





(C) 2012 Junfeng Zhang. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
A pair of fly impressions is described as a new species of a new genus, Orientisargus illecebrosus gen. et sp. n., referred to a new family Orientisargidae fam. n. within Archisargoidea of Brachycera, Diptera. The systematic position of Orientisargidae is discussed. Daohugosargus gen. n. is proposed for Sharasargus eximius KY Zhang et al., 2008. Uranorhagionidae is a junior synonym for Archisargidae. Meanwhile, Mostovskisarginae is a junior synonym for Uranorhagionidae. Mostovskisargus JF Zhang, 2010 and Strenorhagio KY Zhang et al., 2010 are synonymized with Uranorhagio KY Zhang et al., 2010. Uranorhagio includes three species: Uranorhagio asymmetricus (KY Zhang et al., 2010), comb. n., Uranorhagio daohugouensis KY Zhang et al., 2010 and Uranorhagio deviatus (KY Zhang et al., 2010), comb. n. Strenorhagio grimaldi KY Zhang et al., 2010 is synonymous with Uranorhagio deviatus. Mostovskisargus portentosus JF Zhang, 2010, Mostovskisargus signatus JF Zhang, 2010 and Strenorhagio conjugovenius KY Zhang et al., 2010 are synonymous with Uranorhagio asymmetricus. Brevisolva KY Zhang et al., 2010 is a junior synonym for Mesosolva Hong, 1983. A new specific name, Mesosolva zhangae nom. n., is proposed for Brevisolva daohugouensis KY Zhang et al., 2010. Mesosolva jurassica KY Zhang et al., 2010 should be synonymized under Mostovskisargus sinensis KY Zhang et al., 2010. Sinallomyia nom. n. is proposed for Allomyia Ren, 1998. The systematic positions for Helempis eucalla Ren, 1998, Helempis yixianensis Ren, 1998, Pauromyia oresbia Ren, 1998 and Sinallomyia ruderalis (Ren, 1998) are reassessed. These taxa belong to Archisargidae rather than to Tabanidae, Rhagionidae and Protempididae, respectively.
Diptera, Orientisargidae fam. n., Orientisargus illecebrosus gen. et sp. n. , Archisargidae, Daohugosargus gen. n. , Uranorhaginidae, Jurassic and Early Cretaceous, China
The superfamily Archisargoidea comprises three families: Archisargidae, Kovalevisargidae and Eremochaetidae. The Archocyrtidae is probably the fourth family of archisargoids (
Lately, thousands of brachycerans have been discovered from the “Daohugou Formation” in the vicinity of Daohugou, Ningcheng, Chifeng, Inner Mongolia (
A new family, Orientisargidae fam. n., composed of a new genus and species, Orientisargus illecebrosus gen et sp. n., is described here. Another new genus, Daohugosargus gen. n., is proposed for the known species Sharasargus eximius KY Zhang et al., 2008. Daohugosargus eximius (KY Zhang et al., 2008), comb. n. has a characteristic wing venation, which differs sharply from all the known representatives of archisargids, and may be temporarily assigned to the subfamily Uranorhagioninae (stat. n.) within Archisargidae. The systematic position for Uranorhagionidae is reassessed. It is a junior synonym for Archisargidae, and may be degraded as a subfamily within Archisargidae. Mostovskisarginae is a junior synonym for Uranorhagionidae. Meanwhile, Mostovskisargus JF Zhang, 2010 and Strenorhagio KY Zhang et al., 2010 can be synonymized with Uranorhagio KY Zhang et al., 2010. Two species, Strenorhagio deviatus KY Zhang et al., 2010 and Strenorhagio grimaldi KY Zhang et al., 2010, can be united into one species: Uranorhagio deviatus (KY Zhang et al., 2010), comb. n. Mostovskisargus portentosus JF Zhang, 2010, Mostovskisargus signatus JF Zhang, 2010 and Strenorhagio conjugovenius KY Zhang et al., 2010 are synonyms for Uranorhagio asymmetricus (KY Zhang et al., 2010), comb. n. The species of Mesosolva and related taxa recently described from the “Daohugou Formation” are reassessed: Brevisolva KY Zhang et al., 2010 is a junior synonym for Mesosolva Hong, 1983. A new specific name, Mesosolva zhangae nom. n., is proposed for the Brevisolva daohugouensis KY Zhang et al., 2010. Mesosolva jurassica KY Zhang et al., 2010 should be synonymized under Mesosolva sinensis KY Zhang et al., 2010. A new generic name, Sinallomyia nom. n., is proposed instead of Allomyia Ren, 1998 which is a junior homonym for Allomyia Banks, 1916 (a genus of Trichoptera). The Early Cretaceous Helempis eucalla Ren, 1998, Helempis yixianensis Ren, 1998, Pauromyia oresbia Ren, 1998 and Sinallomyia ruderalis (Ren, 1998) from the Yixian Formation previously placed, respectively, in the Tabanidae, Rhagionidae and Protempididae should be transferred to the Archisarginae of Archisargidae.
Material and methodsSpecimen descriptions, photographs, and drawings were obtained without the application of glycerol to the surface of the specimens. The specimens collected by the author in field were examined under a stereomicroscope (Wild Heerbrugg) and illustrated with the aid of a drawing tube attached to it, re-adjusted using image-editing software (Adobe Photoshop CS). The digital photographs were taken using stereomicroscope (AXioCamHR3).
Wing venation terminology here follows
Orientisargus gen. n.
The type genus only.
Large sized (more than 12 mm) flies. Legs and abdomen slender, strongly pubescent but devoid of bristles; antennal pedicel longest, first flagellomere neither segmented nor arista or style on its tip; wing narrow and long, subpetiolate, alula absent; venation not costalized; R pectinate; C running around entire wing margin although thinned beyond wing tip; C, Sc, R and CuA strong; Sc, R1 and R2+3 long; R4+5 simple (not bifurcated); origin of Rs proximal, nearly at level of M fork; discoidal cell slightly shifted distally; crossvein m-cu absent; M3+4 stem strongly flexed, and touching CuA; cell m3 closed; hind legs stout and long, tibial spurs well developed, empodium wanting; female cerci foliaceous.
This new family demonstrates similar body structures and wing venation to the family Archisargidae Rohdendorf, 1962 based on the following characters: large sized flies with hind legs and abdomen stout and long, body strongly pubescent but devoid of bristles; wing narrow and long, subpetiolate, alula absent; venation: C running around wing margin although thinned beyond wing tip; long and strong Sc and R1 (R1 more than four-fifths of wing length, and clearly stouter than M); and the position of r-m, which meets R4+5 and fore margin of d, respectively; as well as the position of disciodal cell, which is more or less shifted distally. However, from all known representatives of Archisargidae it differs by the simple R4+5, which is not bifurcated, the origin of Rs which is clearly proximal, the strongly flexed M3+4 stem, of which bM3+4 section becomes short, crossvein-like, and touches CuA instead of m-cu; Furthermore, considering the characteristic features that R4+5 is simple, R furcated pectinately and the origin of Rs is proximal, Orientisargidae fam. n. is similar to the family Kovalevisargidae Mostovski, 1997, the sister group of Archisargidae within Archisargoidea. It is distinct from all the kovalevisargids in having longer Sc, R1 and R2+3, the closed cell m3, and the absence of m-cu. It is interesting that the long pedicel, the absence of arista or style on the tip of antenna, the absence of empodium may be the unique features of Orientisargidae and are found neither in Archisargidae nor in Kovalevisargidae. It is also interesting that the new family has a pair of foliaceous cerci on the female terminalia, which is only present in Uranorhagioninae (=Mostovskisarginae) within Archisargidae (JF Zhang, 2010a).
As for m-cu is concerned, an alternative explanation is possible that a very short, but thick, rudimentary m-cu connecting CuA and flex point of M3+4 is present (see Figure 1F, G). In such case, however, it becomes too short to measure.
Orientisargus illecebrosus sp. n.
The type species only.
Latin, orient-, oriental, alluding to the origin of the fossils, and sargus, a common ending in archisargid genera (the masculine gender).
First antennal flagellomere conical. R2+3 arched medially, ending in C before wing tip, and far from R1 end. R4+5 ending beyond wing tip. Rs stem and bR4+5 short. Origin of Rs nearly at level of d base. Rs fork shifted distally of M fork. Crossvein r-m meeting R4+5 and M1+2, near to d base. Four medial veins present. Cells d and m3 narrow and long, the latter cell with long petiole. Section bM3+4 shorter than r-m. CuA and CuP subparallel, and cu cell (traditionally anal cell) wide open.
Usually, the generic diagnosis is covered by the familial diagnosis when the family comprises only one genus. Nevertheless, the familial diagnosis can be well defined based on the characteristics of its sister groups Archisargidae and Kovalevisargidae. In such case, a generic diagnosis is temporarily proposed but need to be revised when another new genus (or genera) within the new family has (have) been discovered.
urn:lsid:zoobank.org:act:08E30A96-3B6C-4705-B7BC-26CBF0A791A8
http://species-id.net/wiki/Orientisargus_illecebrosus
Figures 1–3Latin, illecebrosus, enchanting, alluding to the special wing venation.
Holotype: NIGP DHG901a, NIGP DHG901b, part and counterpart, a pair of nearly complete female archisargoid flies, is held in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences.
“Daohugou Formation”, in the vicinity of Daohugou, Ningcheng, Inner Mongolia, China (uppermost Middle Jurassic – lowermost Upper Jurassic).
The Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences.
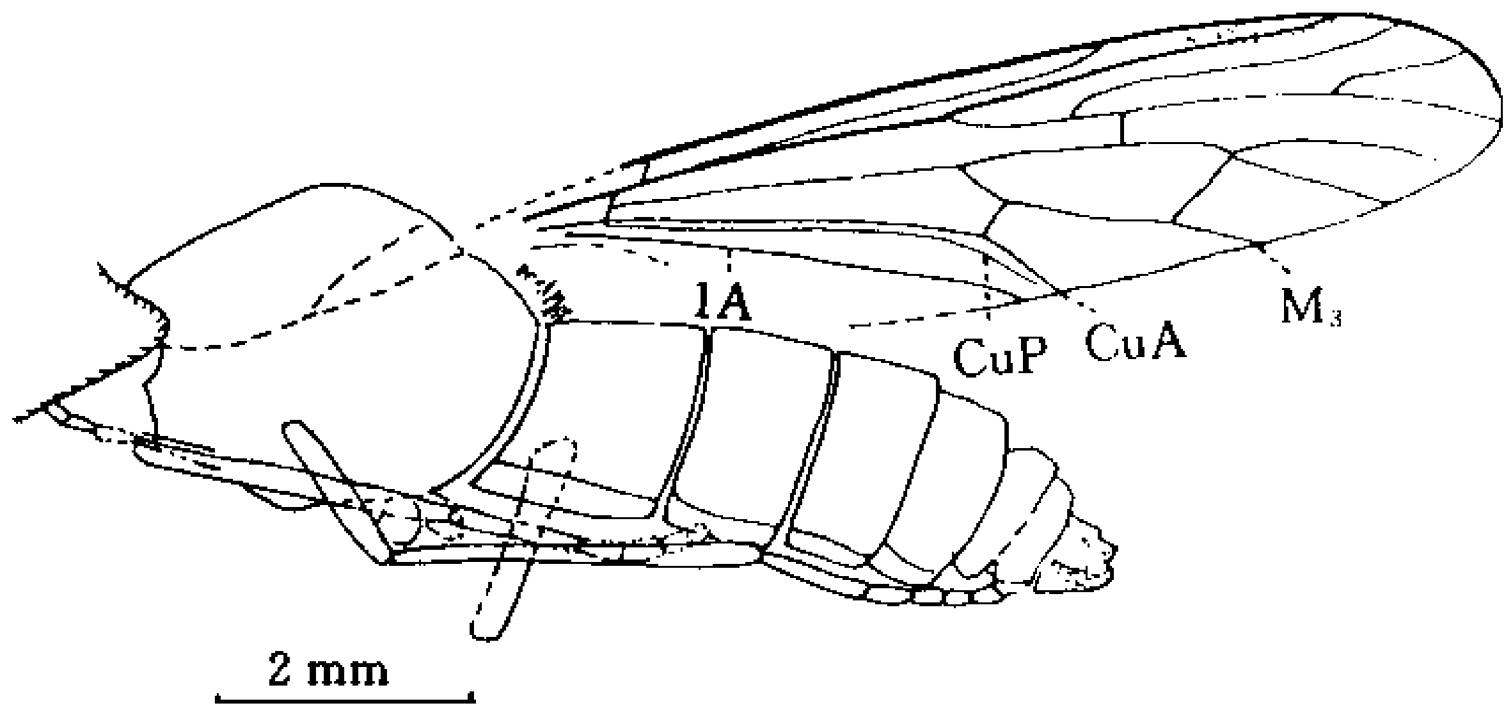
Female insect relatively slender and long. Head and thorax dark brown, otherwise yellowish brown. Head large, semiglobose. Antenna stout, scape ovate, pedicel trapeziform, apically wider than basally, flagellomere short and stout with rounded tip, slightly wider than long. Thorax subovate, clearly longer and wider than head. Wing 3.8 times longer than wide, all veins are markedly thickened except M and CuP, which become moderately thin. Sc nearly four-fifths wing length. R1 straight, more than four-fifths wing length. Origin of Rs slightly basal to midpoint of wing or M fork. Rs stem slightly longer than r-m, and less than one-tenth R2+3 length. Basal section of R2+3 straight, distal section clearly arched forward, ending near to wing tip. Rs fork a little distad to M fork. Section bR4+5 very short, dR4+5 arched, and nearly as long as R2+3. Crossvein r-m dividing anterior margin of discoidal cell as 1:21. Discoidal cell about one-third wing length, and 11 times longer than wide. M1 slightly arched forward, and more than one half diacoidal cell length. M2 straight. Section bM2 some as long as m-m. Section bM3+4 as long as bM1+2, and about one half dM3+4 length. Cell m3 rather narrow and long, some 10 times longer than wide. Section dM3 short, about one half m-m length, and nearly perpendicular to M4. Petiole of cell m3 straight, some one-third cell m3 length. Halter relatively short and stout, club subovate, some one-third halter length. Abdomen cylindrical, more than three times longer than head and thorax combined. Hind leg stout and long. Femur clavate, a little shorter, but stouter, than tibia. Tibial spurs slender and long, nearly one-third basitarsus length. Ratio of tarsomeres 1:0.43:0.28:0.11:0.17, basitarsus nearly as long as remainder tarsomeres combined. Claws small, slender. Female cerci small, longer than ninth tergite.
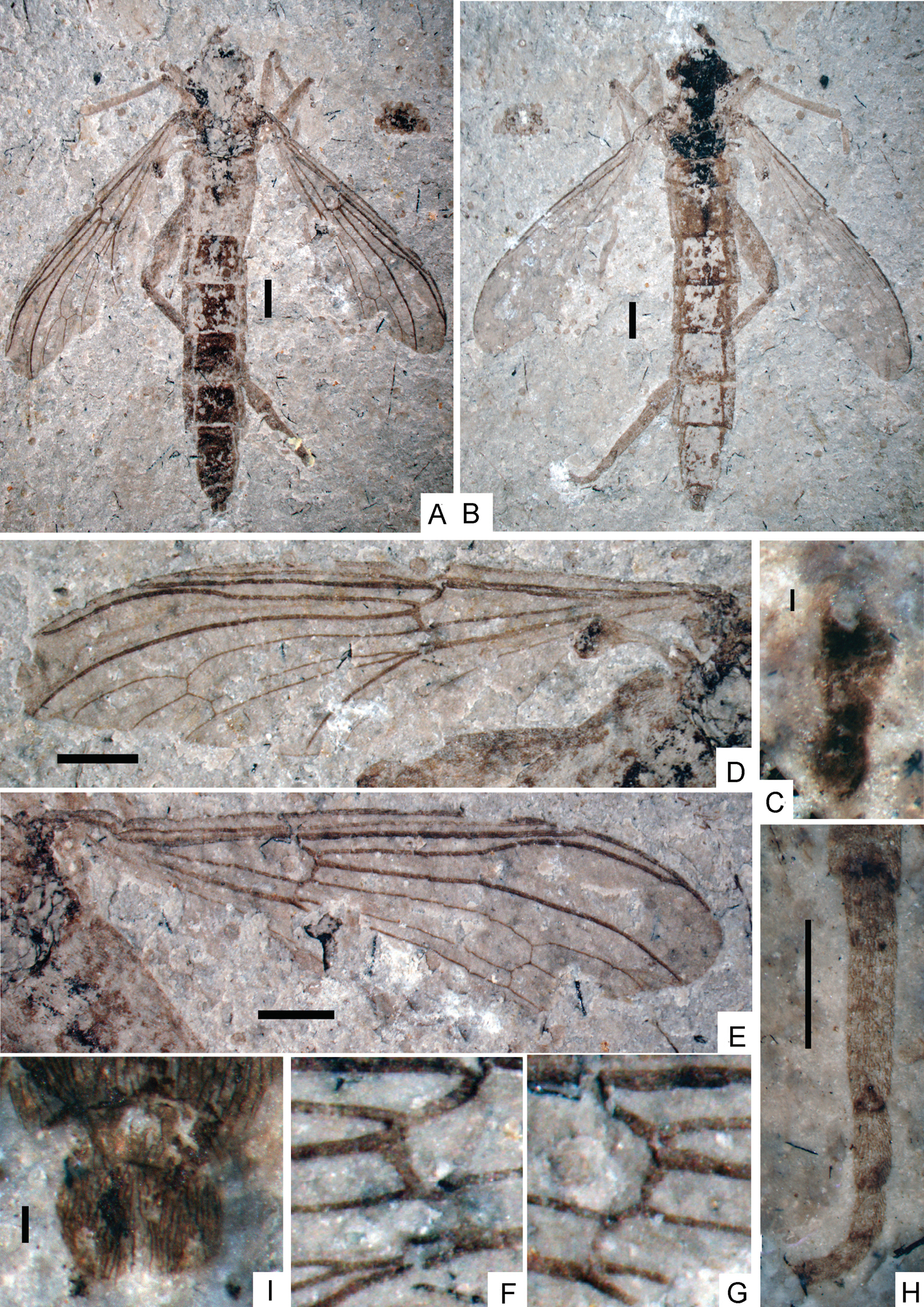
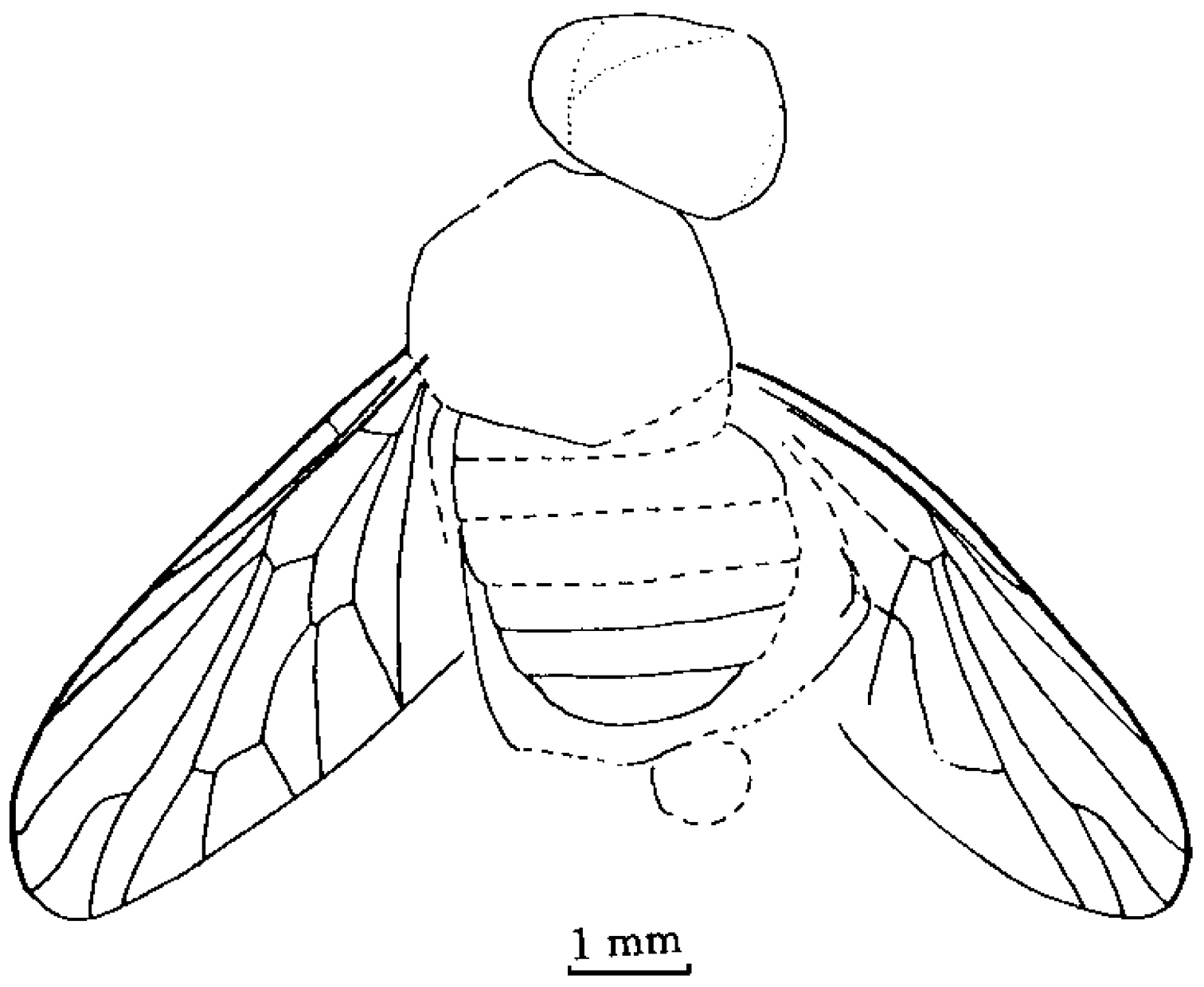
Orientisargus illecebrosus gen et sp. n., NIGP DHG901a, NIGP DHG901b, part and counterpart, holotype, photographs, female, dorsal view. A B body C antenna D left wing E right wing F part enlarged of left wing G part enlarged of right wing H tarsus of hind leg I abdominal apex of female. Scale bars represent 1 mm except for C and I for which scale bars represent 0.1 mm.
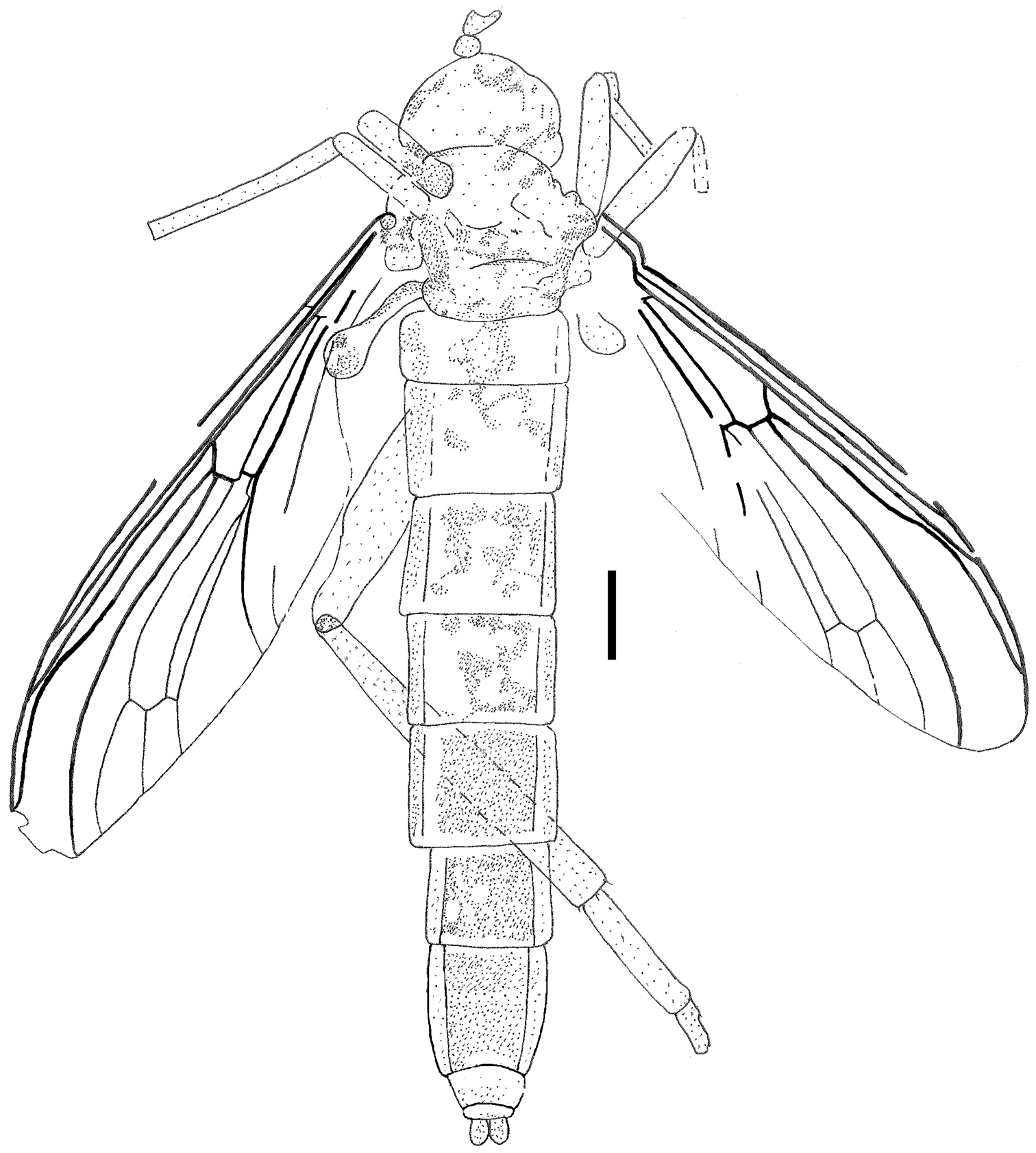
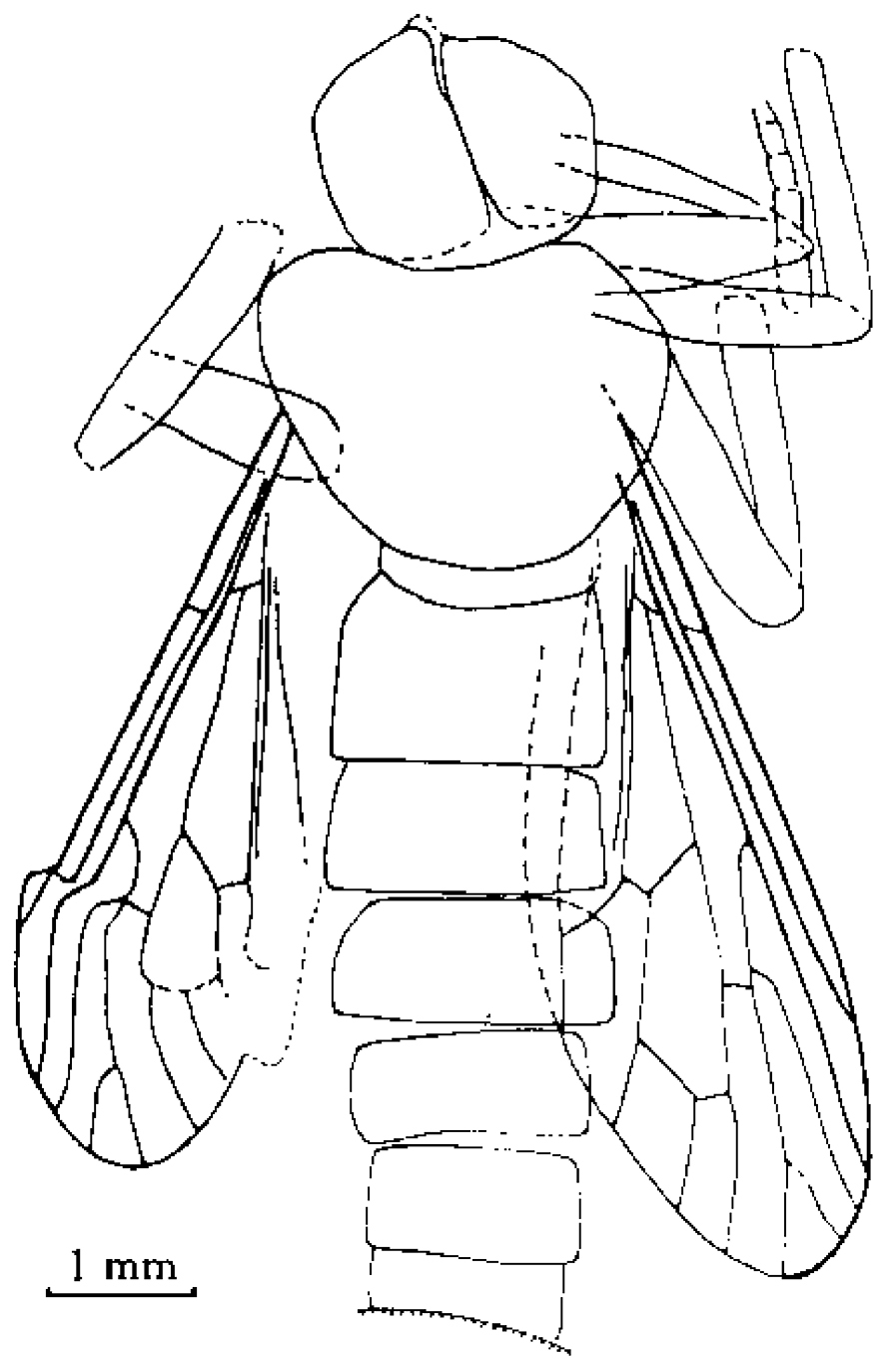
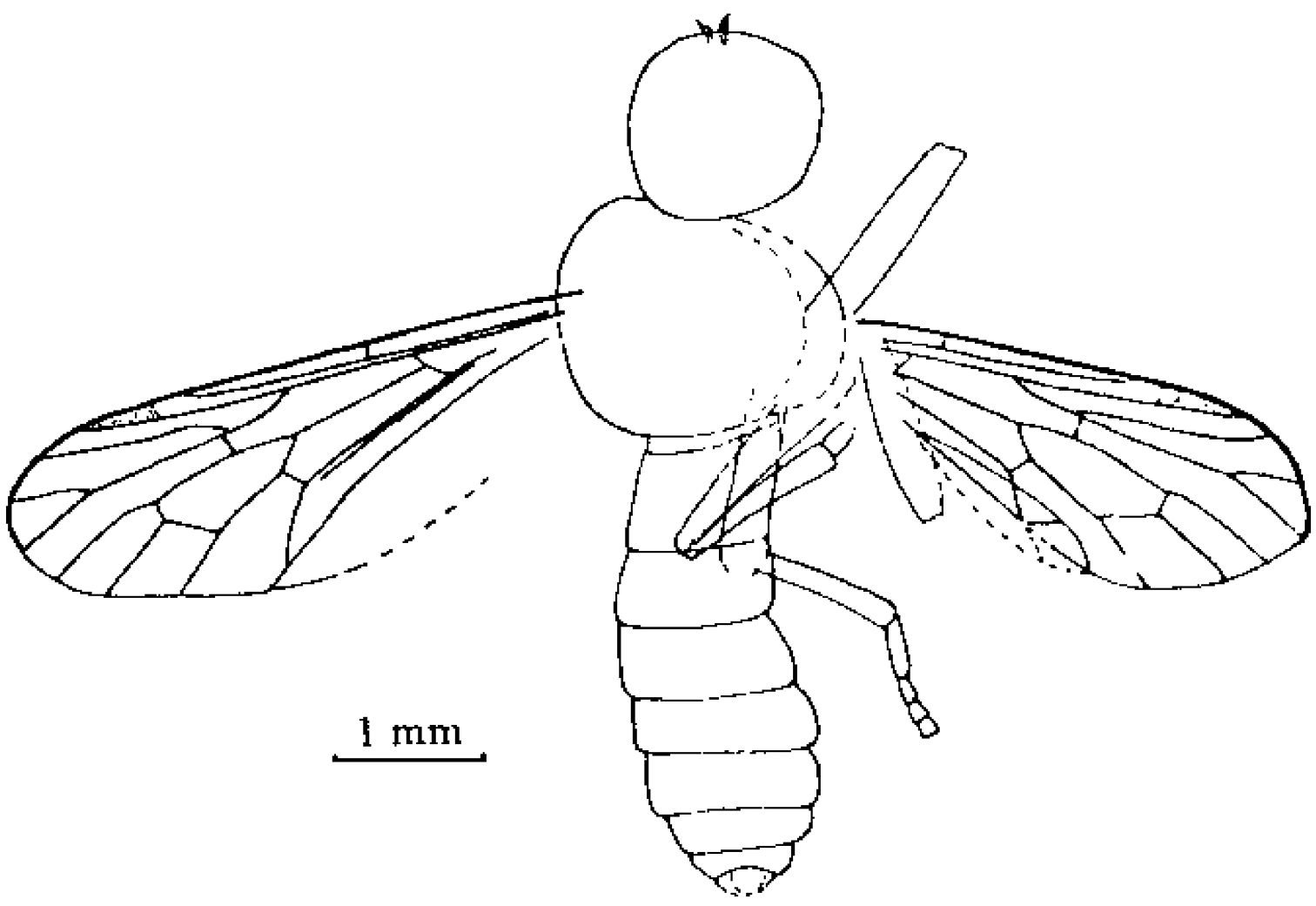
Orientisargus illecebrosus gen et sp. n., body, line drawing of holotype NIGP DHG901a. Scale bars represent 1 mm.
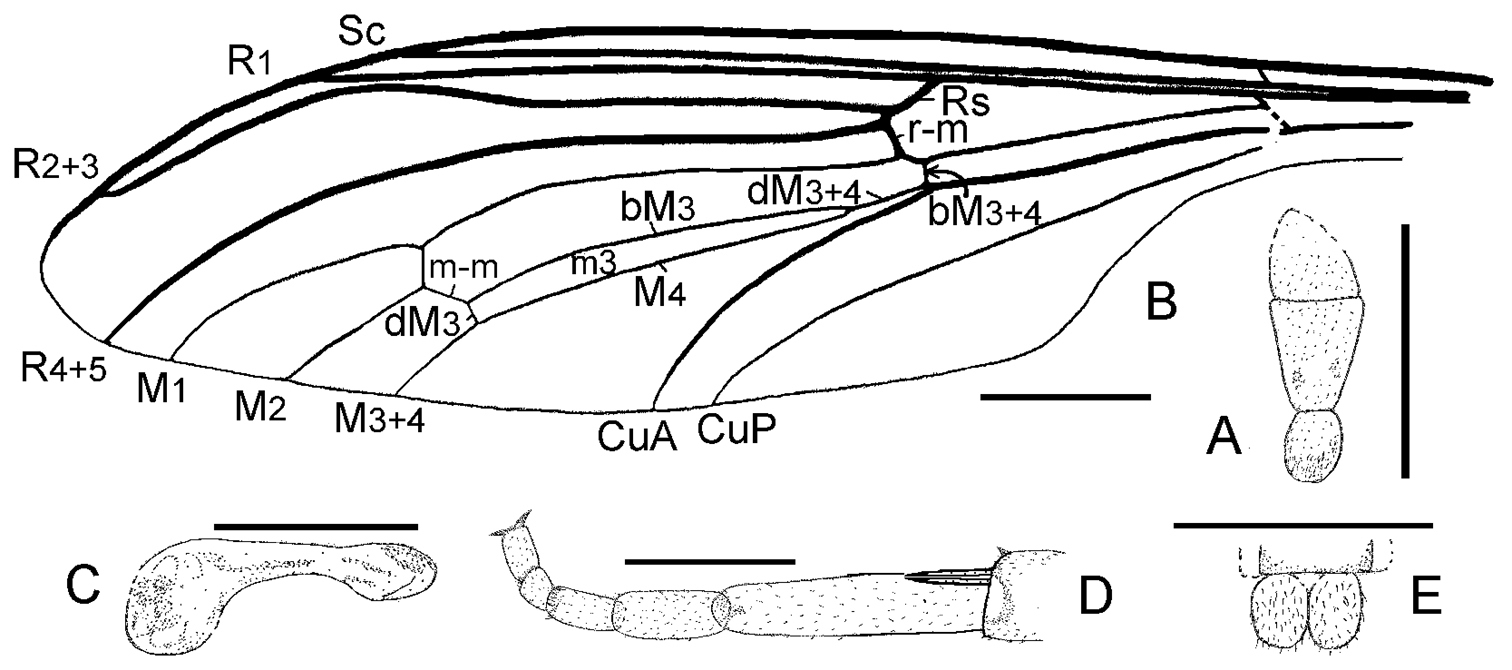
Orientisargus illecebrosus gen et sp. n., line drawings, A antenna of holotype NIGP DHG901b B left wing of holotype NIGP DHG901a (vein sections bM1+2, bM3+4 and dM3+4 reconverted based on right wing) C halter of holotype of NIGP DHG901a D tarsus of hind leg of holotype NIGP DHG901b E female cerci of holotype NIGP DHG901a. Scale bars represent 1 mm.
Length of body 12.2 mm; head, 1.0 mm; thorax, 2.0 mm; abdomen, 9.2 mm. Length of wing, 8.4 mm; width of wing, 2.2 mm. Length of femur of hind leg, c. 4.0 mm; tibia, 4.4 mm; tarsus, 3.2 mm.
http://species-id.net/wiki/Archisargidae
Archisargus Rohdendorf, 1938
Archisarginae Rohdendorf, 1962 and Uranorhagioninae KY Zhang, Yang et Ren, 2010, stat. n. (=Mostovskisarginae JF Zhang, 2010, syn. n.).
Moderate- to large-sized flies. Body robust but usually narrow and long, strongly pubescent but devoid of bristles; first flagellomere of antenna unsegmented, arista well developed at tip of first flagellomere; hind legs stout and long, femora clavate; tibial spurs and pulvilliform empodium well developed; wing narrow and long, subpetiolate, alula absent; all longitudinal veins well developed, ending at wing margin; C running around entire wing margin although thinned near to, or beyond, wing tip, C and R strong, Sc and R1 long, R4+5 bifurcated, R2+3 usually straight and long but in some specific members R2+3 short and significantly curved [see Figure 4, Daohugosargus eximius (KY Zhang et al., 2008) comb. n., originally Sharasargus eximius KY Zhang et al., 2008], in most representatives crossvein r-m meeting R4+5 distad to Rs fork but in some specific members far basad to Rs fork (see Figure 4, Daohugosargus eximius), origin of Rs usually basad to, but in some specific members distad to, d base (see Figure 4, Daohugosargus eximius), discoidal cell shifted distally of wing midpoint.
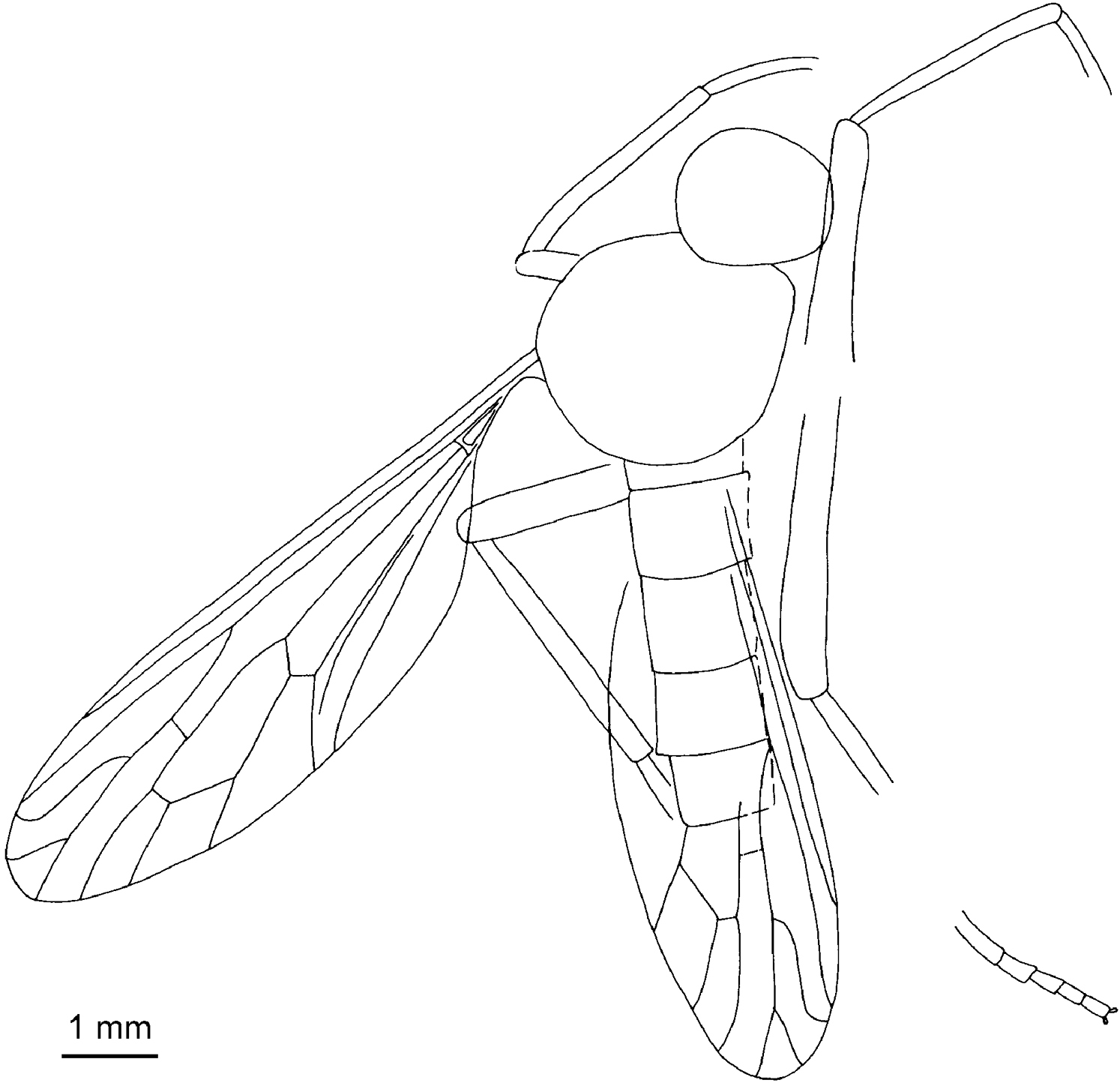
Daohugosargus eximius (KY Zhang, Yang et Ren, 2008) comb. n. (originally Sharasargus eximius KY Zhang, Yang et Ren, 2008; after KY Zhang et al. 2008).
Recently, numerous well preserved archisargid flies have been recovered from the Daohugou biota, China (
Archisargus Rohdendorf, 1938
Archirhagio Rohdendorf, 1938, Archisargus Rohdendorf, 1938, Calosargus Mostovski, 1997, Flagellisargus JF Zhang, 2012, Mesosolva Hong, 1983 (=Prosolva Hong, 1983; Brevisolva KY Zhang et al., 2010, syn. n.), Origoasilus KY Zhang et al., 2011, Ovisargus Mostovski, 1996 (=Helempis Ren, 1998, syn. n.), Parvisargus Mostovski, 1996, Sharasargus Mostovski, 1996 (=Pauromyia Ren, 1998, syn. n.), Sinallomyia nom. n. (pro n Allomyia Ren, 1998).
R1 rather long (four-fifths or more of wing length). R2+3 relatively straight. Crossvein r-m meeting R4+5 distad to Rs fork. Female cerci segmented, not foliaceous.
Uranorhagio KY Zhang, Yang et Ren, 2010
Daohugosargus gen. n. and Uranorhagio KY Zhang, Yang et Ren, 2010 (=Mostovskisargus JF Zhang, 2010; Strenorhagio KY Zhang, Yang et Ren, 2010).
R2+3 significantly bent. Position of r-m inconstant, distad to, just at, or basad to, Rs fork. Female cerci foliaceous, unsegmented.
http://species-id.net/wiki/Uranorhagio
Uranorhagio daohugouensis KY Zhang, Yang et Ren, 2010
Uranorhagio asymmetricus (KY Zhang, Yang et Ren, 2010) comb. n. (=Strenorhagio conjugovenius KY Zhang, Yang et Ren, 2010, syn. n.; Mostovskisargus portentosus JF Zhang, 2010, syn. n.; Mostovskisargus signatus JF Zhang, 2010, syn. n.), Uranorhagio deviatus (KY Zhang, Yang et Ren, 2010) comb. n. (=Strenorhagio grimaldi KY Zhang, Yang et Ren, 2010, syn. n.), besides the type species.
R1 relatively short (some four-fifths of wing length). R2+3 strongly arched basally. Origin of Rs basad to d base. Rs fork basad to midlength of d. Crossvein r-m inconstant: slightly distad to, or just at, or somewhat basad to, Rs fork. M with four terminal branches. M1+2 furcated distad to d end.
Sharasargus eximius KY Zhang, Yang & Ren, 2008
The type species only.
Chinese, Daohugou, alluding to the fossil locality, and sargus, a common ending in archisargid genera (the masculine gender).
R1 long (more than four-fifths of wing length). R2+3 short, significantly S-shaped. Origin of Rs distad to d base. Rs fork nearly at level of d end. Crossvein r-m far basad to Rs fork, and meeting anterior margin of d distad to its midlength. M with three terminal branches. M1+2 bifurcated basad to d end. Cell cu open.
The present new genus differs from Uranorhagio by the short, S-shaped R2+3; the origin of Rs, which is distally of d base, the position of r-m, which is far basad to Rs fork, and meeting anterior margin of d distad to its midlength; and by the M with three terminal branches, on which M1+2 fork basad to d end.
On the other hand, Daohugosargus gen. n. is distinct from Sharasargus Mostovski, 1996 in that: origin of Rs distad to d base; very short, S-shaped R2+3; first fork of Rs nearly at level of d end; and r-m meeting Rs stem far basad to Rs fork.
KY
Comparing Uranorhagionidae with the Archisargidae, however, almost all the characteristics derived from body structures and wing venation in the former family are very closely similar to the latter family. The major difference of significantly bent R2+3 also demonstrates close resemblance to that of Daohugosargus eximius, an undoubted representative of archisargids although its systematic position at generic level is debatable (see Figure 4). It should be noted that the position of r-m in Uranorhagionidae is unstable: in some species slightly distad to, or just at, in other species slightly basad to, Rs fork. Nevertheless, this character also exists in Archisargidae (most representatives versus Daohugosargus eximius). As far as the petiolate M1+2 behind d end is concerned, there are some members of archisargids with M1+2 fork just at d end [see Figure 5, Sharasargus oresbius (Ren, 1998), comb. n., originally Pauromyia oresbia Ren, 1998], which does belong rather to Archisargidae than to Rhagionidae (detailed discussion, see below); and is more or less similar to that of uranorhagionids. The distal position of the M1+2 fork with respect to the d cell distal end may be a unique feature of uranorhagionids, which has the taxonomic significance only at generic, at most subfamilial, rank. Thus, Uranorhagionidae is a junior synonym for Archisargidae, and could be degraded as a subfamily referred to Archisargidae.
KY
Sharasargus oresbius (Ren, 1998) comb. n. (originally Pauromyia oresbia Ren, 1998; after
The author (JF Zhang, 2010a) described two new species referred to a new genus within a new subfamily based on two specimens from the “Daohugou Formation” in the vicinity of Daohugou, Inner Mongolia of China, and assigned them to Archisargidae — Mostovskisarginae: Mostovskisargus: Mostovskisargus portentosus and Mostovskisargus signatus. However, these taxa were published somewhat later (publication date: 16 March, 2010) than those described by KY Zhang et al. (26 February, 2010). Thus, Mostovskisarginae, Mostovskisargus, Mostovskisargus portentosus and Mostovskisargus signatus are junior synonyms of Uranorhagioninae, Uranorhagio and Uranorhagio asymmetricus, respectively.
KY
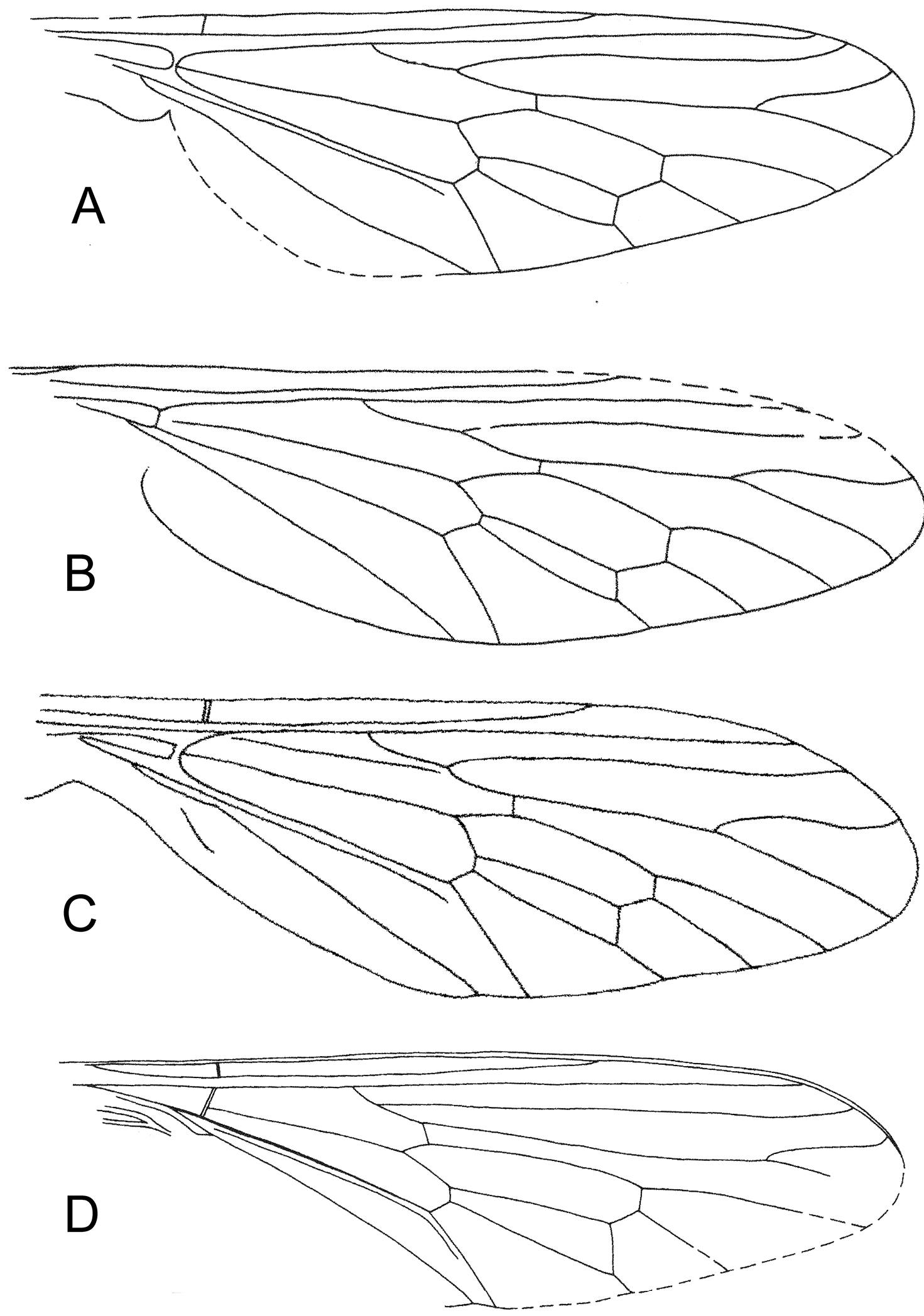
Similarity between four set of wings A Mesosolva sinensis KY Zhang et al., 2010 (after KY Zhang et al., 2010) B Mesosolva sinensis KY Zhang et al., 2010 (originally Mesosolva jurassica KY Zhang et al., 2010; after KY Zhang et al., 2010) C Mesosolva sinensis KY Zhang et al., 2010 (originally Mesosolva jurassica KY Zhang et al., 2010; after KY Zhang et al., 2010) D Mesosolva zhangae (KY Zhang et al., 2010) nom. n. (originally Brevisolva daohugouensis KY Zhang et al., 2010; after KY Zhang et al., 2010).

An undescribed wing of Mesosolva NIGP DHG902.

An undescribed male impression of Mesosolva NIGP DHG903, A habitus (lateral view) B antennae.
Sinallomyia ruderalis from the Lower Cretaceous Yixian Formation was originally regarded as a new genus and species of the subfamily Tabaninae within Tabanidae. Judging from the original illustration (
Sinallomyia ruderalis (Ren, 1998) (originally Allomyia ruderalis Ren, 1998; after
Pauromyia oresbia from the same locality and horizon was previously assigned to the Rhagionidae (
Ovisargus (Helempis) yixianensis (Ren, 1998) comb. n. (originally Helempis yixianensis Ren, 1998; after
Ovisargus (Helempis) yixianensis (Ren, 1998) comb. n. (originally Helempis eucalla Ren, 1998; after
The Origoasilus KY Zhang et al., 2011 previously erected as a new genus and assigned to a new family Origoasilidae KY Zhang et al., 2011 has been transferred to Archisarginae of Archisargidae. The Origoasilidae is a junior synonym for Archisargidae (JF Zhang, 2012b).
Ideally, these previously described species from the upmost Middle–lowest Upper Jurassic “Daohugou Formation” and the Lower Cretaceous Yixian Formation should be revised properly through re-examination of the type material, since the original drawings may contain details resulted from misinterpretation of insufficiently preserved structures. For this reason, until such time as reinvestigation of these specimens is possible, their taxonomic positions could be temporarily assigned to Archisarginae, Archisargidae based on original descriptions and drawings because the Mesozoic archisargid flies have characteristic wing venation (see revised diagnosis of Archisargidae mentioned above) which is easily separated from other extinct and extant families within the lower Orthorrhapha, Brachycera.
I am deeply indebted to Dr M. B. Mostovski (Natal Museum, Pietermaritzburg, South Africa), Dr E. Krzemińska (Department of Invertebrate Zoology, Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Poland) and Dr Dmitry Shcherbakov (Palaeontological Institute, Russian Academy of Sciences, Russia) for critical remarks and improving the previous version of the manuscript. I also sincerely thank Dr Peter Soo (Ex-Brookhaven National Laboratory, Upton, New York, USA) for improving the grammar in the original manuscript. The research was supported by the National Basic Research Program of China (973 Program 2012CB821906) and the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (04ZZ-031309). Open access to this paper was supported by the Encyclopedia of Life (EOL) Open Access Support Project (EOASP).