km SSE Ambanizana, Andranobe (-15.66667°, 49.96667°), rainforest, 425 m alt.; Tampolo, Masoala Peninsula, 40.4 km 154° SSE Maroantsetra (-15.73°, 49.96°), rainforest, 30 m alt.; Res. Ambodiriana, 4.8 km 306° Manompana, along Manompana river (-16.67233°, 49.70117°), rainforest, 125 m alt.; Réserve Spéciale Ambatovaky, Sandrangato river (-16.77274°, 49.26551°), rainforest, 450 m alt.; Ile Sainte Marie, Forêt Kalalao, 9.9 km 34° Ambodifotatra (-16.9225°, 49.88733°), rainforest, 100 m alt.; Réserve Speciale Manombo 24.5 km 228° Farafangana (-23.01583°, 47.719°), rainforest, 30 m alt.
The worker of Mystrium eques is easily distinguished from those of other Mystrium species by a combination of the following characters: strong and deep longitudinal sculpture on the dorsal and lateral surface of pronotum (as in Fig. 17B); anteromedial portion of the clypeus strongly convex anteriorly (Fig. 18B); and sharper angle between the posterior and dorsal faces of the vertex on the median line (as in Fig. 16A). The queen of Mystrium eques is differentiated from other Mystrium queens by having vestigial wings reduced to small appendages (as in Fig. 25D) with undeveloped wing sclerites (ergatoid), posterior face of the vertex forming approximately a right angle with its dorsal face on the median line (Fig. 21A), and a strongly convex anterior clypeal margin (Fig. 22A).
The worker of Mystrium eques is similar to that of Mystrium oberthueri, however, the strongly convex anterior clypeal margin separates Mystrium eques (Fig. 18B) from Mystrium oberthueri (Fig. 18A), in which the anterior margin of the clypeus is straight. The male of Mystrium eques is not yet known. Because of the similarity between the workers and queens of Mystrium eques and Mystrium oberthueri, we assume the male of Mystrium eques is also similar to that of Mystrium oberthueri. The most easily distinguishable character between workers of Mystrium eques and Mystrium oberthueri, the anterior margin of the clypeus (Figs 18A, B), is unfortunately not useful for males because the anterior clypeal margin and the conical setae on it are less developed in males. Mystrium eques (Fig. 56A) has a sympatric distribution with Mystrium oberthueri (Fig. 56D), so collection locality does not separate these two similar species either. The current male specimens identified as Mystrium oberthueri may contain males of Mystrium eques. Further studies are necessary to resolve this issue.
http://zoobank.org/B390A568-7F20-45E0-95AF-74706CEAC3AB
http://species-id.net/wiki/Mystrium_janovitzi
Figs 4D, 5D, 17A, 17C, 23A, 23C, 26D, 28D, 31B, 32B, 32D, 37D, 38D, 39D, 40D, 41D, 50B, 51B, 52B, 53B, 54B, 55B, 56BWorker: CASENT0482698, BLF04508, MADAGASCAR, Mahajanga, Forêt de Tsimembo, 11.0 km 346° NNW Soatana (-18.99528°, 44.4435°), 50 m alt., 21–25.xi.2001, Fisher-Griswold Arthropod Team leg. [CASC].
Description. Measurements: holotype. HL 1.74, HW 1.89, SL 1.13, ML 1.69, HD 1.25, WL 2.15, PnW 1.2, PpW 0.99, PtW 1.07, PtL 0.73, CI 108.5, SI 59.8, MI 89.5, PpI 82.5, PtI 146.
HL 1.40–2.27, HW 1.46–2.67, SL 0.90–1.61, ML 1.30–2.58, HD 1.01–1.79, WL 1.83–2.94, PnW 0.91–1.52, PpW 0.80–1.28, PtW 0.82–1.32, PtL 0.56–0.90, CI 101.6–117.5, SI 59.7–67.9, MI 67.0–103.1, PpI 84.3–94.6, PtI 138.7–160.3 (10 specimens measured).
Posterolateral corner of head moderately expanding posteriorly. Posterior face of vertex forming almost right angle with dorsal face on median line of head, so that declivity of vertex on median part even steeper than on lateral part. Ventral half of vertex sculptured. Eye usually large, but sometimes small. Anterior margin of clypeus straight with small, extremely low conical setae. Genal tooth of head undeveloped, anterolateral corner of head angled. Masticatory margin of mandible almost invisible in full-face view, and width of dorsal surface of mandible almost identical from mandibular shaft to distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) about as long as pedicel (second antennal segment). Shallow, rough, and fragmented longitudinal reticulation irregularly impressed along the entire pronotal dorsum. Shallow, rough, and fragmented longitudinal reticulation irregularly impressed on lateral surface of pronotum. Mesonotum often not differentiated from propodeum in dorsal view, length as long as that of propodeum. Metanotal groove indistinct in lateral view. Metapleural gland bulla moderately developed, and propodeal declivity in lateral view almost straight though sometimes weakly rounded. Petiole relatively long (PtI 138.7–160.3), with slightly widened posterior 1/4 to 1/3 in dorsal view, anterior margin gently rounded or almost straight, and never edged by striae.
Body color black. Four distal segments of antennal club brighter.
Description. Measurements: HL 1.32–1.77, HW 1.37–2.04, SL 0.99–1.34, ML 1.31–1.80, HD 0.99–1.35, WL 1.86–2.43, PnW 0.88–1.92, PpW 0.83–1.08, PtW 0.81–1.19, PtL 0.58–0.69, CI 103.9–114.8, SI 65.7–74.5, MI 88.5–95.3, PpI 48.6–95.1, PtI 139.6–177.3 (10 specimens measured).
Wings vestigial and reduced to small appendages, which are sometimes indistinct. Wing sclerites undeveloped. Posterolateral corner of head moderately expanding posteriorly, expansion not differentiated from that of workers. Posterior face of vertex forming almost right or even slightly acute angle with dorsal face on median line of head, so that declivity of vertex on median part even steeper than on lateral part. Ventral half of vertex sculptured. Eye varied from well developed to small. Ocelli absent. Anterior margin of clypeus straight with extremely short and small conical setae. Genal tooth of head absent, corner usually unangled, sometimes angled into a small, short spine. Masticatory margin of mandible almost invisible in full-face view, and dorsal surface on distal portion as wide as that on mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna as long as pedicel. Setae on pronotum usually narrowing distally, or sometimes widened distally, with strongly sharpened apex. Metapleural gland bulla less developed, not expanding to either dorsum of propodeal spiracle or propodeal declivity margin in lateral view, so that posterior margin of propodeum in lateral view almost straight. Petiole relatively long in dorsal view, about 0.8-1.0× length of abdominal segment III.
Body color black.
Description. Measurements: HL 1.10–1.22, HW 1.52–1.72, SL 0.36–0.42, EL 0.62–0.71, WL 2.45–2.96, MnW 1.57–1.76, CI 135.2–141.0, SI 21.2–25.2, EI 54.1–58.1, MnI 99.3–103.9 (5 specimens measured).
Eye moderately large, occupying 0.6× head length. Ocelli reaching to dorsal margin of head in full-face view, or just failing to reach dorsal margin. Dorsal margin of head in full-face view rounded. Both anterior and lateral ocelli small. Lateral ocellus small and distant from eye: distance between these more than 2× maximum diameter of lateral ocellus. Posterior face of vertex rising steeply from neck, dorsal face slightly shorter than posterior face. Palpal formula 4, 3. First segment of maxillary palp flattened and distinctly wider than second segment. Second maxillary palpomere longer than third. Notauli deeply to shallowly impressed on mesoscutum, but often unclear. Petiole in dorsal view thin, length 0.8–0.65× that of abdominal tergite III. Petiolar dorsum covered with shallow, sparse, irregular punctures, or almost smooth. Abdominal tergum VIII without deep punctures, almost smooth.
Distal portion of abdominal sternum IX smooth and not punctured. Basal ring long, distinctly extending basally. Telomere distinctly extending farther distally than digitus. Basoventral expansion of aedeagus moderately developed basoventrally, longer than dorsal extension. Ventral margin of aedeagus gently curved ventrally in lateral view. Aedeagus moderately narrowing distally on distal half, distal portion widely rounded.
On forewing, cu-a located far basal from junction of Media (M) and Cubitus (Cu).
Body color reddish brown to black.
The specific epithet is the patronym of Dr. Tyler Janovitz in recognition of his interest in myrmecology and his support for research on ants.
MADAGASCAR: as in Figure 56B.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Antsiranana. Ambohitra (Joffreville) (-12.53333°, 49.16667°), rainforest, 1100 m alt.; Parc National Montagne d’Ambre, 3.6 km 235° SW Joffreville (-12.53444°, 49.1795°), montane rainforest, 925 m alt.; Rés. Ankarana, 7 km SE Matsaborimanga (-12.9°, 49.11667°), rainforst, 150 m alt.; Forêt de Binara, 9.1 km 233° SW Daraina (-13.26333°, 49.60333°), rainforest, 800 m alt.; Nosy Be, Réserve Naturelle Intégrale de Lokobe, 6.3 km 112° ESE Hellville (-13.41933°, 48.33117°), rainforest, 30 m alt.; Forêt Ambato, 26.6 km 33° Ambanja (-13.4645°, 48.55167°), rainforest, 150 m alt.; Galoko chain, Mont Galoko (-13.58487°, 48.71818°), rainforest, 520 m alt.; (-13.58745°, 48.71419°), rainforest, 380 m alt.; Ampasindava, Forêt d’Ambilanivy, 3.9 km 181° S Ambaliha (-13.79861°, 48.16167°), rainforest, 600 m alt.; R.S. Manongarivo, 10.8 km 229° SW Antanambao (-13.96167°, 48.43333°), rainforest, 400 m alt.; Makirovana forest (-14.17066°, 49.95409°), rainforest, 225 m alt.; Forêt d’Anabohazo, 21.6 km 247° WSW Maromandia (-14.30889°, 47.91433°), tropical dry forest, 120 m alt.; 6.5 km SSW Befingotra, Rés. Anjanaharibe-Sud (-14.75°, 49.5°), rainforest, 875 m alt. Toamasina. Montagne d’Anjanaharibe, 18.0 km 21° NNE Ambinanitelo (-15.18833°, 49.615°), rainforest, 470 m alt.; Montagne d’Akirindro 7.6 km 341° NNW Ambinanitelo (-15.28833°, 49.54833°), rainforest, 600 m alt.; 6.3 km S Ambanizana, Andranobe (-15.6813°, 49.958°), rainforest, 5 m alt. Mahajanga. Forêt Ambohimanga, 26.1 km 314° Mampikony (-15.96267°, 47.43817°), tropical dry forest, 250 m alt.; Parc National de Namoroka, 17.8 km 329° WNW Vilanandro (-16.37667°, 45.32667°), tropical dry forest, 100 m alt.; Forêt de Tsimembo, 11.0 km 346° NNW Soatana (-18.99528°, 44.4435°), tropical dry forest, 50 m alt.; 8.7 km 336° NNW Soatana (-19.02139°, 44.44067°), tropical dry forest, 20 m alt. Toliara. 48 km ENE Morondava, Kirindy (-20.06667°, 44.65°), tropical dry forest, 30 m alt.; Forêt de Kirindy, 15.5 km 64° ENE Marofandilia (-20.06915°, 44.66041667°), tropical dry forest, 30 m alt.; Makay Mts. (-21.22284°, 45.32477°), gallery forest on sandy soil, 490 m alt.; (-21.25864°, 45.16412°), gallery forest with bamboo, 500 m alt.; Vohibasia Forest, 59 km NE Sakaraha (-22.46667°, 44.85°), tropical dry forest, 780 m alt.; Fianarantsoa. Forêt d’Analalava, 29.6 km 280° W Ranohira (-22.59167°, 45.12833°), tropical dry forest, 700 m alt.
The worker of Mystrium janovitzi is distinguished easily from that of the other Mystrium species by the combination of the following characters: the labrum lacking central longitudinal carina (as in Fig. 8B); low clypeal conical setae (Fig. 17C); the posterior face of the vertex forming a right angle with its dorsal face (as in Fig. 16A); wide pronotum covered with fine sculpture that is neither longitudinal nor reticulate (Fig. 17A). This combination of diagnostic characters for the worker is also useful for distinguishing ergatoid queens of Mystrium janovitzi. The ergatoid queen of Mystrium janovitzi is similar to conspecific workers in its sculpture and body color, but differs in having a narrower mandible with a straight apical tooth (as in Fig. 15B) and distinct vestigial wings (as in Fig. 25D). For the male, small ocelli not strongly protruding from posterior margin of head in full-face view (Fig. 31B), rounded posterior margin of head in full-face view, smooth abdominal tergum VIII (Fig. 28B), and vertex strongly rising from neck (Fig. 32B) separate Mystrium janovitzi from other known Mystrium males in the Malagasy region.
http://zoobank.org/BFEBDF95-71C8-48ED-BDDD-3763E4EE7C0C
http://species-id.net/wiki/Mystrium_mirror
Figs 5C, 8D, 20A, 20C, 20E, 24B, 25A, 25C, 26A, 26C, 27B, 27D, 37E, 38E, 39E, 40E, 41E, 50C, 51C, 52C, 53C, 54C, 55C, 56CWorker: CASENT0429897, BLF04760, MADAGASCAR, Toliara, Parc National de Kirindy Mite, 16.3 km 127° SE Belo sur Mer (-20.79528°, 44.147°), 80 m alt., 6–10.xii.2001, Fisher-Griswold Arthropod Team [CASC].
5 workers: CASENT0429898 [CASC], CASENT0318935 [BMNH], CASENT0318936 [MHNG], CASENT0318937 [MCZC], CASENT0318938 [NHMB]; 3 ergatoid queens: CASENT0429899 [CASC], CASENT0318939 [BMNH], CASENT0318940 [MHNG]; 4 males: CASENT0429893 [CASC], CASENT0429895 [BMNH], CASENT0318941 [MHNG], CASENT0318942 [MCZC], with same data as holotype.
Description. Measurements: holotype. HL 1.60, HW 1.68, SL 1.19, ML 1.72, HD 1.03, WL 1.89, PnW 0.91, PpW 0.76, PtW 0.79, PtL 0.55, CI 104.7, SI 71.1, MI 102.7, PpI 83.2, PtI 143.8.
HL 1.16-1.91, HW 1.16–2.06, SL 0.94–1.48, ML 1.34–2.30, HD 0.84–1.33, WL 1.46–2.27, PnW 0.69–1.08, PpW 0.60–0.94, PtW 0.59–0.91, PtL 0.37–0.59, CI 99.5–108.3, SI 67.9–80.9, MI 103.8–115.7, PpI 81.5–92.6, PtI 143.7–163.2 (10 specimens measured).
Posterolateral corner of head strongly expanding posteriorly. Posterior face of vertex forming a blunt angle with its dorsal face on median line of head, so that declivity of vertex on lateral part distinctly steeper than on median part. Ventral half of vertex sculptured. Eye developed, relatively larger than that of Mystrium voeltzkowi. Anterior margin of clypeus straight to weakly convex with moderately long conical setae. Genal tooth of head weakly developed, reaching or slightly exceeding basal line of lateral lobe of clypeus. Masticatory surface of mandible in full-face view visible on basal half and invisible on distal half, width of dorsal surface of mandible almost identical from mandibular shaft to distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) about 1.0-1.3× length of pedicel (second antennal segment). Pronotal dorsum covered with strong and longitudinal striae, center deeply impressed. Shallow but thick longitudinal striae impressed on lateral surface of pronotum. Mesonotum differentiated from propodeum in dorsal view, length shorter than that of propodeum. Metanotal groove shallowly and gently impressed, mesonotum higher than pronotum in lateral view. Metapleural gland bulla moderately developed, propodeal declivity in lateral view almost straight. Petiole gently narrowing from anterior 1/3 in dorsal view, anterior margin straight to gently rounded and not edged by striae.
Body color reddish brown to dark brown. Four distal segments of antennal club brighter.
Description. Measurements: HL 1.17–1.83, HW 1.19–1.55, SL 0.91–1.24, ML 1.12–1.50, HD 0.83–1.08, WL 1.52–2.02, PnW 0.70–0.87, PpW 0.69–0.87, PtW 0.72–0.92, PtL 0.38–0.49, CI 77.6–106.7, SI 72.0–81.9, MI 87.7–104.0, PpI 94.0–106.6, PtI 159.5–194.5 (10 specimens measured).
Wings usually vestigial and reduced to quite small appendages; sometimes completely absent. Wing sclerites undeveloped. Posterolateral corner of head strongly to weakly expanding posteriorly, expansion relatively weaker than that of workers. Vertex usually thin, forming blunt angle between posterior and dorsal faces on median line of head, so that declivity of vertex on lateral part distinctly steeper than on median part. Ventral half of vertex sculptured, or not differentiated from dorsal region. Eye moderate and distinct. Ocelli absent. Anterior margin of clypeus straight to weakly convex with small conical setae. Genal tooth of head absent and not angled, or angled into small, short spine. Masticatory margin of mandible almost invisible in full-face view, and dorsal surface on distal portion as wide as that on mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellar segment on antenna moderately long, about 1.1-1.2× length of pedicel. Setae on pronotum almost simple, narrowing distally with strongly sharpened apex. Metapleural gland bulla moderately developed and not expanding dorsally to propodeal spiracle, so that propodeal declivity in lateral view weakly convex and rounded posteriorly on its ventral 1/3. Petiole relatively long in dorsal view, about 0.6-0.8× length of abdominal segment III.
Body color yellowish brown, brown or reddish brown.
Description. Measurements: HL 0.93–1.05, HW 1.31–1.56, SL 0.29–0.35, EL 0.67–0.78, WL 1.90–2.38, MnW 1.23–1.40, CI 140.6–149.3, SI 21.5–23.1, EI 70.2–74.3, MnI 88.6–94.0 (6 specimens measured).
Eye quite large, occupying about 0.75× of head length. Ocelli protruding from dorsal margin of head in full-face view. Dorsal margin of head in full-face view rounded. Both anterior and lateral ocelli large. Distance between lateral ocellus and eye equal to or shorter than diameter of lateral ocellus. Posterior face of vertex clearly differentiated from dorsal face, dorsal face distinctly shorter than posterior face. Palpal formula 4, 3. First segment of maxillary palp flattened and distinctly wider than second segment. Second maxillary palpomere longer than third. Notauli shallowly and weakly impressed on mesoscutum, but often unclear. Petiole in dorsal view thin, length 0.55–0.65× that of abdominal tergite III. Petiolar dorsum covered with fine punctures. Abdominal tergum VIII without deep punctures, almost smooth.
Distal portion of abdominal sternum IX smooth and not punctured. Basal ring short, not extending basally. Telomere distinctly extending distally farther than digitus. Basoventral expansion of aedeagus well developed basoventrally, distinctly longer than dorsal extension. Ventral margin of aedeagus almost straight in lateral view. Aedeagus distinctly narrowing distally, distal portion relatively sharp.
On forewing, cu-a located at junction of Media (M) and Cubitus (Cu), or slightly to far basal from junction.
Body color yellowish to reddish brown.
This species name is the English word mirror, inspired by the remarkable variation displayed in this species. This species might confuse an observer as a magic mirror would, which reflects different views to the observer. The species epithet is a noun and invariant.
MADAGASCAR: as in Figure 56C.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Antsiranana. Ambondrobe, 41.1 km 175° Vohemar (-13.71533°, 50.10167°), littoral rainforest, 10 m alt.; Forêt d’Anabohazo, 21.6 km 247° WSW Maromandia (-14.30889°, 47.91433°), tropical dry forest, 120 m alt.; Mahajanga. Forêt Ambohimanga, 26.1 km 314° Mampikony (-15.96267°, 47.43817°), tropical dry forest, 250 m alt.; Parc National de Baie de Baly, 12.4 km 337° NNW Soalala (-16.01°, 45.265°), tropical dry forest, 10 m alt.; Parc National d’Ankarafantsika, Forêt de Tsimaloto, 18.3 km 46° NE de Tsaramandroso (-16.22806°, 47.14361°), tropical dry forest, 135 m alt.; Ampijoroa Station Forestière, 5.4 km 331° NW Andranofasika (-16.29889°, 46.813°), tropical dry forest, 70 m alt.; Ampijoroa Station Forestière, 40 km 306° NW Andranofasika (-16.32083°, 46.81067°), tropical dry forest, 130 m alt.; Réserve d’Ankoririka, 10.6 km 13° NE de Tsaramandroso (-16.26722°, 47.04861°), tropical dry forest, 210 m alt.; Parc National de Namoroka, 17.8 km 329° WNW Vilanandro (-16.37667°, 45.32667°), tropical dry forest, 100 m alt.; 16.9 km 317° NW Vilanandro (-16.40667°, 45.31°), tropical dry forest, 100 m alt.; 9.8 km 300° WNW Vilanandro (-16.46667°, 45.35°), tropical dry forest, 140 m alt.; Réserve forestière Beanka, 50.2 km E Maintirano (-18.02649°, 44.05051°), tropical dry forest on tsingy, 250 m alt.; 52.7 km E Maintirano (-18.0622°, 44.52587°), tropical dry forest on tsingy, 300 m alt.; Parc National Tsingy de Bemaraha, 10.6 km ESE 123° Antsalova (-18.70944°, 44.71817°), tropical dry forest on tsingy, 150 m alt.; 2.5 km 62° ENE Bekopaka, Ankidrodroa River (-19.13222°, 44.81467°), tropical dry forest on tsingy, 100 m alt.; 3.4 km 93° E Bekopaka, Tombeau Vazimba (-19.14194°, 44.828°), tropical dry forest, 50 m alt.; Forêt de Tsimembo, 8.7 km 336° NNW Soatana (-19.02139°, 44.44067°), tropical dry forest, 20 m alt.; Toliara. Forêt de Kirindy, 15.5 km 64° ENE Marofandilia (-20.045°, 44.66222°), tropical dry forest, 100 m alt.; 48 km ENE Morondava, Kirindy (-20.06667°, 44.65°), tropical dry forest, 30 m alt.; Forêt de Beroboka, 5.9 km 131° SE Ankidranoka (-22.23306°, 43.36633°), tropical dry forest, 80 m alt.; Forêt de Mite, 20.7 km 29° WNW Tongobory (-23.52417°, 44.12133°), gallery forest, 75 m alt.; Forêt Vohidava 88.9 km N Amboasary (-24.24067°, 46.28783°), spiny forest/dry forest transition, 500 m alt.; Parc National d’Andohahela, Forêt de Manantalinjo, 33.6 km 63° ENE Amboasary, 7.6 km 99° E Hazofotsy (-24.81694°, 46.61°), spiny forest/thicket, 150 m alt.; Forêt d’Ambohibory, 1.7 km 61° ENE Tsimelahy, 36.1 km 308° NW Tolagnaro (-24.93°, 46.6455°), tropical dry forest, 300 m alt.; Réserve Privé Berenty, Forêt de Bealoka, Mandraré River, 14.6 km 329° NNW Amboasary (-24.95694°, 46.2715°), gallery forest, 35 m alt.; Réserve Spéciale de Cap Sainte Marie, 14.9 km 261° W Marovato (-25.59444°, 45.14683°), spiny forest/thicket, 160 m alt. Fianarantsoa. Parc National d’Isalo, Ambovo Springs, 29.3 km 4° N Ranohira (-22.29833°, 45.35167°), Uapaca woodland, 990 m alt.
The worker of Mystrium mirror is distinguished from workers of other Mystrium species by the combination of a central longitudinal furrow on the pronotal dorsum (as in Fig. 13B), reduced convexity of the anteromedial margin of the clypeus (Fig. 20A), blunter angle between the dorsal and posterior faces of the vertex on the median line of the head (as in Fig. 16B), and the straight posterior declivity of the propodeum (Fig. 20E). The workers of Mystrium mirror are quite similar to those of Mystrium shadow and Mystrium voeltzkowi, and the differences among the three species in workers are even slighter in small-sized individuals. When the comparison is made in workers within the same body size range, Mystrium mirror can be distinguished from Mystrium shadow by the smaller central clypeal conical setae (as in Fig. 19B), and from Mystrium voeltzkowi by either the straight declivity of the propodeum (Fig. 20E) and/or larger compound eye (Fig. 20C). The queen of Mystrium mirror can be distinguished from other Mystrium queens by a combination of the mesosoma without developed wing sclerites (ergatoid), posterior face of the vertex not strongly differentiated from its dorsal face (as in Fig. 21B), simpler body setae (Fig. 25A), a metapleural gland bulla that is moderately developed and not expanding dorsally to the propodeal spiracle (Fig. 24B), and brighter body color. The strength of the body sculpture is remarkably variable even among members in a single colony; some individuals may lack the striae and have a shiny pronotal dorsum - this weakness or lack of striae could prove to be an additional diagnostic character to separate this species from others. The ergatoid queens that are the most similar to Mystrium mirror are those of Mystrium voeltzkowi and Mystrium shadow. The less developed metapleural gland bulla (Fig. 24B) separates Mystrium mirror from Mystrium voeltzkowi, and pronotal setae simply narrowing distally with a sharp apex (Fig. 25A) separate it from Mystrium shadow. For the males, a large eye occupying almost 75% of the lateral margin of the head in full-face view (Fig. 26C), large lateral ocelli protruding from the dorsal margin of the head in full-face view (Fig. 38E), and a shorter distance between the eye and lateral ocellus (Fig. 26A) separate Mystrium mirror and Mystrium voeltzkowi from Mystrium shadow. However, only genital characters can distinguish Mystrium mirror from Mystrium voeltzkowi (Fig. 27).
The distribution range of Mystrium mirror (Fig. 56C) is allopatric with that of Mystrium voeltzkowi (Fig. 56F), but Mystrium mirror has a wider distribution than Mystrium voeltzkowi. Most of the specimens of Mystrium mirror were collected from tropical dry forests throughout most of western Madagascar (a few were collected in northern Madagascar), while most Mystrium voeltzkowi were collected from rainforest in the northern part of the country. This clear separation in distribution range and preferred habitat may be useful to roughly distinguish Mystrium mirror from Mystrium voeltzkowi.
We confirm that Mystrium mirror was previously used as material in one phylogenetic study (
http://species-id.net/wiki/Mystrium_oberthueri
Figs 1A, 16A, 17B, 17D, 18A, 18C, 18E, 22B, 23B, 23D, 30A, 37F, 38F, 39F, 40F, 41F, 50D, 51D, 52D, 53D, 54D, 55D, 56D[here designated]. Worker: CASENT0101993, MADAGASCAR, Insel Ste. Marie (Island of Sainte Marie), Perrot [MHNG: examined].
Description. Measurements: lectotype. HL 2.40, HW 2.61, SL 1.80, ML 2.92, HD 1.67, WL 2.79, PnW 1.39, PpW 1.21, PtW 1.18, PtL 0.81, CI 108.7, SI 69.0, MI 111.8, PpI 87.4, PtI 144.9.
HL 2.00–2.58, HW 2.12–2.76, SL 1.55–1.96, ML 2.39–3.17, HD 1.33–1.69, WL 2.43–2.96, PnW 1.17–1.38, PpW 1.02–1.29, PtW 1.00–1.32, PtL 0.67–0.81, CI 104.4–108.5, SI 68.8–74.9, MI 106.9–122.9, PpI 87.2–93.8, PtI 139.5–163.2 (10 specimens measured).
Posterolateral corner of head moderately expanding posteriorly. Posterior face of vertex forming almost right angle with dorsal face on median line of head, so that declivity of vertex on lateral part as steep as on median part. Whole region of vertex finely striated. Eye moderately small. Anterior margin of clypeus straight or weakly convex with long conical setae. Genal tooth of head relatively long, as long as lateral lobe of clypeus. Masticatory surface of mandible almost invisible in full-face view, and width of dorsal surface of mandible almost identical from mandibular shaft to distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) about 1.2–1.5× length of pedicel (second antennal segment). Strong, deep longitudinal striae regularly impressed on whole central part of pronotal dorsum. Strong, deep longitudinal striae impressed on lateral surface of pronotum. Mesonotum differentiated from propodeum in dorsal view, length slightly shorter than that of propodeum. Metanotal groove shallowly and gently impressed in lateral view. Metapleural gland bulla moderately developed, and propodeal declivity in lateral view almost straight. Petiole widened on posterior 1/3 and gently narrowing anteriorly in dorsal view, anterior margin straight to gently rounded and often edged by thick striae.
Body color black.
Description. Measurements: HL 1.40–1.93, HW 1.46–1.96, SL 1.07–1.47, ML 1.49–2.00, HD 1.00–1.35, WL 1.92–2.56, PnW 0.83–1.11, PpW 0.85–1.13, PtW 0.81–1.07, PtL 0.53–0.70, CI 101.6–108.6, SI 71.6–76.7, MI 96.3–108.8, PpI 96.0–104.4, PtI 149.1–177.1 (10 specimens measured).
Wings vestigial and reduced to small but distinct appendages. Wing sclerites undeveloped. Posterolateral corner of head moderately expanding posteriorly, expansion not differentiated from that in workers. Posterior face of vertex forming almost a right angle with dorsal face on median line of head, so that declivity of vertex on lateral part as steep as on median part. Ventral half of vertex sculptured. Eye small but distinct. Ocelli absent. Anterior margin of clypeus almost straight with long to moderate conical setae. Anterolateral portion of head with short spine. Masticatory margin of mandible almost invisible in full-face view, and dorsal surface on distal portion as wide as that on mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellomere (third antennal segment) long, about 1.2-1.5× length of pedicel (second antennal segment). Setae on pronotum distinctly spatulate, widened distally with sharp or blunt apex. Metapleural gland bulla moderately developed, not expanding dorsally to propodeal spiracle, so that propodeal declivity in lateral view weakly convex and rounded posteriorly on its ventral 1/3. Petiole relatively long in dorsal view, about 0.8× length of abdominal segment III.
Body color blackish to reddish brown.
Description. Measurements: HL 1.44, HW 2.09, SL 0.62, EL 0.95, WL 3.55, MnW 1.94, CI 145.1, SI 29.6, EI 66.2, MnI 92.9 (1 specimen measured).
Eye moderately large, occupying 0.6× of head length. Ocelli relatively distant from dorsal margin of head in full-face view. Dorsal margin of head in full-face view straight. Both anterior and lateral ocelli small. Lateral ocellus small and distant from eye: distance between these more than 1.5× maximum diameter of lateral ocellus. Posterior face of vertex not clearly differentiated from dorsal face, so that vertex almost continuously rounded. Palpal formula 4, 3. First maxillary palpomere flattened and distinctly wider than second segment. Second maxillary palpomere longer than third. Notauli clearly impressed on mesoscutum. Petiole in dorsal view thin, 0.8× as long as abdominal tergite III. Petiolar dorsum covered with shallow, irregular punctures. Abdominal tergum VIII without deep punctures, almost smooth.
Distal portion of abdominal sternum IX smooth and not punctured. Basal ring moderately long, expanding basoventrally. Telomere distinctly extending distally farther than digitus. Basoventral expansion of aedeagus moderately developed basoventrally, longer than dorsal extension. Ventral margin of aedeagus gently convex in lateral view. Aedeagus moderately narrowing distally and distal portion rounded.
On forewing, cu-a located far basal from junction of Media (M) and Cubitus (Cu).
Body color reddish brown to black.
MADAGASCAR: as in Figure 56D.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Antsiranana. Forêt d’ Antsahabe, 11.4 km 275° W Daraina (-13.21167°, 49.55667°), tropical dry forest, 550 m alt.; Makirovana forest (-14.17066°, 49.95409°), rainforest, 415 m alt.; Parc National de Marojejy, Manantenina River, 27.6 km 35° NE Andapa, 9.6 km 327° NNW Manantenina (-14.435°, 49.76°), rainforest, 775 m alt.; 28.0 km 38° NE Andapa, 8.2 km 333° NNW Manantenina (-14.43667°, 49.775°), rainforest, 450 m alt.; Marojejy R.N.I. #12 (-14.43583°, 49.76056°), rainforest, 610 m alt.; Forêt Ambanitaza, 26.1 km 347° Antalaha (-14.67933°, 50.18367°), rainforest, 240 m alt.; 1 km W Andampibe, Cap Masoala (-15.69361°, 50.18139°), lowland rainforest, 125 m alt.; Fotodriana, Cap Masoala (-15.69694°, 50.27028°), lowland rainforest, 25 m alt.; Toamasina. Montagne d’Anjanaharibe, 18.0 km 21° NNE Ambinanitelo (-15.18833°, 49.615°), rainforest, 470 m alt.; Montagne d’Akirindro 7.6 km 341° NNW Ambinanitelo (-15.28833°, 49.54833°), rainforest, 600 m alt.; 19 km ESE Maroantsetra (-15.48333°, 49.9°), rainforest, 350 m alt.; 5.3 km SSE Ambanizana, Andranobe (-15.66667°, 49.96667°), rainforest, 425 m alt.; 6.3 km S Ambanizana, Andranobe (-15.6813°, 49.958°), rainforest, 5 m alt.; 6.3 km S Ambanizana, Andranobe (-15.68131°, 49.958°), rainforest, 25 m alt.; Parc National Mananara-Nord, 7.1 km 261° Antanambe (-16.455°, 49.7875°), rainforest, 225 m alt.; Res. Ambodiriana, 4.8 km 306° Manompana, along Manompana river (-16.67233°, 49.70117°), rainforest, 125 m alt.; Réserve Spéciale Ambatovaky, Sandrangato river (-16.7633°, 49.26692°), rainforest, 520 m alt.; (-16.77274°, 49.26551°), rainforest, 450 m alt.; (-16.7755°, 49.26427°), rainforest, 430 m alt.; (-16.81745°, 49.2925°), rainforest, 400 m alt.; (-16.81753°, 49.29498°), rainforest, 360 m alt.; Ile Sainte Marie, Forêt Kalalao, 9.9 km 34° Ambodifotatra (-16.9225°, 49.88733°), rainforest, 100 m alt.; Réserve Forestière Tampolo, 95.2 km N Toamasina (-17.27808°, 49.42853°), littoral rainforest, 20 m alt.; S.F. Tampolo, 10 km NNE Fenoarivo Atn. (-17.2825°, 49.43°), littoral rainforest, 10 m alt.; Bridge at Onibi, NW of Mahavelona (-17.65°, 49.46667°), in clay forest; Mahavelona (Foulpointe) (-17.66667°, 49.5°), Forest; Parc National de Zahamena, Besaky River (-17.75244°, 48.85321°), rainforest, 760 m alt.; Sahavorondrano River (-17.75257°, 48.85725°), rainforest, 765 m alt.; Onibe River (-17.75908°, 48.85468°), rainforest, 780 m alt.; Reserve Betampona, Camp Vohitsivalana, 37.1 km 338° Toamasina (-17.88667°, 49.2025°), rainforest, 520 m alt.; Réserve Naturelle Betampona, 34.1 km 332° Toamasina (-17.916135°, 49.20185°), rainforest, 550 m alt.; Réserve Nationale Intégrale Betampona, Betampona 35.1 km NW Toamasina (-17.91801°, 49.20074°), rainforest, 500 m alt.; Reserve Betampona, Camp Rendrirendry 34.1 km 332° Toamasina (-17.924°, 49.19967°), rainforest, 390 m alt.; F.C. Sandranantitra (-18.04833°, 49.09167°), rainforest, 450 m alt.; F.C. Andriantantely (-18.695°, 48.81333°), rainforest, 530 m alt.; Sahafina forest 11.4 km W Brickaville (-18.81445°, 48.96205°), rainforest, 140 m alt. Mahajanga. Réserve Spéciale Marotandrano, Marotandrano 48.3 km S Mandritsara (-16.28322°, 48.81443°), transition humid forest, 865 m alt.
The worker of Mystrium oberthueri is easily distinguished from that of other Mystrium species by the combination of the following characters: strong, deep, and thick longitudinal sculpture on the dorsal and lateral surface of pronotum (Fig. 17B); straight anteromedial margin of the clypeus (Fig. 18A); a sharper angle between the dorsal and posterior faces of the vertex on the median line of the head (Fig. 16A). The queen of Mystrium oberthueri is differentiated from other queens of Mystrium species by the vestigial wings reduced to small appendages with undeveloped wing sclerites (ergatoid: as in Fig. 25D), the posterior face of the vertex forming approximately a right angle with its dorsal face on the median line of the head (as in Fig. 21A), straight anterior clypeal margin (Fig. 22B), and moderately developed clypeal conical setae (Fig. 23B). The male of Mystrium oberthueri can be separated from the other known Mystrium males by its smooth abdominal tergum VIII (as in Fig. 28), small lateral ocelli situated distant from posterior margin of head in full-face view, and straight dorsal margin of head in full-face view (Fig. 30A). We should note that the current male character diagnoses may not be able to distinguish between Mystrium oberthueri and Mystrium eques, which is not yet known from the male caste and may be quite similar to Mystrium oberthueri.
The lectotype for Mystrium oberthueri is designated. According to
Although worker descriptions for Mystrium oberthueri are reliable, male descriptions of this species in previous papers are problematic. The original description by
We describe a new species, Mystrium eques, with a worker similar to that of Mystrium oberthueri. As we have already mentioned in the section of Mystrium eques, the unknown male of Mystrium eques is likely to be quite similar to that of Mystrium oberthueri, and the current male diagnosis for Mystrium oberthueri may be insufficient to distinguish these two males (see Mystrium eques).
Material identified as Mystrium oberthueri in
http://zoobank.org/2CF2C722-970D-4829-87ED-C58CBF036481
http://species-id.net/wiki/Mystrium_shadow
Figs 19A, 21B, 25B, 25D, 28D, 30B, 31D, 32A, 32C, 37G, 38G, 39G, 40G, 41G, 50E, 51E, 52E, 53E, 54E, 55E, 56EWorker: CASENT0318933, BLF09661, MADAGASCAR, Antsiranana, Forêt de Binara, 9.1 km 233° SW Daraina (-13.26333°, 49.60333°), 650-800 m alt., 3.xii.2003, B.L. Fisher leg. [CASC].
4 workers: CASENT0077646 [BMNH], CASENT0077648 [MHNG], CASENT0318920 [MCZC], CASENT0318931 [NHMB]; 3 ergatoid queens: CASENT0077647 [CASC], CASENT0318932 [BMNH], CASENT0318934 [MHNG], with same data as holotype.
Description. Measurements: holotype. HL 2.11, HW 2.15, SL 1.60, ML 2.52, HD 1.43, WL 2.42, PnW 1.09, PpW 0.99, PtW 0.90, PtL 0.59, CI 101.9, SI 74.4, MI 116.9, PpI 91.1, PtI 152.6.
HL 1.36–2.25, HW 1.37–2.36, SL 1.00–1.78, ML 1.40–2.89, HD 0.90–1.49, WL 1.55–2.67, PnW 0.74–1.16, PpW 0.67–1.03, PtW 0.72–1.04, PtL 0.40–0.66, CI 97.0–104.9, SI 70.5–76.8, MI 102.3–125.5, PpI 85.6–93.6, PtI 147.3–179.4 (10 specimens measured).
Posterolateral corner of head strongly expanding posteriorly. Posterior face of vertex forming blunt angle with dorsal face on median line of head, so that declivity of vertex on lateral part distinctly steeper than on median part. Ventral half of vertex sculptured. Eye moderately small. Anterior margin of clypeus strongly convex with long conical setae. Genal tooth of head moderately developed, reaching to about half of lateral lobe of clypeus. Masticatory surface of mandible in full-face view slightly visible on basal half and invisible on distal half, width of dorsal surface of mandible almost identical from mandibular shaft to distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) about 1.0-1.3× length of pedicel (second antennal segment). Pronotal dorsum covered with strong longitudinal striae, which are waved and irregular, but median stria always deeply, straightly and clearly impressed. Strong, deep, and irregular longitudinal striae impressed on lateral surface of pronotum. Mesonotum differentiated from propodeum in dorsal view, length shorter than that of propodeum. Metanotal groove shallowly and gently impressed, mesonotum higher than pronotum in lateral view. Metapleural gland bulla moderately developed, and propodeal declivity in lateral view almost straight but sometimes weakly rounded. Petiole relatively short (PtI ), gently narrowing from anterior 1/3 in dorsal view, anterior margin straight to gently rounded and not edged by striae.
Body color reddish brown to black. Four distal segments of antennal club brighter.
Description. Measurements: HL 1.30–1.67, HW 1.31–1.70, SL 1.00–1.42, ML 1.30–1.81, HD 0.93–1.20, WL 1.74–2.33, PnW 0.76–1.00, PpW 0.76–0.98, PtW 0.76–0.96, PtL 0.45–0.61, CI 98.7–105.3, SI 76.2–85.5, MI 97.1–108.2, PpI 95.5–101.7, PtI 157.7–178.6 (10 specimens measured).
Wings vestigial and reduced to small but distinct appendages. Wing sclerites undeveloped. Posterolateral corner of head weakly expanding posteriorly, expansion distinctly weaker than that in workers. Posterior face of vertex forming blunt angle with dorsal face on median line of head, so that declivity of vertex on lateral part distinctly steeper than on median part. Ventral half of vertex sculptured. Eye small but distinct. Ocelli absent. Anterior margin of clypeus widely convex with short conical setae, sometimes with median notch. Genal tooth of head with short spine. Masticatory margin of mandible almost invisible in full-face view, dorsal surface on distal portion as wide as that on mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellomere (third antennal segment) moderately long, about 1.5–1.2× length of pedicel (second antennal segment). Setae on pronotum widened distally with sharp apex. Metapleural gland bulla moderately developed, not expanding to dorsum of propodeal spiracle nor propodeal declivity margin in lateral view, so that posterior margin of propodeum in lateral view almost straight. Petiole relatively long in dorsal view, about 0.5–0.8× length of abdominal segment III.
Body color reddish to blackish brown.
Description. Measurements: HL 1.12–1.18, HW 1.53–1.54, SL 0.41–0.49, EL 0.56–0.60, WL 2.32–2.69, MnW 1.44–1.53, CI 130.6–136.6, SI 26.8–31.8, EI 47.4–53.6, MnI 93.9–99.7 (2 specimens measured).
Eye moderately large, occupying 0.6× head length. Ocelli relatively distant from dorsal margin of head in full-face view. Dorsal margin of head in full-face view weakly rounded. Both anterior and lateral ocelli small. Lateral ocellus relatively small and distant from eye: distance between these more than 2.5× maximum diameter of lateral ocellus. Posterior face of vertex not clearly differentiated from dorsal face, so that margins almost continuously rounded. Palpal formula 4, 3. First maxillary palpomere flattened and distinctly wider than second. Second maxillary palpomere longer than third. Notauli shallowly and weakly impressed on mesoscutum, but often unclear. Petiole in dorsal view thin, length 0.6× that of abdominal tergite III. Petiolar dorsum covered with shallow, irregular punctures. Abdominal tergum VIII without deep punctures, almost smooth.
Distal portion of abdominal sternum IX smooth and not punctured. Basal ring moderately long, gently expanding basally. Telomere distinctly extending distally farther than digitus. Basoventral expansion of aedeagus less developed basoventrally, as long as dorsal extension. Ventral margin of aedeagus almost straight in lateral view. Aedeagus distinctly narrowing distally on distal half, but distal portion rounded.
On forewing, cu-a located far basal from junction of Media (M) and Cubitus (Cu).
Body color reddish brown to black.
This species name is the English word shadow, inspired by the difficulty of recognizing this new species, which is distributed like Mystrium voeltzkowi but hidden in its shadow. The species epithet is a noun and thus invariant.
MADAGASCAR: as in Figure 56E.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Antsiranana. Forêt d’Orangea, 3.6 km 128° SE Remena (-12.25889°, 49.37467°), littoral rainforest, 90 m alt.; Parc National Montagne d’Ambre, 3.6 km 235° SW Joffreville (-12.53444°, 49.1795°), montane rainforest, 925 m alt.; Réserve Spéciale de l’Ankarana, 13.6 km 192° SSW Anivorano Nord (-12.86361°, 49.22583°), tropical dry forest, 210 m alt.; Forêt d’ Antsahabe, 11.4 km 275° W Daraina (-13.21167°, 49.55667°), tropical dry forest, 550 m alt.; Forêt de Binara, 9.1 km 233° SW Daraina (-13.26333°, 49.60333°), rainforest, 800 m alt.; 9.4 km 235° SW Daraina (-13.26333°, 49.6°), montane rainforest, 1100 m alt.; Galoko chain, Mont Galoko (-13.58487°, 48.71818°), rainforest, 520 m alt.; (-13.58745°, 48.71419°), rainforest, 380 m alt.; (-13.59358°, 48.73157°), montane forest, 1100 m alt.; Ambondrobe, 41.1 km 175° Vohemar (-13.71533°, 50.10167°), littoral rainforest, 10 m alt.; Ampasindava, Forêt d’Ambilanivy, 3.9 km 181° S Ambaliha (-13.79861°, 48.16167°), rainforest, 600 m alt.; R.S. Manongarivo, 12.8 km 228° SW Antanambao (-13.97667°, 48.42333°), rainforest, 780 m alt.; Forêt Ambanitaza, 26.1 km 347° Antalaha (-14.67933°, 50.18367°), rainforest, 240 m alt.
The worker of Mystrium shadow is distinguished from other Mystrium species by the combination of the following characters: a central longitudinal furrow on the pronotal dorsum (as in Fig. 13B); projected anteromedial portion of the clypeus (Fig. 19A); blunter angle between the dorsal and posterior faces of the vertex on the median line of the head (as in Fig. 16B); the straight posterior declivity of the propodeum (as in Fig. 20). The workers of Mystrium shadow are quite similar to those of Mystrium mirror and Mystrium voeltzkowi, and the differences among the workers of the three species are even slighter in small-sized individuals. When workers in the same body size range are compared, Mystrium shadow can be distinguished from Mystrium mirror by the central pair of conical setae that is larger in size than the adjacent pair (Fig. 19A), and from Mystrium voeltzkowi by a straight declivity of the propodeum (as in Fig. 20). The queen of Mystrium shadow can be distinguished from the other Mystrium queens by a combination of the mesosoma without developed wing sclerites but with vestigial wing appendages (ergatoid: Fig. 25D), posterior face of the vertex forming a blunt angle with its dorsal face on the median line of the head (Fig. 21B), spatulate setae on dorsum of head and pronotum (Fig. 25B), and the metapleural gland bulla that is moderately developed and not expanding dorsally to the propodeal spiracle (as in Fig. 24B). The ergatoid queens similar to those of Mystrium shadow are those of Mystrium voeltzkowi and Mystrium mirror; however, a less-developed metapleural gland bulla distinguishes Mystrium shadow from Mystrium voeltzkowi (as in Fig. 24), and spatulate setae on the pronotal dorsum (Fig. 25B) and the presence of the genal tooth of the head (Fig. 54E) distinguish Mystrium shadow from Mystrium mirror. The male of Mystrium shadow is not that similar to that of Mystrium voeltzkowi or Mystrium mirror. The male of Mystrium shadow can be easily separated from the latter two males by the small ocelli (as in Fig. 26B) and small eye (as in Fig. 26D). On the other hand, the male of Mystrium shadow is similar to that of Mystrium janovitzi. The shape of the vertex is the character that separates these two similar males: the posterior face of the vertex in posterior view is not clearly differentiated from its dorsal face in Mystrium shadow (Fig. 32A), while the posterior face of the vertex is clearly divided from its dorsal face on the median line of the head in Mystrium janovitzi (Fig. 32B).
This taxonomic revision permits us to update identifications in a recent ecological publication
http://species-id.net/wiki/Mystrium_voeltzkowi
Figs 1B, 4B, 5A, 11B, 13B, 13D, 15, 16B, 19B, 20B, 20D, 20F, 24A, 27A, 27C, 37H, 38H, 39H, 40H, 41H, 50F, 51F, 52F, 53F, 54F, 55F, 56F[here designated]. Worker: CASENT0101952 (the specimen on the bottom side with open mandible out of the two workers on the same pin), MADAGASCAR, Nossi-Bé (Nosy Be), Voeltzkow [MHNG: examined].
[here designated]. Ergatoid queen: CASENT0101995, MADAGASCAR, Nossi-Bé (Nosy Be), Voeltzkow [MHNG: same data with lectotype of Mystrium voeltzkowi, examined].
Description. Measurements: lectotype. HL 2.08, HW 2.12, SL 1.63, ML 2.67, HD 1.35, WL 2.58, PnW 1.17, PpW 1.00, PtW 0.97, PtL 0.66, CI 102.0, SI 76.9, MI 125.8, PpI 84.9, PtI 147.6.
HL 1.45–2.15, HW 1.41–2.26, SL 1.05–1.65, ML 1.47–2.72, HD 0.96–1.46, WL 1.72–2.68, PnW 0.80–1.20, PpW 0.69–1.09, PtW 0.70–1.03, PtL 0.46–0.67, CI 96.6–104.7, SI 71.7–80.1, MI 104.0–125.7, PpI 86.2–91.0, PtI 141.7–164.6 (9 specimens measured).
Posterolateral corner of head strongly expanding posteriorly. Posterior face of vertex forming blunt angle with dorsal face on median line of the head, so that declivity of vertex on lateral part distinctly steeper than on median part. Ventral half of vertex sculptured. Eye moderately small. Anterior margin of clypeus straight to weakly convex with moderately long conical setae. Genal tooth of head moderately developed, reaching about half of lateral lobe of clypeus. Masticatory surface of mandible in full-face view visible on basal half and invisible on distal half, width of dorsal surface of mandible almost identical from mandibular shaft to distal portion. Second maxillary palpomere longer than third. First flagellomere (third antennal segment) about 1.0–1.3× length of pedicel (second antennal segment). Pronotal dorsum covered with strong to weak longitudinal striae often waved and irregular, but median stria always deeply, straightly and clearly impressed. Shallow, fine longitudinal striae irregularly impressed on lateral surface of pronotum. Mesonotum not differentiated from propodeum in dorsal view in small individuals but differentiated in large individuals, its length shorter than that of propodeum. Metanotal groove indistinct in small individuals and shallowly and gently impressed in large individuals; mesonotum higher than pronotum in lateral view. Metapleural gland bulla strongly developed, so that propodeal declivity in lateral view convex posteriorly on ventral side. Petiole widened on posterior 1/4–1/2, gently narrowed anteriorly in dorsal view, anterior margin straight to gently rounded and not edged by striae.
Body color reddish brown to black. Four distal segments of antennal club brighter.
Description. Measurements: HL 1.18-1.59, HW 1.12-1.63, SL 0.88-1.35, ML 1.13-1.64, HD 0.82-1.13, WL 1.56-2.34, PnW 0.70-0.99, PpW 0.70-1.14, PtW 0.68-0.95, PtL 0.38-0.57, CI 95.2-106.4, SI 76.1-85.7, MI 93.1-108.7, PpI 97.1-115.9, PtI 161.8-186.9 (10 specimens measured).
Wings absent or vestigial and reduced to quite small appendages. Wing sclerites undeveloped. Posterolateral corner of head gently expanding posteriorly, expansion distinctly weaker than that in workers. Vertex usually thin, forming blunt angle between dorsal and posterior faces on median line of head, so that declivity of vertex on lateral part distinctly steeper than median part. Ventral half of vertex sculptured, not differentiated from dorsal region. Eye small but distinct. Ocelli absent. Anterior margin of clypeus straight to weakly convex with short conical setae. Genal tooth of head absent, usually even not angular, but angular in smallest individuals. Masticatory margin of mandible almost invisible in full-face view, dorsal surface on distal portion as wide as that on mandibular shaft. Spatulate seta present on basal side of each basal denticle on masticatory margin of mandible. First flagellomere (third antennal segment) moderately long, about 1.1–1.2× length of pedicel (second antennal segment). Setae on pronotum almost simple, narrowing distally with strongly sharpened apex. Metapleural gland bulla extremely developed and expanding dorsally to propodeal spiracle, so that propodeal declivity in lateral view strongly convex posteriorly on ventral 2/3. Petiole relatively long in dorsal view, about 0.5–0.8× length of abdominal segment III.
Body color brown to reddish brown.
Description. Measurements: HL 1.27, HW 1.87, SL 0.42, EL 0.91, WL 2.97, MnW 1.72, CI 146.9, SI 22.2, EI 71.4, MnI 92.0 (1 specimen from original type series measured).
Eye quite large, occupying about 0.75× head length. Ocelli protruding from dorsal margin of head in full-face view. Dorsal margin of head in full-face view rounded. Both anterior and lateral ocelli large. Distance between lateral ocellus and eye equal to or shorter than diameter of lateral ocellus. Posterior half of vertex clearly differentiated from dorsal half, dorsal face distinctly shorter than posterior face. Palpal formula 4, 3. First maxillary palpomere flattened and distinctly wider than second segment. Second maxillary palpomere longer than third. Notauli shallowly and weakly impressed on mesoscutum, but often unclear. Petiole in dorsal view thin, length 0.55× that of abdominal tergite III. Petiolar dorsum covered with shallow, irregular punctures. Abdominal tergum VIII without deep punctures, almost smooth.
Distal portion of abdominal sternum IX smooth and not punctured. Basal ring moderately long, gently expanding basally. Telomere distinctly extending distally farther than digitus. Basoventral expansion of aedeagus well developed basoventrally, distinctly longer than dorsal extension. Ventral margin of the aedeagus gently curved ventrally in lateral view. Aedeagus distinctly narrowing distally on distal half and apex rounded.
On forewing, cu-a located far basal from junction of Media (M) and Cubitus (Cu).
Body color yellowish to reddish brown.
MADAGASCAR and MAYOTTE: as in Figure 56F.
In addition to the type material, specimens from the following localities were examined in this study: MADAGASCAR. Antsiranana. Réserve Spéciale d’Ambre, 3.5 km 235° SW Sakaramy (-12.46889°, 49.24217°), tropical dry forest, 325 m alt.; Parc National Montagne d’Ambre, 3.6 km 235° SW Joffreville (-12.53444°, 49.1795°), montane rainforest, 925 m alt.; Réserve Spéciale de l’Ankarana, 13.6 km 192° SSW Anivorano Nord (-12.86361°, 49.22583°), tropical dry forest, 210 m alt.; 22.9 km 224° SW Anivorano Nord (-12.90889°, 49.10983°), tropical dry forest, 80 m alt.; Forêt d’Ampondrabe, 26.3 km 10° NNE Daraina (-12.97°, 49.7°), tropical dry forest, 175 m alt.; Forêt d’Analabe, 30.0 km 72° ENE Daraina (-13.08333°, 49.90833°), littoral rainforest, 30 m alt.; Forêt d’ Andavakoera, 21.4 km 75° ENE Ambilobe; 4.6 km 356° N Betsiaka (-13.11833°, 49.23°), rainforest, 425 m alt.; Forêt de Bekaraoka, 6.8 km 60° ENE Daraina (-13.16667°, 49.71°), tropical dry forest, 150 m alt.; Forêt d’ Antsahabe, 11.4 km 275° W Daraina (-13.21167°, 49.55667°), tropical dry forest, 550 m alt.; Nosy Be, Réserve Naturelle Intégrale de Lokobe, 6.3 km 112° ESE Hellville (-13.41933°, 48.33117°), rainforest, 30 m alt.; Forêt d’Anabohazo, 21.6 km 247° WSW Maromandia (-14.30889°, 47.91433°), tropical dry forest, 120 m alt.
MAYOTTE. Mt. Combani (-12.8°, 45.13333°), forest litter, 420 m alt.; (-12.80486°, 45.15266°), rainforest, 460 m alt.; (-12.80632°, 45.15314°), rainforest, 370 m alt.; Dapani (-12.96279°, 45.15037°), rainforest, 135 m alt.
The worker of Mystrium voeltzkowi is distinguished from other Mystrium workers by the combination of the following characters: the vertex forming a blunt angle between posterior and dorsal faces on the median line of the head (Fig. 16B), small compound eye (Fig. 20D), the genal tooth of the head reaching to half of the lateral lobe of the clypeus (Fig. 20B), and a medial pair of conical setae on the clypeus that is smaller than adjacent pair (Fig. 19B). In addition, the characters of a central longitudinal furrow on the pronotal dorsum (Fig. 13B) but without deep, stout striae impressed on dorsal and lateral surface of pronotum, and the posterior declivity of the propodeum convex posteriorly on the ventral half (Fig. 20F), so that the margin is somewhat rounded in lateral view, are also useful. The workers of Mystrium mirror and Mystrium shadow are quite similar to those of Mystrium voeltzkowi, and the differences among the three species in workers are even slighter in small-sized individuals. When the comparison is made in workers within the same body size range, Mystrium voeltzkowi can be distinguished from Mystrium shadow by the small central clypeal conical setae (Fig. 19B), and from Mystrium mirror by either a rounded declivity of the propodeum (Fig. 20F) or smaller compound eye (Fig. 20D). The queen of Mystrium voeltzkowi is easily distinguished from the other Mystrium queens by the mesosoma without developed wing sclerites (ergatoid: as in Fig. 25C), the developed metanotal gland bulla expanding dorsally to the propodeal spiracle (Fig. 24A), and the brighter body color. Among males, only genital characters distinguish Mystrium voeltzkowi from Mystrium mirror (Fig. 27).
The male of Mystrium voeltzkowi was described with conspecific workers from the same colony in the original description (
Mystrium fallax Forel, 1897 is synonymized with Mystrium voeltzkowi in this study. Mystrium fallax was apparently described by a simple mistake. According to the original description of Mystrium voeltzkowi by
The distribution of Mystrium voeltzkowi is limited to Mayotte and the northern part of Madagascar (Fig. 56F). The collection localities of Mystrium mirror are in tropical dry forest, gallery forest, and spiny forest (Fig. 56C), while those of Mystrium voeltzkowi are in tropical dry forest, montane rainforest, littoral rainforest, and rainforest (Fig. 56F). The distinct development of the metapleural grand bulla observed in Mystrium voeltzkowi (Figs 20F, 24A) may be related to its nesting environment, which has higher moisture levels and requires stricter control of fungi.
The reference to Mystrium voeltzkowi in recent molecular phylogenetic studies (
This study was partially supported by the National Science Foundation under Grant No. DEB-0072713, DEB-0344731, and DEB-0842395. We would like to acknowledge Philip S. Ward and Stefan P. Cover, and Agnièle Touret-Alby for the loan of materials, Francisco Hita Garcia, Flávia Esteves, Georg Fischer, Mathieu Molet, Christian Peeters, Barry Bolton, Wojciech J. Pulawski, Robert Zuparko, Eli M. Sarnat, Donat Agosti, and Barry Lee Bressler for editorial suggestions. We are grateful to Alex L. Wild for providing images; Estella Ortega, Michele Esposito, Erin Prado, and Cerise Chen for creating montage images; Michele Esposito for help with database management; and Charles Griswold for the use of the compound microscopic imaging system. The fieldwork on which this study is based could not have been completed without the gracious support of the Malagasy people, and the Arthropod Inventory Team (Balsama Rajemison, Jean Claude Rakotonirina, Jean-Jacques Rafanomezantsoa, Chrislain Ranaivo, Coco Randriambololona, Hanitriniana Rasoazanamavo, Nicole Rasoamanana, Clavier Randrianandrasana, Valerie Rakotomalala, and Dimby Raharinjanahary).
A data sheet for all specimens of Mystrium examined
Authors: Masashi Yoshimura, Brian L. Fisher
Data type: occurrences
Explanation note: Comprehensive data for all Mystrium specimens examined including information of specimens ID, species name, sex and caste, collection data, and depository.
Copyright notice: This dataset is made available under the Open Database License (http://opendatacommons.org/licenses/odbl/1.0/). The Open Database License (ODbL) is a license agreement intended to allow users to freely share, modify, and use this Dataset while maintaining this same freedom for others, provided that the original source and author(s) are credited.