





(C) 2013 Hee-Wook Cho. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Citation: Cho H-W, Borowiec L (2013) A new species of the genus Ambrostoma Motschulsky (Coleoptera, Chrysomelidae, Chrysomelinae) from South Korea, with larval descriptions and biological notes. ZooKeys 321: 1–13. doi: 10.3897/zookeys.321.4972
Ambrostoma koreana sp. n. is described from South Korea. Detailed descriptions and illustrations of adult and all larval instars are provided and differences to closely related species are discussed. Ovoviviparity is reported for the first time in the genus Ambrostoma. Notes on distribution, host plant and biology are also provided.
Chrysomelidae, Ambrostoma, new species, larva, biology, ovoviviparity, Korea
The genus Ambrostoma was described by
In 2006, the first author collected unusual Ambrostoma specimens, adults and larvae, on Namhaedo Island in South Korea. After a detailed examination we concluded that they belong to a new species described below. We compared the larval morphology of Ambrostoma to those of Parambrostoma. Notes on distribution, host plant, biology and occurrence of ovoviviparity are also provided.
The type specimens were deposited in the Department of Biodiversity and Evolutionary Taxonomy, University of Wrocław, Poland (DBET) and H.-W. Cho’s private collection, Republic of Korea (HWC). Biological observations were made in April–May 2006, at the type locality and under laboratory conditions. Adults and larvae collected from the type locality were kept in plastic containers (10 cm diameter, 12 cm deep) with leaves of host plant and checked daily. All larval specimens used in the study were preserved in 70% ethanol. For morphological studies of minute structures, some larvae were dissected, cleared in 10% sodium hydroxide solution, rinsed in distilled water, and then mounted on slides with Swan’s liquid (20 g distilled water, 15 g gum arabic, 60 g chlorhydrate, 3 g glucose, and 2 g glacial acetic acid). Genitalia were dissected from adult specimens softened in plastic containers with wet tissue paper for 12–24 hours. The aedeagus was softened in 10% sodium hydroxide solution for 6–12 hours and placed in distilled water. The careful insertion of a sharp-pointed thick nose hair and injection of 5% ethanol into the foramen of aedeagus were repeated until the internal sac was fully everted. After washing with absolute ethanol, the genitalia were preserved in a microvial with glycerin and pinned with the specimen. Descriptions and illustrations were prepared using Nikon SMZ800 and Nikon Eclipse E600 microscopes, each equipped with a drawing tube. Habitus images were taken by a Nikon Coolpix 4500 digital camera attached to a Nikon SMZ1500 microscope. The letters L, S and M in parentheses signify long, short, and micro setae, respectively.
http://zoobank.org/4FEAAE6E-87B0-4AFA-B40F-3DAADB68CF75
http://species-id.net/wiki/Ambrostoma_koreana
Figs 1–20South Korea: Gyeongnam Province, Namhaedo Island, Mangunsan Mountain, 34°51'52"N, 127°51'47"E.
Holotype: male (DBET), KOREA: Gyeongnam Province, Namhaedo Island, Mangunsan Mountain, 34°51'52"N, 127°51'47"E 280 m, 7.IV.2006, H.-W. Cho. Paratypes: 2 males and 6 females (DBET), same data as holotype; 10 males and 10 females (HWC), same data as holotype except for 13.VI.2007, H.-Y. Kwon; 1 male (HWC), KOREA: Gyeongnam Province, Miryang, Gajisan Mountain, 35°35'46"N, 128°59'38"E 500 m, 17.VIII.2001, T.-H. Ahn; 1 female (HWC), same data as preceding paratype except for 25.V.2001, G.-S. Jung.
42 larvae collected or obtained from adults, same data as holotype except for 23.IV.–15.V.2006.
(Figs 1–7, 19).

Habitus of Ambrostoma koreana sp. n., holotype. 1 dorsal view 2 ventral view.
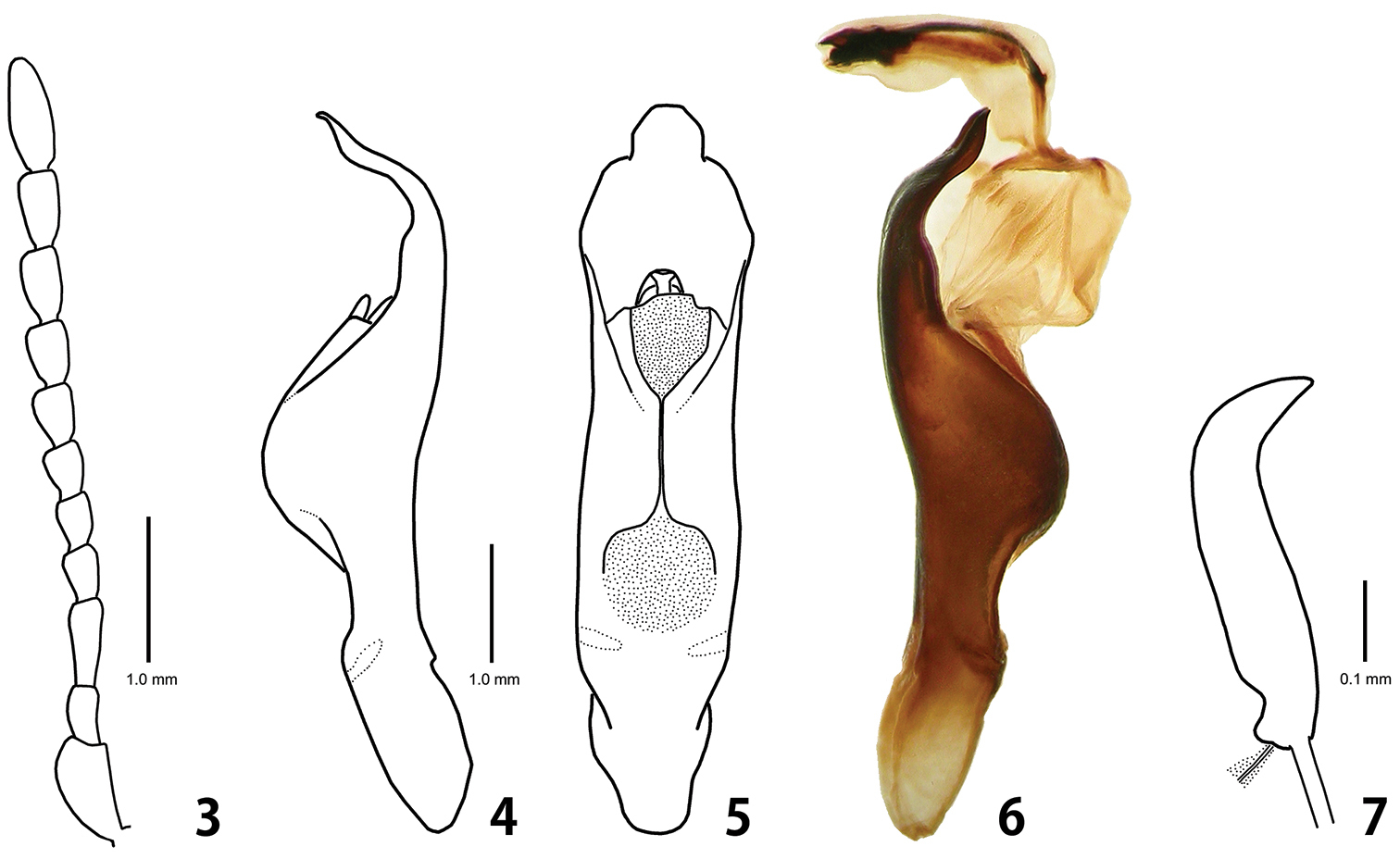
Adult of Ambrostoma koreana sp. n. 3 antenna 4 aedeagus lateral view 5 aedeagus dorsal view 6 aedeagus with everted internal sac laternal view7 spermatheca.
Diagnosis. The new species is very similar in coloration to Ambrostoma leigongshana Wang, but can be distinguished by the following characters: antennomere 3 much longer than 4 (equal in length in Ambrostoma leigongshana); pronotum with moderately dense punctures in lateral depression (dense and coarse punctures in Ambrostoma leigongshana); aedeagus widest at apical 1/5, thence narrowed with trapezoidal apex (elongate apically with rounded apex in Ambrostoma leigongshana); spermatheca almost straight, curved at apex (strongly curved in Ambrostoma leigongshana).
Description. Holotype (Figs 1–2). Body length 11.7 mm, width 5.7 mm, strongly convex dorsally. Head emerald green to ultramarine with 2 pairs of small orange yellow markings on central part. Mouthparts midnight blue. Antennomeres 1–7 ultramarine, 8–11 black. Pronotum emerald green to ultramarine with a pair of large orange yellow markings. Scutellum ultramarine. Elytra ultramarine with a pair of orange yellow markings at base and 2 pairs of orange yellow longitudinal markings on central-posterior part, all markings surrounded by emerald green. Venter mainly ultramarine with greenish luster. Legs midnight blue.
Head. Width 3.4 mm, interocular distance 2.5 mm. Vertex and frons with sparse and small punctures. Clypeus and labrum with small punctures bearing long setae. Mandibles with moderately dense punctures bearing setae on outer surface. Maxillary palp 4-segmented with apical palpomere distinctly widened and truncate. Antennae (Fig. 3) reaching elytral humeri; antennomere 1 robust, longer than 3; antennomere 2 as long as 4; antennomere 3 distinctly longer than 4; antennomeres 5–10 moderately widened; antennomere 11 longest and 2.6 times as long as wide.
Pronotum. Length 2.7 mm, width 5.4 mm. Lateral sides roundly widened anteriorly, widest at anterior 1/4. Anterior margin widely emarginated. Trichobothria on anterior and posterior angles. Disc with dense and small punctures, larger than those of head; hardly confluent large punctures in lateral longitudinal depression; interspaces with spare and minute punctures. Scutellum subtriangular, slightly wider than long, impunctate.
Elytra. Length 8.8 mm, width 6.8 mm. Sides moderately widened posteriorly and widest at posterior 2/5, thence rounded at apex. Disc with double irregular rows of punctures; transverse depression with large punctures, subequal to those of side of pronotum; interspaces with spare and minute punctures. Epipleuron flat, inner margin with micro setae along an entire length. Hind wing well developed.
Venter. Hypomera impunctate; prosternum with sparse and small punctures; prosternal process strongly enlarged and slightly emarginated apically. Abdominal sternites wrinkled laterally with sparse and small punctures; last abdominal sternite deeply emarginated on both sides. Legs moderately robust; tibiae simple without preapical tooth; fore and middle legs with tarsomere 1 slightly narrower than 3; tarsal claws simple.
Aedeagus. Strongly convex at middle, curved and sinuated at apex in lateral view (Fig. 4); subparallel-sided, widest at apical 1/5, thence narrowed with trapezoidal apex, with 2 weakly sclerotized plates in dorsal view (Fig. 5); internal sac curved, shorter than median lobe with thick and long flagellum (Fig. 6).
Paratypes. Body length 10.9–12.7 mm, width 5.2–6.2 mm. Coloration similar to holotype. Female: larger than male; tarsomere 1 of fore and middle legs distinctly narrower than 3; last abdominal sternite rounded; spermatheca (Fig. 7) almost straight, curved at apex.
(Figs 15, 17).
Description. Body length 2.7–4.2 mm, width 1.2–1.7 mm, head width 0.90–0.95 mm (n = 5). Body convex dorsally on abdomen. Pale yellow with head dark brown, tubercles and legs brown (Fig. 17). Tubercles weakly developed (Fig. 15). Sclerotized platelets on dorsum dense and strong, on venter almost absent. Setae longer than in other instars, bases of setae sclerotized.
Head. Vertex and temporal side with 30–32 pairs of long setae and 15–19 pairs of short setae. Frons with 16–19 pairs of long setae. Clypeus and labrum, each with 2 pairs of long setae. Mouthparts similar in shape and chaetotaxy to those of the last instar larva, except for stipes with 3 setae and postmentum with 3 pairs of setae.
Thorax. Prothorax with D-DL-EP (53–57L 4–6S); P (1L); ES (1L) weakly sclerotized; SS represented by a short seta; sternal region with 1–2 additional setae. Meso- and metathorax with Dae (1L); Dpi (1L); dorsal region with 11–13 additional short setae; DLi (1–3L) well developed with egg burster; DLe (4–5L 8–11S); EPa (4–5L 5S) fused with spiracle; EPp (2L 3–4S); P (1L); ES (1L) weakly sclerotized; SS represented by a short seta; sternal region with 2–3 additional setae.
Abdomen. Dorsal and dorsolateral regions with 31–35 short setae; Dpi (1L); Dpe (1L); DLp (1L); EP (9–10L 8–10S); P (3S) divided; PS-SS represented by 4–5 short setae; ES (1S); segment 8 with D-DL (3–4L 11–12S) fused; segment 9 with D-DL-EP (3L 11–12S) fused; segment 10 with pygopod developed; egg burster on segment 1 much smaller than thoracic ones.
Similar to the last instar larva except for following characters: body length 4.2–5.7 mm, width 1.8–2.4 mm, head width 1.25–1.35 mm (n = 4); black markings on head and dark patterns on dorsum much larger than those of the last instar larva; pronotum with black markings partly connected.
Description. Similar to the last instar larva except for following characters: body length 7.2–8.1 mm, width 2.8–3.8 mm, head width 1.90–2.05 mm (n = 3); dark patterns on dorsum much larger than those of the last instar larva.
(Figs 8–14, 16, 18).
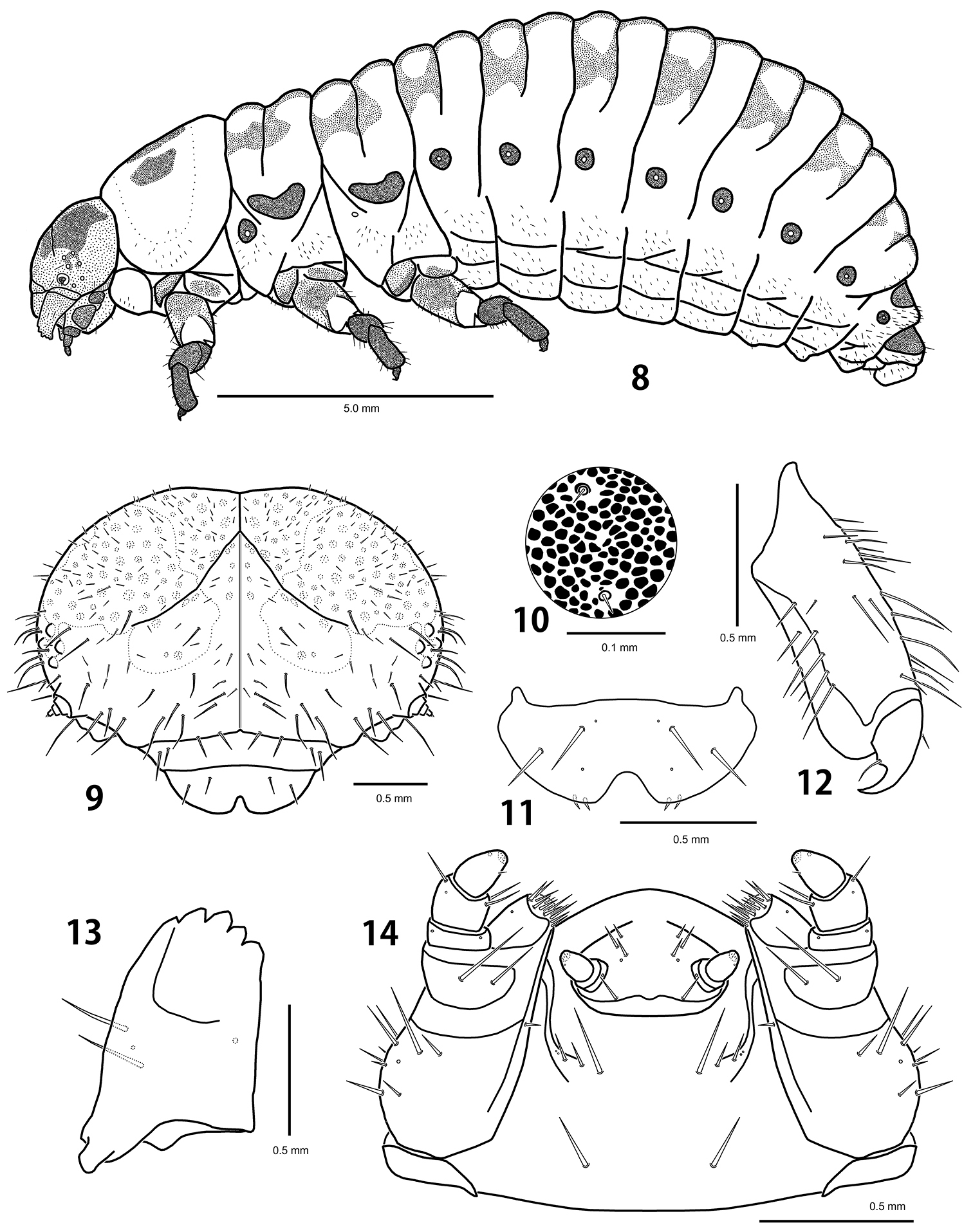
Last instar larva of Ambrostoma koreana sp. n. 8 habitus 9 head 10 sclerotized platelets on dorsal mesothorax 11 labrum 12 tibia and tarsungulus 13 mandible 14 lower mouthparts.
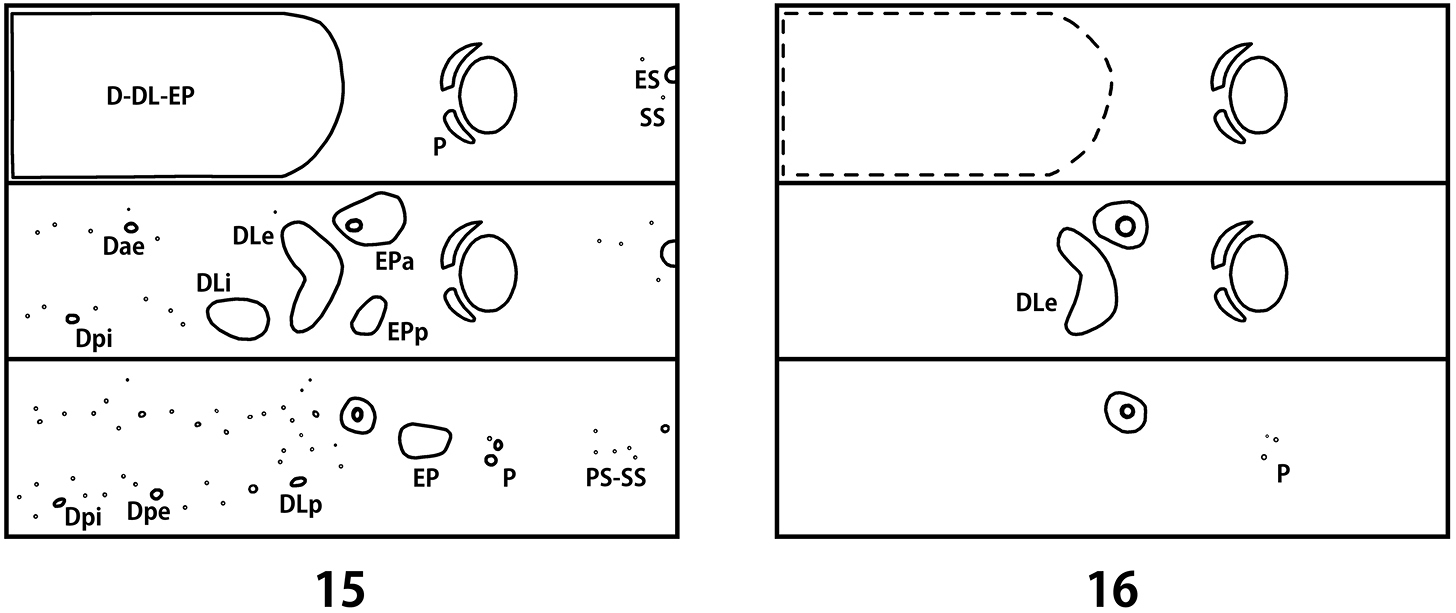
Schematic presentation of tubercular patterns (top: prothorax, middle: mesothorax, bottom: 2nd abdominal segment). 15 first instar larva 16 last instar larva.

Larvae of Ambrostoma koreana sp. n. on leaves of Zelkova serrata. 17 first instar larva 18 last instar larva.

Overwintered adults of Ambrostoma koreana sp. n.
The last instar larva is easily distinguished from larva of Ambrostoma superbum (Thunberg) in the following characters: pronotum with 3 black markings (2 black markings in Ambrostoma superbum); dorsolateral posterior region of each abdominal segment without dark patterns (with dark patterns in Ambrostoma superbum); width of peritreme less than half width of abdominal segment (equal to half width in Ambrostoma superbum).
Body length 11.6–12.8 mm, width 4.7–5.3 mm, head width 2.60–2.65 mm (n = 5). Body strongly convex dorsally on abdomen (Fig. 8). Yellowish white in alcohol specimens, creamy white with orange stripes in live specimens (Fig. 18). Head yellowish brown with a pair of large black markings. Pronotum with 3 black markings. Tubercles, spiracles and legs dark brown. Dorsum with dark patterns consisting of dense and strong sclerotized platelets, sparsely covered with micro setae (Fig. 10). Tubercles undeveloped (Fig. 16). Venter covered with short and moderately long setae, bases of setae not sclerotized.
Head. Hypognathous, rounded, well sclerotized, covered with dark spots (Fig. 9). Vertex and temporal side with 10–11 pairs of long setae and 77–82 pairs of short setae. Epicranial suture Y-shaped; coronal suture distinct along an entire length; frontal suture indistinct for lateral 2/5. Frons slightly depressed medially with 14–15 pairs of long setae and 17–22 pairs of short setae. Endocarina distinct; epistomal suture developed. Six stemmata on each side. Antenna 3-segmented; antennomere 1 without seta; antennomere 2 with a sensory papilla and 4 setae; antennomere 3 with 5 setae. Clypeus trapezoid with 2 pairs of setae. Labrum (Fig. 11) deeply emarginate anteriorly with 2 pairs of setae; epipharynx with 2 pairs of setae. Mandibles (Fig. 13) symmetrical, 5-toothed with 2 setae. Maxillary palp (Fig. 14) 3-segmented; palpomere 1 without seta; palpomere 2 with 3 setae; palpomere 3 conical with a seta; palpiger with 2 setae; mala with 13–14 setae; stipes with 10–13 setae; cardo without seta. Labial palp 2-segmented; prementum with 4–5 pairs of setae; postmentum with 4 pairs of setae.
Thorax. Prothorax with D-DL-EP scattered with micro setae, not sclerotized; P (6–7S); ES and SS, each represented by a short seta; sternal region with 14–16 additional setae. Meso- and metathorax with DLe (27–30M); P (8–10S); EPa and EPp, each represented by 23–27 short setae; ES and SS, each represented by a long seta; sternal region with 20–35 additional setae; mesothoracic spiracle annuliform with large peritreme; metathoracic spiracle vestigial. Legs rather stout; tibia with 28–30 setae; tarsungulus strongly curved; basal tooth weakly developed with a seta (Fig. 12).
Abdomen. Tubercles absent on segments 1–7 except for P represented by 3–4 short setae arising from sclerotized bases; epipleural to sternal regions with a lot of short setae. Segment 8 with D-DL (10–12S); segment 9 with D-DL-EP (12–14L 1–2M); segment 10 with pygopod well developed; spiracle with large peritreme similar to mesothoracic one, but smaller; eversible glands absent.

Larvae inside the abdomen of female of Ambrostoma koreana sp. n.
This endemic species is named after the type locality, Korea.
Specimens were collected in the southeastern part of the Korean Peninsula and on adjacent Namhaedo Island.
Overwintered adults were observed under bark or in fallen leaves near the base of trees in early April (Fig. 19). Adults and larvae fed on leaves of Zelkova serrata Makino (Ulmaceae) which is quite a common tree in Korea. Ulmus pumila Linnaeus (also Ulmaceae) was the only known host plant for three other Ambrostoma species (
| 1 | Punctures of transverse impression of elytra subequal to those of the lateral sides of pronotum | 2 |
| – | Punctures of transverse impression of elytra much finer than those of sides of pronotum | 3 |
| 2 | Elytra with 3 broad transverse bands of purplish copper-red | Ambrostoma rugosopunctatum Chen |
| – | Elytra with 1 transverse and 4 longitudinal bands of orange yellow | Ambrostoma koreana sp. n. |
| 3 | Elytra with large well-defined post-median violaceous patch surrounded by green | 4 |
| – | Elytra without well-defined post-median violaceous patch surrounded by green | 5 |
| 4 | Arrangement of elytral punctures almost entirely irregular; last abdominal sternite of male deeply emarginate on both sides | Ambrostoma fulgurans (Achard) |
| – | Elytral punctures arranged in simple rows at base before transverse impression, slightly irregular beyond impression; last abdominal sternite moderately sinuate on both sides | Ambrostoma fasciatum Chen |
| 5 | Pronotum slightly dilated in front of middle region; interspaces between elytral striae finely and densely punctuate | Ambrostoma superbum (Thunberg) |
| – | Pronotum strongly dilated in front of middle region; interspaces between elytral striae finely and very sparsely punctuate | 6 |
| 6 | Elytra with single striae | Ambrostoma chinkinyui Kimoto & Osawa |
| – | Elytra with double striae | 7 |
| 7 | Pronotum with dense and coarse punctures in lateral depression | 8 |
| – | Pronotum with sparse punctures in lateral depression | Ambrostoma omeishanum Gressitt & Kimoto |
| 8 | Elytra with longitudinal purple markings on media posterior surrounded by green; last abdominal segment in male shallowly sinuate | Ambrostoma leigongshanum Wang |
| – | Elytra with 4 purple longitudinal stripes on media posterior part; last abdominal segment in male deeply sinuate | Ambrostoma fortunei (Baly) |
The Ambrostoma species has not been recorded in Korea since
We are very grateful to Mauro Daccordi (Museo Civico di Storia Naturale, Italy) for many helpful comments. We also thank Hua Yi (Chinese Academy of Sciences, China) for providing the picture of larva of Ambrostoma superbum. This work was supported by scientific grant of the Zoological Institute, University of Wrocław, 2021/M/KBTE/12.