





(C) 2012 Christophe Faynel. This is an open access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC-BY), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Seven new species of the Neotropical hairstreak genus Oenomaus are described: Oenomaus mancha Busby & Faynel, sp. n. (type locality Ecuador); Oenomaus gwenish Robbins & Faynel, sp. n. (type locality Panama); Oenomaus lea Faynel & Robbins, sp. n. (type locality Ecuador); Oenomaus myrteana Busby, Robbins & Faynel, sp. n. (type locality Ecuador); Oenomaus mentirosa Faynel & Robbins, sp. n. (type locality Peru); Oenomaus andi Busby & Faynel, sp. n. (type locality Ecuador) and Oenomaus moseri Robbins & Faynel, sp. n. (type locality Brazil, Santa Catarina). For each new Oenomaus species, we present diagnostic characters and notes on its habitat and biology. We illustrate adults, genitalia, and distribution. New distributional and biological data are presented for 21 previously described Oenomaus species. Oenomaus melleus guyanensis Faynel, 2008 is treated as a new synonym of Oenomaus melleus melleus (Druce, 1907). Females are described and associated with males for ten species using a variety of factors, including mitochondrial COI DNA “barcode” sequences. We summarize the reasons why the number of recognized Oenomaus species has grown in the past decade from one species to 28 species. Finally, we overview the habitats that Oenomaus species occupy and note that the agricultural pest on Annonaceae, Oenomaus ortygnus, is the only Oenomaus species that regularly occurs in greatly disturbed habitats.
RésuméSept nouvelles espèces appartenant au genre néotropical Oenomaus Hübner sont décrites : Oenomaus mancha Busby & Faynel, sp. n. (localité type : Equateur); Oenomaus gwenish Robbins & Faynel, sp. n. (localité type : Panama); Oenomaus lea Faynel & Robbins, sp. n. (localité type : Equateur); Oenomaus myrteana Busby, Robbins & Faynel, sp. n. (localité type : Equateur); Oenomaus mentirosa Faynel & Robbins, sp. n. (localité type : Pérou); Oenomaus andi Busby et Faynel, sp. n. (localité type : Equateur) et Oenomaus moseri Robbins & Faynel, sp. n. (localité type : Brésil, Santa Catarina). Pour chaque nouvelle espèce, les caractères diagnostiques sont présentés et sont accompagnés de remarques sur l’habitat et la biologie. Les faces dorsales et ventrales des adultes et les organes génitaux sont illustrés et des cartes de répartition sont données. Dans une deuxième partie, de nouvelles données sont présentées pour 21 espèces d’Oenomaus précédemment décrites. Oenomaus melleus guyanensis Faynel, 2008 est mis en synonymie avec Oenomaus melleus melleus (Druce, 1907). Les femelles sont décrites et associées avec des mâles en utilisant plusieurs moyens, y compris les séquences d’ADN mitochondrial COI. Les raisons pour lesquelles le nombre d’espèces d’Oenomaus décrites a fortement augmenté ces dernières années sont évoquées. Finalement, une vue d’ensemble des habitats occupés par les différentes espèces d’Oenomaus est présentée. Oenomaus ortygnus, qui est un parasite agricole connu sur Annonaceae, est la seule espèce d’Oenomaus qui se trouve régulièrement dans des habitats perturbés.
Annonaceae, Neotropics, Porthecla
The widespread Neotropical hairstreak Oenomaus ortygnus (Cramer) is a pest of cultivated soursop (Annona muricata L.) and relatives (Annonaceae), and aspects of its biology have been documented for nearly a century (e.g.,
A close phylogenetic relationship between Oenomaus and Porthecla Robbins was suggested when
We present new species level taxonomic information for Oenomaus in this paper. We describe seven new Oenomaus species. Next, we update information on the distribution, habitat, variation, and biology of the 21 species that were previously described in or transferred to Oenomaus (
Genitalic terms follow those in
Illustrated adults of Oenomaus are noted in the material examined sections, and each genitalia drawing is of the adult illustrated. The structure of the male genitalia valvae in Oenomaus is complex, for which reason we present them in ventral, lateral, and dorsal views.
Biogeographical zones follow
The ventral wing pattern in Oenomaus is sexually monomorphic, so associating the sexes of species with distinct ventral wing patterns, such as Oenomaus ortygnus, is straightforward. However, a majority of Oenomaus species have a ventral wing pattern that is similar to that of Oenomaus atena (Hewitson). Among these, some can be associated because they have distinct ventral wing pattern elements, such as those of Oenomaus isabellae (
The mitochondrial COI gene sequence (commonly called a DNA “barcode”) has been useful, when combined with other characters, in distinguishing lepidopteran species in a single area (e.g.,
We use the following acronyms for collections, following those for public institutions listed on the website hbs.bishopmuseum.org/codens/codens-inst.html (accessed April 2012):
AMNH American Museum of Natural History, New York, New York, USA.
ANSP Academy of Natural Sciences, Philadelphia, Pennsylvania, USA.
CF Private collection of Christophe Faynel, France.
CMNH Carnegie Museum of Natural History, Pittsburgh, Pennsylvania, USA.
DZUP Universidade Federal do Paraná, Curitiba, Paraná, Brazil.
FSMC Florida Museum of Natural History, Allyn Museum, University of Florida, Gainesville, Florida, USA.
JFLC Private collection of Jean François Le Crom, Bogotá, Colombia.
LYD Private collection of Louis and Yvan Diringer, France.
MC Private collection of Alfred Moser, São Leopoldo, Rio Grande do Sul, Brazil.
MNHN Muséum national d’Histoire naturelle, Paris, France.
MUSM Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Lima, Perú.
OSAC Oregon State University Corvallis, Oregon, USA.
PB Private collection of Pierre Boyer, Le Puy Sainte Réparade, France.
RCB Private collection of Robert C. Busby, Andover, Massachusetts, USA.
SMF Forschungsinstitut und Naturmuseum Senckenberg, Frankfurt-am-Main, Germany.
USNM Smithsonian Institution, Washington, DC USA.
New speciesA distinguishing trait of Oenomaus and Porthecla among members of the Panthiades Section of the Eumaeini is the lack of an orange cubital spot (
urn:lsid:zoobank.org:act:4DF6102F-0A25-46B8-8F56-6E33D791A5F2
http://species-id.net/wiki/Oenomaus_mancha
Figs 1, 2, 20, 26, 38, 46Ecuador: Sucumbíos, 5 km Puerto Libre-La Bonita Road, 0°13.0'N, 77°29.3'W, 700 m. The road going west from Puerto Libre increases in elevation as the terrain becomes hillier. The collecting spot was in wet forest and was easily accessed by a muddy logging trail. Since 2005, logging has continued, leaving very few tall trees in the once beautiful forest.
Holotype ♂ (Fig. 1) labeled as “ECUADOR: Sucumbios / 5 km Puerto Libre-La Bonita Road / 0°13.0'N, 77°29.3'W, 700 m / 23 February 2005 / Robert C. Busby, leg.” [rectangular, white, printed], “11:00 hrs / 5 m” [rectangular, white, handwritten, blue ink], “GENITALIA No. / 2011: 419♂ / C. FAYNEL” [rectangular, green, printed] “Holotype ♂ / Oenomaus mancha / Busby & Faynel, 2012” [rectangular, red, printed]. Deposited in USNM.
Paratypes: Ecuador. 2 ♂: Napo, 14 km Tena-Puyo Road, 1°06.7'S, 77°46.9' W, 600 m, 24.IX.2011, (Apuya) Robert C. Busby leg. (RCB); Napo, Pimpilala, [ GPS data : 1°04.6 S, 77°56.2'W ], 600–700 m, Euclides Aldaz leg., XII.2003, gen. prep. CF n°290 (PB); 10♀: Napo, 28 km Tena-Puyo Road, 1°11.3'S, 77°49.9'W, 800 m, VIII.2006 (El Capricho) I. Aldas & R. C. Busby leg. (RCB); Napo, 12 km Tena-Puyo Road, 1°05.3'S, 77°47.4' W, 600 m, 28.VIII.2009, (Finca San Carlo) D. H. Ahrenholz, R. C. Busby, leg. (RCB); Napo, 14 km Tena-Puyo Road, 1°06.7'S, 77°46.9'W 600 m, VIII.2005, (Apuya) I. Aldas & R. C. Busby leg. (RCB) ; Napo, 14 km Tena-Puyo Road, 1°06.7'S, 77°46.9'W, 600 m, 17.X.2010, (Apuya) I. Aldas & R. C. Busby leg. (RCB); Napo, 14 km Tena-Puyo Road, 1°06.7'S, 77°46.9'W, 600 m, 22.X.2010, (Apuya) I. Aldas & R. C. Busby leg. (RCB); Pastaza Province, 32 km S. of Puyo, 1000 m, 21–23.X.1995 Robert C. Busby leg. (RCB); Pastaza Province, 45 km Puyo-Arajuno Rd, 1000 m, 15.IX.1999, Robert C. Busby leg., gen. prep. CF n°420 (RCB); Pastaza Province, 45 km Puyo-Arajuno Rd, 1000 m, 26.IX.1999, Robert C. Busby leg., gen. prep. CF n°421 (RCB); Pastaza Province, 45 km Puyo-Arajuno Rd, 1000 m, 26.IX.1999, Robert C. Busby leg. (RCB); Pastaza, Puyo, 1000 m, 14.X.1989, D.H. Ahrenholz leg., gen. prep. CF n°407 (USNM ENT 00180037) (Fig. 2).

Oenomaus, new species: adults (dorsal surface at left, ventral surface at right). 1 ♂ Oenomaus mancha (holotype, Ecuador) 2 ♀ Oenomaus mancha (paratype, Ecuador) 3 ♀ Oenomaus gwenish (holotype, Panama) 4 ♂ Oenomaus lea (holotype, Peru) 5 ♂ Oenomaus myrteana (holotype, Ecuador) 6 ♀ Oenomaus myrteana (paratype, Ecuador) 7 ♂ Oenomaus mentirosa (holotype, Peru) 8 ♂ Oenomaus andi (holotype, Ecuador) 9 ♀ Oenomaus andi (paratype, Ecuador) 10 ♂ Oenomaus moseri (holotype, Brazil) 11 ♀ Oenomaus moseri (paratype, Brazil).
Male FW length: 20.8 mm (SD = 1.9, N = 2). Female FW length: 19.4 mm (SD = 0.5, N = 3). Wing pattern (Figs 1, 2) and genitalia (Figs 20, 26) illustrated. Oenomaus mancha, Oenomaus ortygnus, and Oenomaus gwenish (named below) share a unique ventral wing pattern in which the VFW postmedian line (displaced basally, but by tradition still called the postmedian line) is composed of “disjointed” large black spots on a gray ground color (Fig. 3 for Oenomaus gwenish and Figs 2, 4 for Oenomaus ortygnus in

Oenomaus, newly associated females: adults (dorsal surface at left, ventral surface at right). 12 Oenomaus geba (Brazil) 13 Oenomaus magnus (French Guiana) 14 Oenomaus brulei (French Guiana) 15 Oenomaus gaia (Panama) 16 Oenomaus cyanovenata (Costa Rica) 17 Oenomaus cyanovenata (French Guiana) 18 Oenomaus taua (Panama) 19 Oenomaus taua (Ecuador).

Oenomaus male genitalia: ventral view at left (setae drawn only on the right side and penis removed), left lateral view in the middle (with the penis and valvae displaced ~45° to make them more visible), and dorsal view at right (with setae drawn only on the right side and penis removed). 20 Oenomaus mancha (holotype, Ecuador) 21 Oenomaus lea (holotype, Peru) 22 Oenomaus myrteana (holotype, Ecuador) 23 Oenomaus mentirosa (holotype, Peru) 24 Oenomaus andi (holotype, Ecuador) 25 Oenomaus moseri (holotype, Brazil).
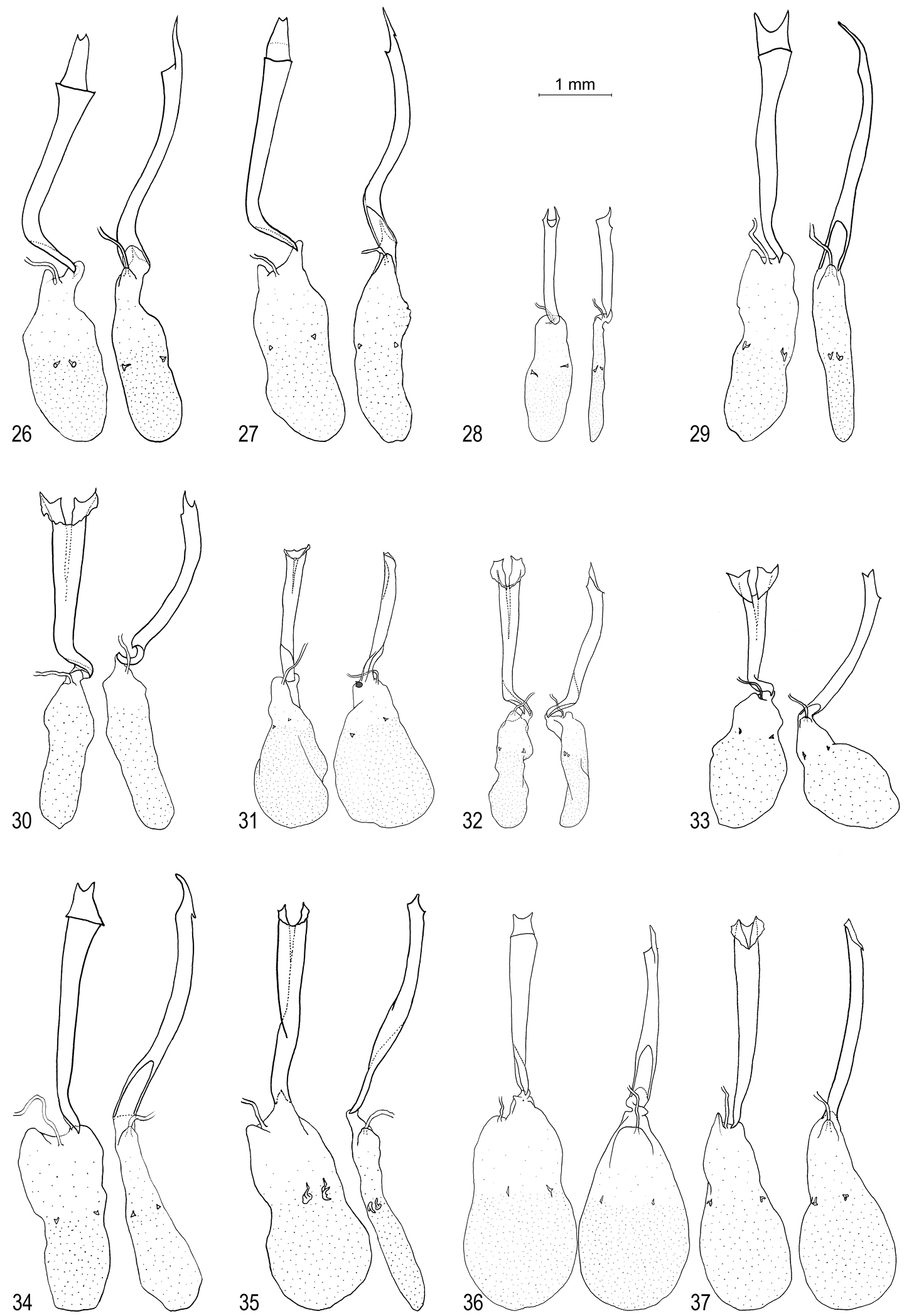
Oenomaus female genitalia: ventral view at left, lateral view on right. 26 Oenomaus mancha (paratype, Ecuador) 27 Oenomaus gwenish (holotype, Panama) 28 Oenomaus myrteana (paratype, Ecuador) 29 Oenomaus andi (paratype, Ecuador) 30 Oenomaus moseri (paratype, Brazil) 31 Oenomaus brulei (French Guiana) 32 Oenomaus cyanovenata (French Guiana) 33 Oenomaus cyanovenata (Costa Rica) 34 Oenomaus gaia (Panama) 35 Oenomaus geba (Brazil) 36 Oenomaus magnus (French Guiana) 37 Oenomaus taua (Panama).
Comparison of inter- and intraspecific divergences (in % rounded to the nearest tenth) for the DNA “barcodes” of 19 Oenomaus male species obtained with BOLD (noted as -- when not available). Number of males examined in brackets.
| Oenomaus ambiguus | Oenomaus atena | Oenomaus atesa | Oenomaus brulei | Oenomaus cortica | Oenomaus curiosa | Oenomaus cyanovenata | Oenomaus gaia | Oenomaus isabellae | Oenomaus jauffreti | Oenomaus lea | Oenomaus magnus | Oenomaus mancha | Oenomaus morroensis | Oenomaus moseri | Oenomaus nigra | Oenomaus ortygnus | Oenomaus poirieri | Oenomaus taua | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oenomaus ambiguus (3) | 0.2 | ||||||||||||||||||
| Oenomaus atena (1) | 6.9 | -- | |||||||||||||||||
| Oenomaus atesa (1) | 7.2 | 7.0 | -- | ||||||||||||||||
| Oenomaus brulei (2) | 6.2 | 6.2 | 7.4 | 0.3 | |||||||||||||||
| Oenomaus cortica (3) | 2.8 | 5.5 | 7.5 | 5.9 | 0.5 | ||||||||||||||
| Oenomaus curiosa (4) | 7.9 | 6.5 | 5.3 | 6.9 | 7.4 | 0.6 | |||||||||||||
| Oenomaus cyanovenata (4) | 7.1 | 5.4 | 7.2 | 7.3 | 5.8 | 6.4 | 0 | ||||||||||||
| Oenomaus gaia (3) | 2.4 | 5.3 | 7.0 | 5.6 | 1.0 | 7.2 | 5.9 | 0.7 | |||||||||||
| Oenomaus isabellae (1) | 6.2 | 6.7 | 7.6 | 7.3 | 5.8 | 7.9 | 6.1 | 5.6 | -- | ||||||||||
| Oenomaus jauffreti (4) | 7.0 | 6.5 | 7.0 | 6.2 | 6.4 | 7.6 | 6.9 | 6.1 | 4.9 | 1.6 | |||||||||
| Oenomaus lea (1) | 6.9 | 4.7 | 6.7 | 6.3 | 6.1 | 6.2 | 5.9 | 5.9 | 7.3 | 5.8 | -- | ||||||||
| Oenomaus magnus (1) | 7.9 | 6.9 | 7.6 | 7.6 | 7.0 | 7.5 | 6.9 | 6.8 | 4.5 | 5.9 | 7.7 | -- | |||||||
| Oenomaus mancha (1) | 5.2 | 5.2 | 5.7 | 5.7 | 4.9 | 5.9 | 5.6 | 4.3 | 4.9 | 5.4 | 5.6 | 6.4 | -- | ||||||
| Oenomaus morroensis (1) | 2.3 | 4.8 | 7.1 | 4.8 | 0.9 | 7.1 | 5.4 | 0.9 | 5.7 | 6.3 | 5.7 | 6.8 | 5 | -- | |||||
| Oenomaus moseri (2) | 5.7 | 5.9 | 6.7 | 6.9 | 5.7 | 6.9 | 4.0 | 5.2 | 5.6 | 5.8 | 5.8 | 6.6 | 4.9 | 5.1 | 0 | ||||
| Oenomaus nigra (1) | 7.7 | 6.7 | 5.7 | 8.4 | 7.8 | 5.9 | 7.6 | 7.2 | 7.8 | 6.9 | 5.9 | 7.2 | 6.4 | 7.7 | 7 | -- | |||
| Oenomaus ortygnus (2) | 6.7 | 5.7 | 6.6 | 7.1 | 6.3 | 6.4 | 6.3 | 6.1 | 6.3 | 6.3 | 6.3 | 6.2 | 3.5 | 6.2 | 5.4 | 6.6 | 1.5 | ||
| Oenomaus poirieri (1) | 7.7 | 6.9 | 7.5 | 7.2 | 7.5 | 8.0 | 6.6 | 7.0 | 5.2 | 4.6 | 7.5 | 6.2 | 6.1 | 6.9 | 5.3 | 7.6 | 6.2 | -- | |
| Oenomaus taua (2) | 5.9 | 5.2 | 5.4 | 6.1 | 5.3 | 6.0 | 5.1 | 4.9 | 5.2 | 4.7 | 4.3 | 5.9 | 4.1 | 5.4 | 4.8 | 6.1 | 4.9 | 5.6 | 0 |
Species of Oenomaus sampled with BOLD (project NLYCA), with sample identifications, localities and GenBank accession numbers.
| Species | Sample ID | Locality | GenBank Accession Numbers |
| Oenomaus ambiguus Faynel, 2008 | CF-LYC-025 | Peru | HQ966548 |
| Oenomaus ambiguus Faynel, 2008 | CF-LYC-183 | Peru | JX458731 |
| Oenomaus ambiguus Faynel, 2008 | CF-LYC-189 | Peru | JX458734 |
| Oenomaus atena (Hewitson, 1867) | CF-LYC-084 | Peru | HQ966592 |
| Oenomaus atesa (Hewitson, 1867) | CF-LYC-003 | French Guiana | HQ966543 |
| Oenomaus brulei Faynel, 2008 | CF-LYC-033 | French Guiana | HQ966552 |
| Oenomaus brulei Faynel, 2008 | CF-LYC-035 | French Guiana | HQ966554 |
| Oenomaus cortica (D'Abrera, 1995) | CF-LYC-051 | Brazil | HQ966565 |
| Oenomaus cortica (D'Abrera, 1995) | CF-LYC-052 | Brazil | HQ966566 |
| Oenomaus cortica (D'Abrera, 1995) | CF-LYC-188 | Peru | JX458722 |
| Oenomaus curiosa Faynel & Moser, 2008 | CF-LYC-036 | French Guiana | HQ966555 |
| Oenomaus curiosa Faynel & Moser, 2008 | CF-LYC-037 | French Guiana | HQ966556 |
| Oenomaus curiosa Faynel & Moser, 2008 | CF-LYC-016 | Peru | JX458726 |
| Oenomaus curiosa Faynel & Moser, 2008 | CF-LYC-184 | Peru | JX458730 |
| Oenomaus cyanovenata (D'Abrera, 1995) | CF-LYC-049 | Brazil | HQ966564 |
| Oenomaus cyanovenata (D'Abrera, 1995) | CF-LYC-048 | Brazil | HQ966563 |
| Oenomaus cyanovenata (D'Abrera, 1995) | CF-LYC-047 | French Guiana | JX458737 |
| Oenomaus cyanovenata (D'Abrera, 1995) | CF-LYC-182 | Peru | JX458728 |
| Oenomaus gaia Faynel, 2008 | CF-LYC-024 | Peru | JX458720 |
| Oenomaus gaia Faynel, 2008 | CF-LYC-023 | French Guiana | JX458719 |
| Oenomaus gaia Faynel, 2008 | CF-LYC-187 | Peru | JX458721 |
| Oenomaus isabellae Faynel, 2006 | CF-LYC-006 | Brazil | HQ966545 |
| Oenomaus jauffreti Faynel & Moser, 2008 | CF-LYC-030 | Brazil | JX458724 |
| Oenomaus jauffreti Faynel & Moser, 2008 | CF-LYC-029 | French Guiana | HQ966549 |
| Oenomaus jauffreti Faynel & Moser, 2008 | CF-LYC-028 | Brazil | JX458727 |
| Oenomaus jauffreti Faynel & Moser, 2008 | CF-LYC-186 | Peru | JX458732 |
| Oenomaus lea Faynel & Robbins, 2012 | CF-LYC-005 | Peru | HQ966544 |
| Oenomaus magnus Faynel & Moser, 2008 | CF-LYC-020 | Peru | HQ966547 |
| Oenomaus mancha Busby & Faynel, 2012 | CF-LYC-190 | Ecuador | JX458723 |
| Oenomaus morroensis Faynel & Moser, 2008 | CF-LYC-015 | Brazil | JX458736 |
| Oenomaus moseri Robbins & Faynel, 2012 | CF-LYC-012 | Brazil | JX458735 |
| Oenomaus moseri Robbins & Faynel, 2012 | CF-LYC-063 | Brazil | HQ966576 |
| Oenomaus nigra Faynel & Moser, 2008 | CF-LYC-148 | Peru | JX458729 |
| Oenomaus ortygnus (Cramer, 1779) | CF-LYC-146 | Mexico | JX458738 |
| Oenomaus ortygnus (Cramer, 1779) | CF-LYC-147 | Peru | JX458733 |
| Oenomaus poirieri Faynel, 2008 | CF-LYC-011 | French Guiana | JX458725 |
| Oenomaus taua Faynel & Moser, 2008 | CF-LYC-085 | Peru | HQ966593 |
| Oenomaus taua Faynel & Moser, 2008 | CF-LYC-185 | Peru | JX458739 |
The name of this species is derived from the Spanish word “mancha”, which means spot, referring to the very distinctive, elongated black spot in VHW cell Sc+R1-Rs. The name is a feminine noun in apposition.
Oenomaus mancha occurs widely in wet forest in eastern Ecuador at elevations ranging from 400 to 1100 m (Fig. 46). Although it is sympatric with Oenomaus ortygnus in wet forest, it does not occur in the highly disturbed habitats in which Oenomaus ortygnus sometimes occurs. It is yet an open question whether Oenomaus mancha is a lowland or lower montane species.

Oenomaus, eighth abdominal tergum: ventral view, anterior edge at bottom. 38 ♂ Oenomaus mancha (holotype, Ecuador) 39 ♂ Oenomaus lea (holotype, Peru) 40 ♂ Oenomaus mentirosa (holotype, Peru) 41 ♂ Oenomaus myrteana (holotype, Ecuador) 42 ♂ Oenomaus andi (holotype, Ecuador) 43 ♀ Oenomaus andi (paratype, Ecuador) 44 ♂ Oenomaus moseri (holotype, Brazil) 45 Male genitalia in lateral view: Oenomaus moseri (left) and Oenomaus morroensis holotypes. Diagnostic characters (1), (2) and (3) are explained in the text.

Distributions. 46 Oenomaus mancha, Oenomaus gwenish, Oenomaus myrteana, Oenomaus moseri 47 Oenomaus andi, Oenomaus lea, Oenomaus mentirosa.
The holotype male was landed on a leaf about 5 m above the ground at 11:00 hours. Males and females are attracted to traps baited with rotting fish (vouchers in RCB).
urn:lsid:zoobank.org:act:D35807B0-C3C3-4384-BB59-E294B3E06F97
http://species-id.net/wiki/Oenomaus_gwenish
Figs 3, 27, 46Panama: Darién, Serranía de Pirre, Cana, 7°55'57"N, 77°42'58"W, 1000 m. Serranía de Pirre at 1000 m was uncut wet lower montane forest in 1984. The only disturbance was a defunct gold mine camp and associated dirt runway at Cana.
Holotype ♀ (Fig. 3) labeled as “PANAMA: Darien: / Serrania de Pirre: / Cana: 1, 000 m / 5 January 1984 / Leg. G.B. Small” [rectangular, white, printed and handwritten], “GENITALIA NO. / 2011: 406♀ / C. FAYNEL” [rectangular, green, printed] “Holotype ♀ / Oenomaus gwenish / Robbins & Faynel, 2012” [rectangular, red, printed]. Deposited in USNM.
Female FW length: 20 mm (N = 1). Wing pattern (Fig. 3) and genitalia (Fig. 27) illustrated. The wing patterns of Oenomaus gwenish and Oenomaus mancha are distinguished from that of Oenomaus ortygnusby the black patch in the distal part of the VHW cell Sc+R1-Rs and by the absence of a black mark in VFW cell Costa-Sc. However, the ventral wing pattern of Oenomaus gwenish differs from that of Oenomaus mancha by (1) the lack of a black band crossing the VFW discal cell, (2) the absence of a black spot in VFW cell M3-Cu1, and (3) the presence of two black spots of equal size along VHW veins mdc and ldc instead of a single large black spot at vein mdc with no mark or a faint vestigial mark at vein ldc. Thefemalegenitalia of Oenomaus gwenish (Fig. 27) are similar to those of Oenomaus mancha.
We hesitated to describe this species because we cannot assess its intraspecific variation. However, the series of 10 females of Oenomaus mancha show little variation in the traits that distinguish them from the holotype of Oenomaus gwenish. For this reason, a hypothesis of specific distinctness is better supported than a hypothesis of geographical variation.
The holotype of Oenomaus gwenish is a unique and distinctive female, for which reason it gives us great pleasure to name this species for entomologist Dr. Jennifer (Gwen) Shlichta. The name is a feminine noun in apposition.
Oenomaus gwenish is probably a lower montane species, so far known only from wet forest at 1000 m elevation in Darién, Panama (Fig. 46). While Oenomaus gwenish and Oenomaus ortygnus are both known from Panama, we do not know if they are sympatric.
urn:lsid:zoobank.org:act:A8315354-6FFF-4C71-AEE5-92925AE8833B
http://species-id.net/wiki/Oenomaus_lea
Figs 4, 21, 39, 47Ecuador: Napo, Misahuallí Rd, Latas Grande, 7.7 km E Puerto Napo, 1°02.0'S, 77°44.1'W, 470 m. The holotype was collected along the road from Tena to Misahuallí, which in 1991 was a patchwork of “fincas” and remnant second growth forest.
Holotype ♂ (Fig. 4) labeled as “ECUADOR Napo / Misahualli Rd. 470m / Latas Grande / 9 Nov.’ 91 / S. S. Nicolay” [rectangular, white, printed and handwritten], “USNM ENT 00180040” [rectangular, white, printed], “GENITALIA NO. / 2011: 408♂ / C. FAYNEL” [rectangular, green, printed] “Holotype ♂ / Oenomaus lea / Faynel & Robbins, 2012” [rectangular, red, printed]. Deposited in USNM.
Paratypes: Ecuador. 1 ♂: La Merced on Río Pastaza below Baños, Alt. 4000 ft. [= 1220 m], W. J. Coxey, III.1930, A.N.S. Lot 217, genitalia NO. 1992: 12 ♂ R.K. Robbins (ANSP). Peru. 3 ♂: UC, Pucallpa, 200 m, X.2007, Michael Büche leg. (CF); LO, Contamana, Río Ucayali, 300 m, 7°19'S, 74°48'W, IX.2010, leg J. Ramírez (LYD); SM, Juanjui, upper Huallaga River, IX. 1934, collector G. Klug, collection E.I. Huntington NO. 1055 (AMNH).
Male FW length: 18.2 mm (SD = 0.8, N = 4). Wing pattern (Fig. 4) and genitalia (Fig. 21) illustrated. Oenomaus lea and Oenomaus atesa (Hewitson, 1867)are the only two Eumaeini sharing the striking underside wing pattern with two transverse brown bands on the VHW. However, males of Oenomaus lea differ from males of Oenomaus atesa (Figs 5, 6 in
This species is named for Léa Faynel, daughter of Christophe Faynel. The name is a feminine noun in apposition.
Oenomaus lea occurs in wet lowland forest up to 1200 m elevation in eastern Ecuador and eastern Peru (Fig. 47).
urn:lsid:zoobank.org:act:B8EBB664-61DA-4178-91C8-52D2D9BFF0AC
http://species-id.net/wiki/Oenomaus_myrteana
Figs 5, 6, 22, 28, 41, 46Ecuador: Morona Santiago, Santiago (Hill North of Town), 3°02.3'S, 78°00.3'W, 350 m. The holotype was collected in wet secondary forest in the low hills on the north edge of Santiago.
Holotype ♂ (Fig. 5) labeled as “ECUADOR: Morona Santiago / Santiago (Hill North of Town) / 3°02.3'S, 78°00.3'W 350 m / 20 September 2004 / Robert C. Busby, leg.” [rectangular, white, printed], “GENITALIA NO. / 2003: 35♂ / R.K. ROBBINS” [rectangular, green, printed] “Holotype ♂ / Oenomaus myrteana / Busby, Robbins & Faynel, 2012” [rectangular, red, printed]. Deposited in USNM.
Paratypes: Ecuador. 1 ♂: Morona Santiago, Santiago (Hill North of Town), 3°2.3'S, 78°0.3'W, 350 m, 20.IX.2006, Robert C. Busby leg. (RCB) ; 1♀: Morona-Santiago Province, 1.8 km Santiago-Puerto Morona Rd., 3°2.4'S, 77°59.7'W, 300–350 m, 20.IX.2006, D.H. Ahrenholz & Robert C. Busby leg., gen. prep. CF n°415 (RCB) (Fig. 6). Brazil. 2 ♂: RO, Cacaulândia, 1–5.IX.1997, E. Furtado & A. Moser leg., gen. prep. CF n°442 (MC 250); RO, Candeias do Jamari, Rio Preto, 27–31.VIII.1997, E. Furtado & A. Moser leg., gen. prep. CF n°443 (MC 251).
Male FW length: 12.8 mm (SD = 0.1, N = 3). Female FW length: 12.1 mm (N = 1). Wing pattern (Figs 5, 6) and genitalia (Figs 22, 28) illustrated. Oenomaus myrteana has a conspicuous round white spot in VHW cell Sc+R1-Rs, which is similar to those species of Oenomaus with a ventral wing patterns similar to that of Oenomaus atena. However, Oenomaus myrteana lacks the inclined white median line of the VFW, which is characteristic of species with the Oenomaus atena wing pattern. Instead, Oenomaus myrteana has a vertical, distally displaced postmedian line of white dashes, inwardly bordered by black. This character appears to be unique among Oenomaus species. In addition, Oenomaus myrteana may have a few red-orange scales in VHW cell Cu1-Cu2. This red-orange cubital spot is uniformly lacking in other Oenomaus and Porthecla. The male genitalia of Oenomaus myrteana are very similar to those of Oenomaus nigra, which has an “atena-like” ventral wing pattern. The female genitalia of Oenomaus myrteana are similar to those Oenomaus that have a bifid posterior end of the ductus bursae and a signa with a two pointed spine in the middle of the corpus bursae.
The ventral wing pattern of Oenomaus myrteana is superficially similar to those of Enos myrtea (Hewitson) and Allosmaitia myrtusa (Hewitson), but in these genera, males lack a scent pad on the DFW. The genitalia of Oenomaus myrteana, as noted, are typical of Oenomaus.
The name Oenomaus myrteana is intended to highlight the striking resemblance between the ventral hindwing of this species and that of Enos myrtea (Hewitson). The name is a feminine noun in apposition.
Oenomaus myrteana occurs in lowland wet forest from eastern Ecuador to western Brazil (Rondônia) (Fig. 46). Busby observed males in Ecuador low in the understory at 11:00 hours. This species and Enos myrtea have been found at the same site.
urn:lsid:zoobank.org:act:FDD83214-61B2-4B8A-B468-984D9DA0560D
http://species-id.net/wiki/Oenomaus_mentirosa
Figs 7, 23, 40 , 47Peru: Madre De Dios, Río La Torre, Tambopata Res., 12°50'13"S, 69°17'35"W, 300 m. Tambopata is at the mouth of the Río La Torre. In 1986 there was a lodge and a network of trails through uncut wet lowland forest. The holotype was collected during the transition between the dry and wet seasons when butterfly abundance and diversity generally peak.
Holotype ♂ (Fig. 7) labeled as “PERU Madre De Dios / Rio La Torre 300m / Tambopata Res. / 3 Oct.’ 86 / S. S. Nicolay” [rectangular, white, printed and handwritten], “GENITALIA NO. / 2011: 409♂ / C. FAYNEL” [rectangular, green, printed] “Holotype ♂ / Oenomaus mentirosa / Faynel & Robbins, 2012” [rectangular, red, printed]. Deposited in USNM.
Paratypes: Peru. 4 ♂: LO, km 28, Iquitos-Nauta, 180 m, 0359/7326, 30.X.2003, J.J. Ramírez leg. (MUSM) ; MD, Boca Río La Torre, 300 m, 17.IX.1984, I. Bohórquez leg., Genitalia NO. 1992: 47♂ R.K. Robbins (MUSM); MD, Boca Río La Torre, 300 m, 27.X.1981, G. Lamas et al., Genitalia NO. 1992: 48♂ R.K. Robbins (MUSM) ; MD, Tambopata Reserve, 12°50'S, 69°17'W, 300 m, 27.X.1990, Leg. R. Robbins, Genitalia NO. 1992: 39♂ R.K. Robbins (USNM ENT 00180049).
Male FW length: 14.9 mm (SD = 0.3, N = 2). Wing pattern (Fig. 7) and genitalia (Fig. 23) illustrated. The ventral wing pattern of Oenomaus mentirosa is very similar to some species of the Porthecla gemma group (
The name of this species comes from the Spanish word ‘mentirosa’, which means a feminine liar. We picked this name because the underside wing pattern resembles that of Porthecla gemma (Druce) and Porthecla minyia (Druce), but this resemblance appears to be a false indicator of relationship. We treat the name as a feminine noun in apposition.
Oenomaus mentirosa is known from lowland wet forest in Amazonian Peru (Fig. 47).
Resemblance of the ventral wing patterns of Oenomaus mentirosa and Porthecla gemma/Porthecla minyia was noted in the etymology. Adults of all three species fly in the same habitats at the same time of year in the vicinity of Puerto Maldonado, Peru.
urn:lsid:zoobank.org:act:C6C09BC9-F0B9-4E7A-8951-0D9923BE9E73
http://species-id.net/wiki/Oenomaus_andi
Figs 8, 9, 24, 29, 42, 43, 47Ecuador: Zamora Chinchipe Prov., Zamora (ridge W. of town), 4°04.5'S, 78°58.1'W, 1450 m. The ridge west of Zamora rises rather sharply from the city and is accessed by a dirt road which goes up to about 1300 m. The top of the ridge is still forested but a significant part of the surrounding land has been turned into pasture.
Holotype ♂ (Fig. 8) labeled as “ECUADOR / Zamora Chinchipe Prov. / Zamora (ridge W. of town) / 18. ix. 2000 (1450m) / leg. Robert C. Busby” [rectangular, white, printed], “GENITALIA NO. / 2009: 344♂ / C. FAYNEL” [rectangular, green, printed] “Holotype ♂ / Oenomaus andi / Busby & Faynel, 2012” [rectangular, red, printed]. Deposited in USNM.
Paratypes: Ecuador. 3♀: Morona-Santiago, 1 km E Río Abanico, 1600 m, 2°15.4'S; 78°11.7'W, 15.IX.2003, Robert C. Busby leg., gen. prep. CF n°416 (RCB) (Fig. 9) ; Morona-Santiago, 14 km W. of Macas, 1600m, 28.IX.1998, Río Abanico, leg. Robert C. Busby (RCB); Zamora Chinchipe, Zamora (ridge W. of town), 4°04.5'S, 78°58.1'W, 1450 m, 06.X.2007, D. H. Ahrenholz, R. C. Busby leg. (RCB).
Bolivia. 1♀: La Paz, Nor Yungas, Caranavi, 1500 m, XII. 2004, gen. prep. CF n°445 (MC 253).
Male FW length: 16.3 mm (N = 1). Female FW length: 16.7 mm (SD = 0.8, N = 2). Wing pattern (Figs 8, 9) and genitalia (Figs 24, 29) illustrated. The ventral wing pattern of Oenomaus andi is similar to that of many other Oenomaus, but this species is distinguished by (1) a white spot on the basal side of VHW cell Rs-M1, (2) an elongated double valvae of equal size, (3) a large posterior part of the saccus in lateral view, (4) a swollen terminal end of the penis, and (5) modified anterior and posterior edges of the male 8th tergum (detailed under remarks).
This species is named for Andrea (Andi) Busby, wife of Robert Busby, in appreciation for her long standing support of his research. The name is a feminine noun in apposition.
Valvae structure in Oenomaus andi is very similar to that found in Oenomaus gaia Faynel, suggesting that this new species belongs to the Oenomaus cortica subgroup (as characterized by
Oenomaus andi is a species of montane forest (> 1300 m) that is recorded from Ecuador to Bolivia (Fig. 47).
A male and two females were attracted to traps baited with rotting fish (vouchers in RCB).
urn:lsid:zoobank.org:act:DC5D1BF6-2149-4BB9-802D-58C27D11AB38
http://species-id.net/wiki/Oenomaus_moseri
Figs 10, 11, 25, 30, 44, 45, 46Brazil: SC, Joinville, 26°19'39"S, 48°57'38"W, 10–200 m. Miers collected butterflies for decades in the wet lowland forests around Joinville, where he lived. His favorite collecting spot was a hill that he called “Serrinha” (little hill in Portuguese) in Vila Nova, approximately 10 km west, south-west of the center of Joinville. According to DZUP butterfly curator Olaf Mielke, specimens collected on Serrinha, including the holotype, have an elevation label 10–200 m, which distinguishes them from those specimens collected in other parts of the Joinville area.
Holotype ♂ (Fig. 10): Brazil, SC, Joinville, 10–200 m, 2.IV.1978, Miers leg., gen. prep. CF n°218, DZ 10.065, CF-LYC-012 (DZUP).
Paratypes: Brazil. 12 ♂: SC, Joinville, 200 m, 26°19'S, 48°58'W, 20.V.1971, H.Miers leg., gen. prep. CF n°444 (MC 252) ; SC, São Bento do Sul, 600 m, 25.IV.2002, Moser & Rank leg., gen. A. Moser, n°234 (MC 034) ; SC, Joinville, 200 m, 5.II.1993, A. Moser leg., gen. A. Moser, n°226 (MC 032); SC, Joinville, 200 m, 5.II.1993, A. Moser leg., gen. A. Moser n°233 (MC 033) ; SC, Joinville, 10–200 m, 8.XII.1983, Leg. H. Miers, R.K. Robbins collection (USNM) ; SC, Joinville, 10–200 m, 6.I.1984, Leg. H. Miers, R.K. Robbins collection (USNM) ; PR, Ponta Grossa, Buraco do Padre, 900 m, 20.II.2009, Carlos Mielke leg., CF-LYC-063 (CF) ; SP, Serra do Japi, 110[0m], 23°15'S, 46°54'W, 12.IV.1991, Robbins & K. Brown, territorial behavior at 14:23, Genitalia NO. 1992: 27♂ R.K. Robbins (USNM) ; SP, Serra do Japi, 110[0]m, 23°15'S, 46°54'W, 12.IV.1991, Robbins & K. Brown, territorial behavior at 14:48 (USNM) ; SP, Serra do Japi, 800–1250 m, 23°12'S, 47°02'W, 23°17'S, 46°53'W, 25.III.1990, Leg. K. Brown (x2, USNM) ; SP, Serra do Japi, 800–1250 m, 23°12'S, 47°02'W, 23°17'S, 46°53'W, 28.III.1990, Leg. K. Brown (USNM) ; RJ, Petrópolis, 6.I.1980, Leg. C. Callaghan, R.K. Robbins collection, Genitalia NO. 1992: 79♂ R.K. Robbins (USNM ENT 00180045). 1♀: SC, Joinville, 10–200 m, 9.III.1973, Leg. H. Miers, R.K. Robbins collection, gen. prep. CF n°410 (USNM) (Fig. 11).
Male FW length: 16.1 mm (SD = 0.9, N = 8). Female FW length: 15.7 mm (N = 1). Wing pattern (Figs 10, 11) and genitalia (Figs 25, 30) illustrated. The adult wing pattern of Oenomaus moseri is similar to that of the sympatric Oenomaus morroensis Faynel & Moser and to that of Oenomaus cyanovenata (D’Abrera); the species with which it was previously confused (
Preliminary data on divergence of “barcode” DNA sequence data is consistent with morphology. The divergence among three individuals of Oenomaus moseri (CF-LYC-012 & CF-LYC-063) is 0%, among four individuals of Oenomaus cyanovenata (CF-LYC-047, CF-LYC-048, & CF-LYC-049) is 0%. In contrast, the divergence between Oenomaus moseri and Oenomaus cyanovenata is more than 4% and between two Oenomaus moseri and a paratype of Oenomaus morroensis (CF-LYC-015) is more than 5%.
It is with great pleasure that we name this distinctive species for our good friend and collaborator Alfred Moser. Alfred lives in Rio Grande do Sul and has made prodigious contributions to the knowledge of Lepidoptera from southern Brazil, including co-authoring papers on the taxonomy of Oenomaus and Porthecla (
Robbins observed two males of Oenomaus moseri exhibiting territorial behavior on a hill top from 14:23 hours to 14:48 hours at Serra do Japi (SP, Brazil) on 12 April 1991 (vouchers in USNM). A male of Oenomaus moseri was reared by Hipólito Ferreira Paulino Neto in Itirapina, SP, Brazil on Duguetia furfuracea (A. St. Hil) Benth. and Hook. f. (Annonaceae), a plant of frequent occurrence in the cerrado. We identified the male from a digital image and from the locality where it was reared. However, it is possible that it is a male of Oenomaus morroensis, even though this species is not known to occur as far north as São Paulo.
Oenomaus moseri occurs in lowland and lower montane forest in southern Brazil (Fig. 46).
For each of the 21 previously described Oenomaus species, we give distribution, habitat, and remarks. We then note, where relevant, new information on taxonomy, intraspecific variation, behavior/biology, associated females, and COI DNA sequences. The species are treated in alphabetical order. Oenomaus curiosa and Oenomaus melleus are included in this section, even though their generic placement is yet unresolved (
Oenomaus ambiguus is a poorly known, lowland species whose ventral wing pattern is virtually indistinguishable from those of Oenomaus cortica and Oenomaus gaia. It has been recorded from French Guiana and Amazonian Peru. The previous record from Amazonas, Brazil (
. 1♂: Bas Maroni, Guyane Française, gen. prep. CF n°319 (MNHN H-452). Peru.– 1♂: MD, Río La Torre, 300 m, Tambopata Res., 27.IX.1987, S.S. Nicolay, gen. prep. CF n°404 (USNM).
Unknown.
The paratype from Peru has been barcoded (CF-LYC-025), and the sequence is 2–3% divergent from those of Oenomaus cortica, Oenomaus gaia and Oenomaus morroensis (Table 1).
Oenomaus atena is a widely distributed lowland species that is reliably recorded from Costa Rica, Panama, western Ecuador, French Guiana, Venezuela, Peru, and Brazil (AM, MT). Most species with an “atena-like” ventral wing pattern have historically been identified as Oenomaus atena, which means that virtually all literature records for Oenomaus atena from before 2005 are unreliable.
Costa Rica.– 1♂: Guápiles, 850 ft. alt., June, Schaus and Barnes coll., genitalia on slide X-10-1946, W.D.F. 2333 (USNM). Panama.– 1♂: Cerro Campana, 2000’, XII-22-1963, G.B. Small, Genitalia 1992: 15♂ R.K. Robbins (USNM). Ecuador.– 1♂: Esmeraldas, 25 km San Lorenzo-Lita Road, 1°10.0'N, 78°40.0'W, 100 m, VI.2003, San Francisco, R. Aldas & Robert C. Busby leg., gen. prep. CF n°343 (RCB). Peru.– 2♂: MD, 30 km S.W. Pto. Maldonado, 300 m, 20.X.1983, S.S. Nicolay, Genitalia 1992: 16♂ R.K. Robbins (USNM); MD, 10 km north Puerto Maldonado, 200 m, 12°36'S, 69°11'W, 26–30.XI.1993, leg. C. Tello (USNM).
The female of this species was determined by a pair collected in copula and was illustrated by
Three specimens of Oenomaus atena have been barcoded, including a male from Peru (LO) (CF-LYC-084) and two females from French Guiana (CF-LYC-054 and CF-LYC-057). The latter two have the same brown dorsal wing pattern, ventral wing pattern, and genitalia as the female of Oenomaus atena found in copula. The three barcodes show 0.4% divergence.
Oenomaus atesa is a widespread species that has been recorded from Mexico, Panama, western Ecuador, French Guiana, Venezuela, Colombia, eastern Ecuador, Peru, and Brazil (AM, DF, MG, RJ, SP, SC). The vast majority of museum specimens were collected in the lowlands, but males have also been found at 1375–1700 m in western Ecuador and at 2200 m in western Colombia (
Venezuela.– 1♀: Venezuela, Aragua, Rancho Grande, 1100 m, 29.V.1985, S.S. Nicolay leg., gen. prep. CF n°404 (USNM). Ecuador.– 2♂: Pichincha 5 km Nanegal-García Moreno Rd, 0°09.2'N, 78°39.4'W, 4.VI.2008, 1375–1700 m, Robert C. Busby leg., gen. prep. CF n°340 (RCB);Napo Province, 14 km S of Tena, 17–18.X.1996, 600 m, Robert C. Busby leg., gen. prep. CF n°347 (RCB). 1♀: Río Chuchuví, Lita vers San Lorenzo km12, 700 m (provincia de Esmeraldas), VIII.2001, Euclides Aldaz leg. (PB). Peru.– 1♀: LO, 180 m, San Salvador, 5 km NNW Contamana, 08°19'S, 75°01'W, 27.XI.2002, D.H. Ahrenholz leg., gen. prep. CF n°403 (USNM). Brazil.– 1♂: DF, Parque do Gama, 950 m, 14.V.1969, S.S. Nicolay leg., gen. prep. CF n°405 (USNM ENT 00180586).
Despite substantive geographical variation in Oenomaus atesa, we lack sufficient material to determine if this variation might represent more than one species. Females from Venezuela and western Ecuador have more extensive dorsal blue and a somewhat lighter color than females from Panama, French Guiana, eastern Ecuador, and Peru. In addition, males from western Ecuador have more blue on the dorsal forewings than males from eastern Ecuador. However, this variation is small compared to that between males of Oenomaus atesa and Oenomaus lea. For example, the forewing dorsal blue area never reaches the cells from vein R3 to Cu1 as it does in Oenomaus lea. Structure of the female genitalia also varies geographically. Females from Venezuela and Peru have two processes at the posterior end of the lamella postvaginalis while a female from French Guiana had none (see
Males exhibited territorial behavior on a hilltop in Panama (Canal Area, Gamboa, Cerro Pelado) from 13:15 to 15:30 hours (19 males, 10 different days during the months of January, February, March, April, August, September, October, and December, 15 vouchers in USNM). Similarly, territorial males on a hilltop in Brazil (Santa Catarina, Villa Nova, Serrinha) were observed from 14:40 to 14:55 hours (3 males, March, 3 vouchers in USNM).
Females are associated with males by their ventral wing pattern, which is unique among the Eumaeini. Characters were noted for distinguishing the ventral wing pattern of Oenomaus atesa from that of Oenomaus lea.
One male of Oenomaus atesa from French Guiana has been barcoded (CF-LYC-003).
French Guiana.– 1♂: Guyane, no date, S. Fernandez leg., CF-LYC-033 (CF). 1♀: Montagne des Singes, 5°07'N, 52°69'W, 5.XII.2007, T. Rosant leg., gen. prep. CF n°440, CF-LYC-034 (CF) (Fig. 14).
Female. We associate a female (Figs 14, 31) which has the same ventral wing pattern as the male, which occurs in French Guiana (as do the known males), and which has a very similar COI DNA sequence to that of the males.
Divergence among the three known specimens is 0.2%.
This species occurs in wet lowland forest and is recorded from Panama, Guyana, Peru, and Brazil (PA, AM). Oenomaus cortica, Oenomaus gaia, and Oenomaus ambiguus have very similar wing patterns, but their genitalic structures are distinct.
Panama.– 1♂: Gatún, C. Z., 2.V.1970, G.B. Small leg., Genitalia 1992: 13♂ R.K. Robbins (USNM). Guyana.– 1♂: Potaro Riv., VIII-IX.1902, C.B. Roberts, Genitalia 1992: 74♂ R.K. Robbins (FSMC). Peru.– 1♂: MD, Parque Manu, Pakitza 340 m, 11°55'48"S, 71°15'18"W, 14.X.1991, Leg. R. Robbins, Genitalia No. 1996: 3♂ R.K. Robbins (USNM ENT 00180044).
The male from Panama has the posterior edge of its 8th tergum more deeply incised than in others.
Unknown. A female paratype of Oenomaus cortica from Espírito Santo, Brazil was illustrated in
Two males from Brazil, Pará have been sequenced (CF-LYC-051 and CF-LYC-052) and show 0.6% divergence.
Oenomaus curiosa is a species of wet lowland forest that is recorded from French Guiana, Peru (LO, MD), and Brazil (RO).
Peru.– 2♂: MD, 300 m, 30 km S. W. Pto Maldonado, 26.X.1983, S.S. Nicolay, Genitalia No. 1992: 25♂ R.K. Robbins (USNM); LO, 120 m, Pebas, river Amazonas, 03°19'S, 71°51'W, II. 2011, Ramírez leg. (CF). Brazil.– 1♂: RO, 62 km SW Ariquemes, Línea 20, lot 21, 23, 25 (Fazenda Rancho Grande), 11.X.1993, AVZ Brower, gen. prep. CF n°433 (OSAC).
Unknown
Two males from French Guiana, including one of the paratypes, have been sequenced (CF-LYC-036 and CF-LYC-037) and show 0.8% divergence.
A species of very wet lowland forest, it has been recorded from Costa Rica, Panama, French Guiana, Venezuela, Bolivia, and Brazil (PA, AM). The previous record for Brazil (SC) was incorrect; this specimen is now treated as Oenomaus moseri.
Costa Rica.– 1♂: Guápiles, 850 ft. alt., Schaus and Barnes coll., Genitalia 1992: 76♂ R.K. Robbins (USNM); 2♀: Area de Conservación Guanacaste, voucher: D.H. Janzen & W. Hallwachs 97-SRNP-62841.1, Genitalia 2009: 30♀ R.K. Robbins (USNM) (Fig. 16); 97-SRNP-6283. Panama.– 1♂: Colón, Piña, 100 m, 9.IV.1971, H.L. King, genitalia slide/vial #4710, prep. S.S. Nicolay (USNM). French Guiana.– 3♀: Roura, Route de Kaw - PK 16, 18.VII.2004, C. Faynel leg., CF-LYC-053 (CF); Roura, Route de Kaw, 26.I.2005, J.Y. Gallard leg., gen. prep. CF n°441, CF-LYC-055 (CF) (Fig. 17); Roura, Route de Kaw - PK 8, 20.XII.2001, J.Y. Gallard leg., CF-LYC-056 (CF). Brazil.– 3♀ : PA, Santo Antônio do Tauá, Reserva Sonho Azul, 1°15'S, 48°03'W, 12.VI.2009, P. & J. Jauffret leg., CF-LYC-059 (CF); PA, Santo Antônio do Tauá, Reserva Sonho Azul, 1°15'S, 48°03'W, 3.VIII.2009, P. & J. Jauffret leg., CF-LYC-060 (CF); PA, Santo Antônio do Tauá, Reserva Sonho Azul, 1°15'S, 48°03'W, 8.V.2009, P. & J. Jauffret leg., CF-LYC-061 (CF).
Females from French Guiana and Brazil, Pará (Fig. 17) are uniformly brown on the dorsal wing surface while the female from Costa Rica (Fig. 16) has the basal parts of both wings blue. Their genitalia, however, are uniform. Additionally, their COI DNA sequences are similar. This geographic variability is similar to that in Oenomaus taua.
Two females were reared in Costa Rica (97-SRNP-62841.1 and 97-SRNP-6283) from Guatteria verrucosa R.E. Fr. (Annonaceae) (adult vouchers in USNM). Details of the rearing records along with images of the immatures can be found in
Females of this species (Figs 16, 17, 32, 33) have the same ventral wing pattern as males, occur at the same localities, and have similar COI DNA sequences. A female paratype of Oenomaus cyanovenata from Pará, Brazil was designated and illustrated in
Four males and seven females from French Guiana and Brazil, Pará were barcoded. One male (CF-LYC-046) is 6.7% divergent from the other three males, but its sequence is identical with that from a male of Oenomaus magnus (CF-LYC-020). Potential explanations for this result range from contamination to biologically significant, but until we have additional information, we omit this male from the following results. Divergence among the 10 other specimens of Oenomaus cyanovenata was 0.1%. The reared females from Costa Rica, which were barcoded in another project, are 0.4% divergent from the South American specimens.
This species was described from one Brazilian (AM) male, which is the only known specimen. As noted, its genitalia are similar to those of Oenomaus mentirosa, but it has a distinctly different ventral wing pattern.
Unknown.
This species occurs in lowland and lower montane habitats with wet or deciduous forest. It is recorded from eastern Ecuador and Brazil (AM, MT, DF, GO, PR).
Ecuador.– 1♂: Pastaza Province, 45 km Puyo-Arajuno Rd., 1000 m, 26.IX.1999, Robert C. Busby leg., gen. prep. CF n°342 (RCB). Brazil.– 2♂: GO, 163 km W. Jataí S. Rita Araguaia, 850 m, 29.V.1969, S.S. Nicolay, genitalia slide/vial #4367, prep. S.S. Nicolay (USNM); PR, Highlands, 24.XI.1934, coll. Karl Schmitt, E.I. Huntington, Genitalia 1992: 19♂ R.K. Robbins (AMNH).
Described by
This species occurs in wet and dry lowland forest. It has been recorded from Panama, French Guiana, Venezuela, eastern Ecuador, Peru (LO, SM, UC, MD) and Brazil (PA, AM, RO, MT, GO). This species, Oenomaus floreus, and maybe Oenomaus griseus occur in drier forest than other species with an “atena-like” ventral wing pattern.
Panama.– 1♂: Los Ríos, C. Z., 15.XII.1964, S.S. Nicolay leg., Genitalia 1992: 73♂ R.K. Robbins (USNM ENT 00180046). 2♀: Los Ríos, C. Z., 27.I.1965, S.S. Nicolay, gen. prep. CF n°430 (USNM) (Fig. 15); Los Ríos, C. Z., 19.XII.1964, G.B. Small, gen. prep. CF n°431 (USNM). Ecuador.– 1♂: Morona-Santiago 15 km S Gualaquiza, 850 m, 3°27.6'S, 78°33.1'W, 27.IX.2000, Robert C. Busby leg. (RCB). Peru.– 1♂: MD, Parque Manu, Pakitza 340 m, 11°55'48"S, 71°15'18"W, 15.X.1991, Leg. M. Casagrande, Genitalia No. 1992: 38♂ R.K. Robbins (USNM). Brazil.– 2♂: PA, Obidos, IX.1930, Ex coll. Le Moult, Genitalia No. 1992: 75♂ R.K. Robbins; PA, Santo Antônio do Tauá, Reserva Sonho Azul, 1°15'S, 48°03'W, 16.VII.2003, P. & J. Jauffret leg., CF-LYC-072 (CF); RO, 62 km SW Ariquemes, Línea 20, lot 21, 23, 25 (Fazenda Rancho Grande), 11.X.1993, AVZ Brower, gen. prep. CF n°411 (OSAC); GO, Pirenópolis, 820 m, 15°49'S, 48°59'W, E. Emery leg. (MC 255).
Four males in the USNM were collected on hills on Los Ríos hill (approximately 9°00'32"N, 79°35'34"W) and in Cocolí (approximately 8°58'46"N, 79°35'59"W), Canal Area, Panama. These areas are drier (<2 m annual precipitation,
We also associate a female from Brazil, Pará (CF-LYC-072) with a male of Oenomaus gaia from French Guiana because they have the same ventral wing pattern and have similar barcode sequences (0.2%).
As noted previously, interspecific variation in the barcode sequences of Oenomaus ambiguus, Oenomaus cortica, Oenomaus gaia, Oenomaus morroensis is less than 2%, in contrast to interspecific divergences among other species in Oenomaus. For example males of Oenomaus gaia (CF-LYC-023) and Oenomaus cortica (CF-LYC-052) are 0.8% divergent. Another male of Oenomaus gaia (CF-LYC-024) and Oenomaus morroensis (CF-LYC-015) are 1.1% divergent.
This species is a relatively uncommon inhabitant of lower montane forest in southern Brazil, so far known only from the state of Santa Catarina. Previously, it was known only from the male holotype, which lacks locality data.
Brazil.– 5♂: SC, Highlands near Massaranduba-Blumenau, Collection E.I. Huntington No. 1009 (AMNH, x4); SC, Highlands near Massaranduba-Blumenau, Collection E.I. Huntington No. 1009, genitalia slide/vial #4705, prep. S.S. Nicolay, Allyn Museum Photo No. 022078-7, 8 VI (AMNH) . 2♀: SC, Rio Vermelho, São Bento do Sul, 10.III.1973, leg. Rank, gen. prep. CF n°414 (USNM ENT 00180041); SC, Highlands near Massaranduba-Blumenau, Collection E.I. Huntington No. 1009, genitalia slide/vial #4707, prep. S.S. Nicolay, Allyn Museum Photo No. 022078-9, 10 VI (AMNH).
The female (Figs 12, 35) occurs in the same habitat as the male and has the same ventral wing pattern.
This species appears to be endemic to Brazil’s central plateau (DF).
Unknown.
This widespread South American species occurs in wet and dry lowland forests. It is recorded from French Guiana, Colombia, eastern Ecuador, Peru, Bolivia, and Brazil (AM, MG).
Colombia.– 1♀: Muzo, 400b. 800 m, coll. Fassl (SMF). Ecuador.– 2♀: Morona-Santiago, Santiago (Hill North of Town), 3°02.3'S, 78°00.3'W, 350 m, 20.IX.2010, Robert C. Busby leg. (RCB); 27 km Santiago-Puerto Moreno Rd., 2°56.4'S, 77°49.5'W, 500–550 m, 17 IX 2005, Robert C. Busby, leg. (RCB). Peru.– 1♀: JU, Aldea, 600–700 m, 1054/7455, 23.VIII.2003, J.J. Ramírez (MUSM). Brazil. – 2♂: MG, km 500 Belo Horizonte-Brasília, Hwy, 11.IV.1973, C. Callaghan, genitalia slide/vial #4737, prep. S.S. Nicolay (USNM); AM, Rio Amazonas, Vila Nova (ca. Tonantins, 0252S/6748), 100 m, IX.1993, M. Büche leg. (MUSM). Bolivia.– 1♂: Las Juntas, XII. 1913, Steinbach Acc. 5045, genitalia slide/vial #4743, prep. S.S. Nicolay (CMNH).
The distinctive ventral wing pattern of this species allows identification of the female.
Sequences from a Brazilian male (CF-LYC-006) and French Guiana female paratype (CF-LYC-007) diverge 3.0%.
This species inhabits wet lowland forest. It is recorded from French Guiana, eastern Ecuador, Peru, Bolivia, and Brazil (PA, MT).
Ecuador.– 1♂: Pastaza Province, 32 km S. of Puyo, 20–21.X.1996, 1000 m, Robert C. Busby leg., gen. prep. CF n°346 (RCB). Peru.– 1♂: LO, Agua Blanca, 0356/7328, 130 m, 10.XI.2005, J.J. Ramírez (MUSM). 2♀: JU, vic. Satipo, c. 800 m, Villa Esperanza, c. 11°16'S, 74°15'W, V.1983, leg. M. Callegari (USNM) ; LO, Cerros de Contamana, El Indio, 200 m, 10.IX.1986, P. Hocking (MUSM). Bolivia.– 1♂: Río Songo, 750 m, coll. Fassl, Genitalia No. 2002: 5♂ R.K. Robbins (SMF).
As noted by
Females were associated with males by their characteristic ventral wing pattern (
This is a poorly understood species that occurs in South American lowland forest. It has been recorded from French Guiana, Peru, Bolivia, and Brazil (AM, MT).
French Guiana.– 1♀: Approuague - Mapaou, 4°31'N, 52°13'W, 29. XII. 2008, S. Fernandez leg. (CF) (Fig. 13). Peru.– 1♀: SM, Upper Huallaga Valley, V-VI 2000, Purch. Thorne 7/01, gen. prep. CF n°428 (USNM). Brazil.– 1♂: MT, Diamantino, Alto Rio Arinos, 5.X.1998, E. Furtado leg., gen. prep. CF n°446 (MC 254). Bolivia.– 1♀: “Thecla melleus Drc.♀, Buenavista 750 m, Bolivia Steinbach., Modt. 22/2 1927 af, qui Steinbach Bolivia, Coll. C.S. Larsen, Faaborg, gen. prep. CF n° 449 (MNHN).
We associate females from French Guiana (Figs 13, 36), Peru, and Bolivia with this species. They have the same ventral wing pattern, a similar geographic range, and limited COI DNA sequences are the same.
The sequence of a female from French Guiana (CF-LYC-039) is the same as (0% divergence) that of the male paratype from Peru (CF-LYC-020). This female is the first record of Oenomaus magnus from French Guiana.
This species occurs in wet lowland forest. It is recorded from Nicaragua, Costa Rica, French Guiana, Guyana, Venezuela, Colombia, Peru, Bolivia, and Brazil (PA, PE, ES).
Costa Rica.– 2♂: Heredia, prov. Finca La Selva, 3 km S. Puerto Viejo, 10°26'N, 84°01'W, 26.VII.1992, leg. H.A. Hespenheide (USNM) ; prov. Heredia, F. La Selva, 3 km S. Pto. Viejo, 10°26'N, 84°01'W, 5.IV.1985, H.A. Hespenheide (USNM). 1♀: Area de Conservación Guanacaste, voucher Janzen & Hallwachs #97-SRNP-62841, legs away for DNA, Genitalia 2009: 30♀ R.K. Robbins (USNM) (Fig. 20). Guyana.– 1♂: Region 7 Lower Cuyuni River nr. Arimu R. 100’, 6°34'N, 58°58'W, 2.IX.1991, leg. S. Fratello (USNM ENT 00180024). Brazil.– 1♂: ES, Itaguassú, IX.1971, Paulo César Elias, A.C. Allyn Acc. 1971-38, genitalia slide/vial #4700, prep. S.S. Nicolay (USNM). 1♀: PE, Recife, 5.I.1962, leg. Ebert (USNM).
The wing pattern of Oenomaus melleus is highly variable. The type from Colombia and two specimens from Nicaragua and Venezuela are relatively large (male FW length = 16.8 mm, SD = 1.3, N = 3). They have a white spot on the basal part of VHW cell Rs-M1, no reddish scales on the basal part of ventral wing, and a black spot in VHW cell Cu1-Cu2. The specimens from French Guiana, Guyana, Brazil (PA), Venezuela and Peru (UC) are smaller (male FW length = 14.1 mm, SD = 0.4, N = 5). They have a white spot on the basal part of VHW cell Rs-M1, reddish scales on the basal part of ventral wing, and no black spot in VHW cell Cu1-Cu2. The males from Costa Rica are also relatively small (male FW length = 14.7 mm, SD = 1.6, N = 3). They have no white spot on the basal part of VHW cell Rs-M1, no reddish scales on the basal part of ventral wing and a black spot in VHW cell Cu1-Cu2.
Described by
Described by
A small white spot on VHW cell Sc+R1-Rs that is displaced basally (
Although Oenomaus morroensis is unrecorded north of Santa Catarina, a reared male from São Paulo (see under Oenomaus moseri) could possibly be this species.
Unknown.
One paratype has been barcoded (CF-LYC-015). As already noted, this sequence is 5.0% divergent from the sympatric and superficially similar Oenomaus moseri.
This species occurs in wet lowland forest. It has been recorded from Peru and Brazil (AM). As noted, the genitalia of this species are similar to those of the newly described Oenomaus myrteana.
Peru.– 3♂: LO, Agua Blanca, 0356/7328, 130 m, 17.V.2004, J.J. Ramírez leg. (MUSM, x3). Brazil.– 1♂: AM, S. Paulo de Olivença, X.1983, Via Kesselring, Genitalia No. 1983: 133♂ R.K. Robbins (USNM ENT 00180054).
Unknown.
One male of Oenomaus nigra from Peru has been barcoded (CF-LYC-148).
This species occurs in many different habitats from sea level up to 1000 m. It is unique in the genus in that it is often found in highly disturbed habitats. It is the most common Oenomaus species in collections and has been recorded from the United States, Mexico, Guatemala, Honduras, Nicaragua, Costa Rica, Panama, French Guiana, Surinam, Guyana, Trinidad, Venezuela, Colombia, Ecuador, Peru, and many states throughout Brazil. As noted in the introduction, this species is a well-known pest of commercial Annonaceae.
The blacks spots on ventral wings vary in size and the blue on the dorsal wings vary from light cyan to dark purple. The “Thecla lauta Draudt” phenotype from western Mexico is smaller and duller than individuals from the remainder of its range.
Males were territorial on hilltops between 14:00–15:15 hours in Panama (Canal Area, hilltops in Paraíso, 7 males during June and August, 6 vouchers in USNM) and between 14:29 and 15:15 on Serrinha in Brazil (hilltop in Santa Catarina, Villa Nova, 200 m, 3 males in March, vouchers in USNM).
Both sexes are recognized by their ventral wing pattern, which is unique in the genus.
Sequences from a Peruvian male (CF-LYC-147) and a Mexican male (CF-LYC-146) diverge 1.5%.
This species occurs in wet lowland forest. It has been recorded from French Guiana and Brazil (PA, AM).
Brazil.– 1♂. Santarém, Amazons, A.H. Fassl, 3.IV.1920, gen. prep. CF n°317 (MNHN H-447).
Described by
This species is widespread in wet lowland forest. It is recorded from Guatemala, Panama, French Guiana, eastern Ecuador, Peru, and Brazil (PA, AM, RO). It is one of the more common species in the genus and mating pairs have been collected in Panama, Ecuador, and Brazil.
Guatemala.– 1♂: Cayuga, Sept., Schaus & Barnes coll., Genitalia No. 1992: 28♂ R.K. Robbins (USNM). Panama.– 2♂: Canal Zone, Gamboa, 5.I.1979, Leg. R. Robbins, in copula 15:00, Genitalia No. 1982: 125♂ R.K. Robbins (USNM ENT 00180050); Canal Zone, Summit, 17.III.1979, Leg. R. Robbins, in copula 15:00, gen. prep. CF n°423 (USNM). 2♀: Canal Zone, Gamboa, 5.I.1979, Leg. R. Robbins, in copula 15:00, Genitalia No. 1982: 126♀ R.K. Robbins (USNM ENT 00180051) (Fig. 18); Canal Zone, Summit, 17.III.1979, Leg. R. Robbins, in copula 15:00, gen. prep. CF n°424 (USNM). Ecuador.– 2♂: Napo, 14 km Tena-Puyo Road, 1°06.7'S, 77°46.9'W, 600 m, X.2010 (Apuya), I. Aldas & Robert C. Busby leg., gen. prep. CF n°418 (RCB);Napo Province, 14 km S. of Tena, 600 m, 17–18.X.1996 (Apuya), mating pair, Robert C. Busby leg., gen. prep. CF n°345 (RCB). 1♀: Napo Province, 14 km S. of Tena, 600 m, 17–18.X.1996 (Apuya), mating pair, Robert C. Busby leg., gen. prep. CF n°417 (RCB) (Fig. 19). Brazil.– 1♂: RO, 160–350 m, vic. Cacaulândia, 10°32'S, 62°48'W, 19.X.1991, in copula, Leg. J. MacDonald, gen. prep. CF n°412 (USNM). 1♀: RO, 160–350 m, vic. Cacaulândia, 10°32'S, 62°48'W, 19.X.1991, in copula, Leg. J. MacDonald, gen. prep. CF n°413 (USNM); PA, Santo Antônio do Tauá, Reserva Sonho Azul, 1°15'S, 48°03'W, 2.III.2010, P. & J. Jauffret leg. (CF).
Females from Brazil and Ecuador (Fig. 19) are uniformly brown on the dorsal wing surface while the female from Panama (Fig. 18) has the basal parts of both wings blue. Their genitalia, however, are uniform. This geographic variability is similar to that in Oenomaus cyanovenata.
Territorial behavior on a hilltop in Panama (Canal Area, Gamboa, Cerro Pelado) was observed in January and August at 15:00 hours (vouchers in USNM). Two mating pairs were also collected on the same hilltop in January and March at 15:00 hours (vouchers in USNM).
We illustrate adult females that were collected in copula (Figs 18–19) and the genitalia of one (Fig. 37).
A female from Brazil, Pará (CF-LYC-064), which has a wing pattern similar to the females collected in copula, is 3.1% divergent from a male from Peru (CF-LYC-085).
Taxonomy. A decade ago Oenomaus was a monotypic genus, but it now consists of 28 described species (albeit, it is still unclear if Oenomaus melleus and Oenomaus curiosa belong to Oenomaus or Porthecla). Further, if a phylogenetic analysis shows that Porthecla is paraphyletic in terms of Oenomaus, which is possible because Porthecla was distinguished by character states that may be plesiomorphic, then Oenomaus will be one of the most species-rich eumaeine genera with 40 species (
There are three biological reasons why the diversity of Oenomaus was not recognized until recently. First, about ¾ of the species have an indistinguishable, or barely distinguishable, ventral wing pattern that is similar to that of Oenomaus atena (e.g., Figs 12–19). Among species with this wing pattern, there is a great diversity of male genitalic forms that were first documented by
DNA barcoding. Thirty-eight Oenomaus males belonging to 19 species have been successfully “barcoded” (>200 bp) (extraction and sequencing methods given in
Male-female associations. Associating males and females in Oenomaus is sometimes very difficult. Only eight of the 28 recognized species previously had the sexes associated. In this paper we associate the sexes of another ten species based on mating pairs collected in copula and on similarity of ventral wing patterns, habitats, geographic distributions, and mitochondrial COI DNA sequences. The DNA “barcodes” have great potential (e.g.,
Biology. Oenomaus ortygnus is a well-known pest of cultivated soursop (also called guanábana, Annona muricata L., Annonaceae) and relatives (e.g.,
Most Oenomaus species inhabit relatively undisturbed lowland wet forest, but some species seem to be restricted to other habitats. Oenomaus andi is montane, being found so far only above 1450 m, while Oenomaus geba is known only from lower montane forest. Oenomaus morroensis occurs so far only in subtropical and lower montane forest. Oenomaus druceus has been found only in scrubby deciduous forest. A number of other Oenomaus species have broader habitat requirements. Some wet lowland species also occur in montane habitats, for which Oenomaus atesa and Oenomaus moseri are representative examples. Others, such as Oenomaus floreus, O. gaia, and Oenomaus isabellae, may inhabit dry deciduous forest. As previously noted, Oenomaus ortygnus is the only Oenomaus species that is regularly found in both undisturbed and disturbed habitats.
We are indebted to Evgeny V. Zakharov (data manager, University of Guelph, Biodiversity Institute of Ontario, Canada) for collaboration in sequencing the mitochondrial COI gene (BOLD project) and Dr D. H. Janzen for allowing us to use his barcode results for Oenomaus from Costa Rica. Accordingly, we thank Stéphane Brûlé (SEAG), Pierre and Jacques Jauffret for hairstreak legs; Jean-Yves Gallard, Serge Fernandez and Thibault Rosant for material from French Guiana.
For allowing us to examine specimens in collections under their care, we are grateful to Dr Jacques Pierre (MNHN), Blanca Huertas (NHM), Dr Gerardo Lamas (MUSM), Dr Olaf H.H. Mielke (DZUP), Dr David Grimaldi and Suzanne Rab Green(AMNH). For giving us access to their personal collections, we thank Pierre Boyer (France), Louis and Yvan Diringer (France), Christian Castelain (France), Alfred Moser (Brazil), Carlos Prieto (Colombia) and Jean-François Le Crom (Colombia).
For comments on the type locality of Oenomaus moseri, we are grateful to Olaf H.H. Mielke. For allowing us to use his data on the breeding of Oenomaus moseri, we thank Hipólito Ferreira Paulino Neto (Brazil). For checking Peruvian localities, we thank Gerardo Lamas. For reading and commenting upon the manuscript, we thank John Burns and Carlos Prieto. We also thank the anonymous reviewers for their helpful comments.
Finally, the senior author acknowledges a Smithsonian Institution short term visitor fellowship during summer 2011.