





(C) 2011 Thomas Bourguignon. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Recently, we completed a revision of the Termitidae from New Guinea and neighboring islands, recording a total of 45 species. Here, we revise a second family, the Rhinotermitidae, to progress towards a full picture of the termite diversity in New Guinea. Altogether, 6 genera and 15 species are recorded, among which two species, Coptotermes gambrinus and Parrhinotermes barbatus, are new to science. The genus Heterotermes is reported from New Guinea for the first time, with two species restricted to the southern part of the island. We also provide the first New Guinea records for six species of the genera Coptotermes and Schedorhinotermes. We briefly describe soldiers and imagoes of each species and provide a key based on soldier characters. Finally, we discuss the taxonomic and biogeographical implication of our results. A replacement name, Schedolimulus minutides Bourguignon, is proposed for the termitophilous staphylinid Schedolimulus minutus Bourguignon, to solve a question of secondary homonymy.
termites, Papua New Guinea, Indonesia, new species
The Rhinotermitidae constitute one of the most widespread termite families, including numerous pest species. First established under the invalid name Mesotermitidae (
The Rhinotermitidae are widely distributed across tropical, subtropical and temperate regions (
Some recent revisions or compilations of distributional data are available for Rhinotermitidae in Southeast Asia and Australia. Faunal lists mention 21 species from Sundaland (Peninsular Malaysia to Borneo:
Extensive termite collecting was carried out by J. M. Pasteels (in collection records: JMP), Y. Roisin (YR) and M. Leponce (ML) in New Guinea and some neighboring islands between 1978 and 1995 although records from Indonesian Papua are almost exclusively limited to the “bird’s neck” area, around Nabire and Kaimana (Figs 30, 82). Termite specimens were collected with tweezers and preserved in 80% alcohol or fixed in Bouin’s fluid or in a formol-alcohol-acetic acid (20:75:5) mixture. We also had the opportunity to examine samples collected in alcohol by Alfred E. and Eleanor Emerson in 1962–1963 (AE), as well as a few samples from other sources. Localities where specimens were collected are given as well as their approximate geographic coordinates (Appendix 1), obtained by cross-checking maps, GoogleTM Earth positioning and the National Geospatial Agency GEOnet Names Server (http://geonames.nga.mil/ggmagaz/).
Systematic charactersThis study is based on the morphology of soldier and alate castes. Soldiers supply most of the important taxonomic information at the species level. The following characters are of major interest: size, general shape of the head, shape of mandibles, shape of postmentum, pilosity of head and number of antennal articles. Alates, when available, may give taxonomic information at the species level by the general shape of their head and pronotum.
Measurements and their abbreviationsThe measurements used, detailed below, follow the guidelines of
Soldiers. (Fig. 1, 2, 4): HLF – Head length to fontanelle; HLC – Head length to apex of the clypeus; HLL – Head length to apex of the labrum; HW– Head maximum width; PL – Pronotum length; PW – Pronotum width; RML – Right mandible length; LML – Left mandible length; PML – Postmentum length; MPW –Maximum postmentum width; mPW – Minimum postmentum width; T3L – Hind tibia length.

Measurements. A-B, D soldier: (1), HLF, head length to fontanelle; (2), HLC, head length to apex of the clypeus; (3), HLL, head length to apex of the labrum; (4), HW, head maximum width; (5), PL, pronotum length; (6), PW, pronotum width; (7), RML, right mandible length; (9), PML, postmentum length; (10), MPW, maximum postmentum width; (11), mPW, minimum postmentum width. C imago: (14), HLC, head length to apex of the clypeus; (15), HWE, head width with eyes; (16), PL, pronotum length; (17), PW, pronotum width.
Imagoes. (Fig. 3): TBL – Total body length (without wings); HLC – Head length to apex of the clypeus; HWE – Head width with eyes; PL – Pronotum length; PW – Pronotum width; FWL – Forewing length (with scale); ED – Eyes maximum diameter
MicroscopyFor scanning electron microscopy, specimens were dehydrated in a conventional ethanol series, impregnated for 24 h in hexamethyldisilazane, air dried and gold coated. Digital images were taken with a Philips XL 30 ESEM.
Collections and their abbreviationsSpecies determinations were carried out after comparison with type series or identified specimens kept in the following museums
AMNH - American Museum of Natural History, New York, USA
BMNH - Natural History Museum, London, UK
CUMZ - Cambridge University Museum of Zoology, Cambridge, UK
IRSNB - Institut Royal des Sciences Naturelles, Brussels, Belgium
MVMA - Museum of Victoria, Abbotsford, Victoria, Australia
NHRS - Naturhistoriska Riksmuseet, Stockholm, Sweden
Samples numbered #PNGT*** (Papua New Guinea Termites) or #IRJT*** (Irian Jaya Termites) are in the authors’ laboratory collection at the Université Libre de Bruxelles. The major part of this collection will ultimately be deposited at the IRSNB.
Systematicshttp://species-id.net/wiki/Coptotermes
Termes gestroi Wasmann, 1896, by monotypy.
Imago head circular, covered by many setae. Fontanelle indistinct and appearing as a small spot in the middle of the head. Large ocelli located above the eyes. Pronotum and abdomen densely hairy. Soldiers with pyriform head capsule, slightly hairy. Fontanelle very large, directed forward, through which the latex-like secretion of the frontal gland is discharged. Labrum short, triangular-shaped. Mandibles narrow and elongated, curved at tip; right mandible without subsidiary teeth and serrations; left one with a basal tooth and serrations. Antennae with 12 to 16 articles.
The genus Coptotermes is broadly distributed, occurring in all tropical and subtropical regions. It comprises species adapted to all major biomes, from tropical rainforest to arid steppes and deserts. Nests are generally found in logs, in the heartwood of living trees, or underground (
http://species-id.net/wiki/Coptotermes_elisae
Figs 5–9, 30Syntypes: PAPUA NEW GUINEA: Morobe: Simbang, 12.ix.1898 (L. Biró), alates only (IRSNB). Other material. PAPUA NEW GUINEA: Madang: Laing Island, 28.xi.1978 (JMP), with queen and alate (#PNGT22); Laing Island, 11.vi.1987 (YR), with alates (#PNGT1101); Laing Island, 24.viii.1988 (YR), nest within fallen Erythrina tree (#PNGT1260); Manam Island, 23.vi.1981 (JMP), feeding on live tree (#PNGT101); Bogia–Josephstaal road, 10 km S Guam bridge, 26.vi.1984 (YR) (#PNGT711); Hansa Point, 22.viii.1984 (YR) (#PNGT785): Awar, 16.ix.1984 (YR) (#PNGT831); Tabele (Manam Island), 19.ix.1984 (YR), in decaying palm (#PNGT839); Sepen No.1, 01.iii.1988 (YR) (#PNGT1164); Baitabag, v.1999 and 17.x.1999 (L. Čižek) (#7, #15, J. Šobotník’s collection). Morobe: Sirasira, 14.v.1988 (YR) (#PNGT1213); Bulolo, 22.v.1987 (YR), in dead hoop pine (Araucaria cunninghammi) trunk on the ground (#PNGT1080); Central: Brown River forest, 15.xi.1962 (AE), from dead branch on forest floor (AMNH); Brown River, 21.xi.1962 (AE), from standing dead stump in forest (AMNH); 24 km NE Port Moresby, 23.xi.1962 (AE), covered galleries over large surface of live tree, with hole to heart wood (AMNH); Subitana plantation, 24.xi.1962 (AE), from standing dead rubber tree in mature grove (AMNH); Sogeri, 15.vii.1984 (YR), in dead liana along tree trunk in small patch of forest (#PNGT740); Sogeri, 23.iii.1985 (JMP & YR) (#PNGT1005); Oro: Kokoda, 13.iii.1985 (JMP & YR) (#PNGT952); Kokoda, 17.iii.1985 (JMP & YR) (#PNGT981); Koiasi, 14.iii.1985 (JMP & YR) (#PNGT959); New Ireland: Konos, 29.v.1984 (JMP & YR) (#PNGT624); Sandaun: Yapsiei, 10.iii.1994 (YR & ML) (#PNGT1733 & 1734); Fly: Morehead, 25.iii.1989 (YR & ML) (#PNGT1447); Wipim, 01.iv.1989 (YR & ML) (#PNGT1509); Tabubil, 20.v.1990 (YR & ML) (#PNGT1541); Lake Murray, 24.v.1990 (YR & ML) (#PNGT1582); Lake Murray, 25.v.1990 (YR & ML), with queen (#PNGT1589); Nomad, 29.v.1990 (YR & ML) (#PNGT1609); Nomad, 31.v.1990 (YR & ML) (#PNGT1624); Nomad, 01.vi.1990 (YR & ML) (#PNGT1637); Southern Highlands: Pimaga, alt. 950m, 18.x.1988 (YR) (#PNGT1316); INDONESIA: Papua: Road Nabire-Mapia km 43, 26.xi.1995 (YR) (#IRJT171); Road Nabire-Mapia km 62, 18.xi.1995 (YR) (#IRJT72, 73); Coa, 22.xi.1995 (YR) (#IRJT123); Kaimana, 23.xi.1995 (YR) (#IRJT140).
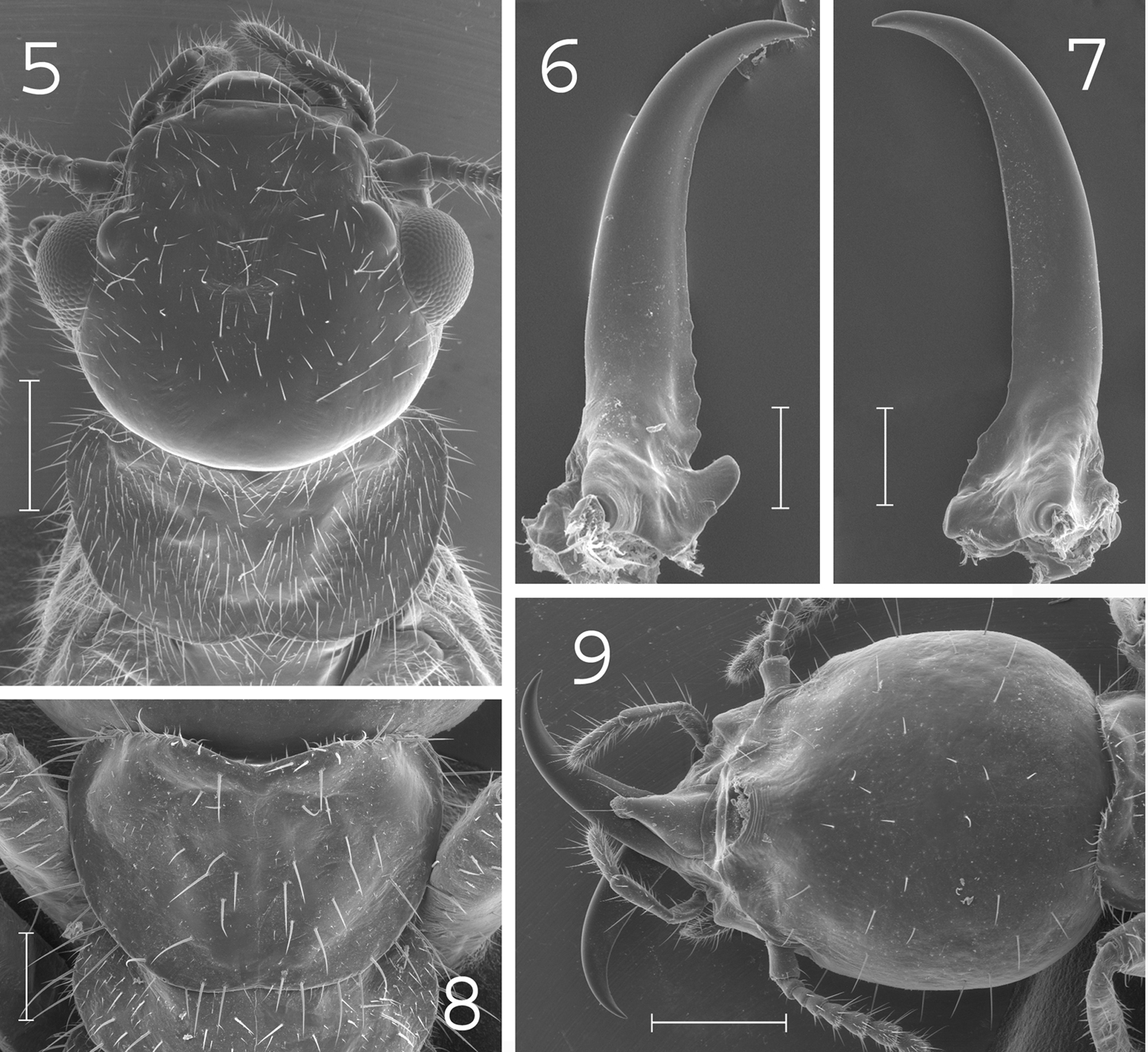
Coptotermes elisae. Imago: 5 head and pronotum. Soldier: 6 left mandible; 7 right mandible; 8 pronotum; 9 head. Scale bars: 5, 9: 0.5mm; 6, 7, 8: 0.2mm.
(Fig. 5).Head rounded and densely hairy. Large eyes. Pronotum wide and elongated, covered by numerous setae of medium and large size. Antennae with 20 to 22 articles. Measurements (mm) of 8 imagoes from the type colony and 4 imagoes from one other colony (between parentheses): TBL: 8.27–8.99 (7.81–8.12); HLC: 1.53–1.87 (1.48–1.62); HWE: 1.84–1.96 (1.79–1.81); PL: 1.04–1.12 (1.02–1.07) ; PW: 1.63–1.70 (1.59–1.61); FWL: n.a. (13–13.2); ED: 0.45–0.50 (0.45–0.5).
(Figs 6–9).Soldiers of large size. Head rounded, covered by about 40 setae. Fontanelle large, with opening well visible from above. Antennae generally with 16 articles. Pronotum elongated, covered by about 70 setae mainly located on the edges. Mesonotum, metanotum and abdomen bearing plenty of setae. Mandibles with tips extremely curved. Measurements (mm) of 99 soldiers from 33 colonies: HLF: 1.11–1.47; HLL: 1.70–2.20; HW: 1.21–1.54; PL: 0.38–0.64; PW: 0.77–1.03; RML: 0.97–1.20; MPW: 0.30–0.49; T3L: 1.05–1.34.
Coptotermes elisae can be distinguished from other New Guinean species by its large rounded head with fontanelle opening well visible from above, its strongly curved mandibles and elongated pronotum in the soldier caste.
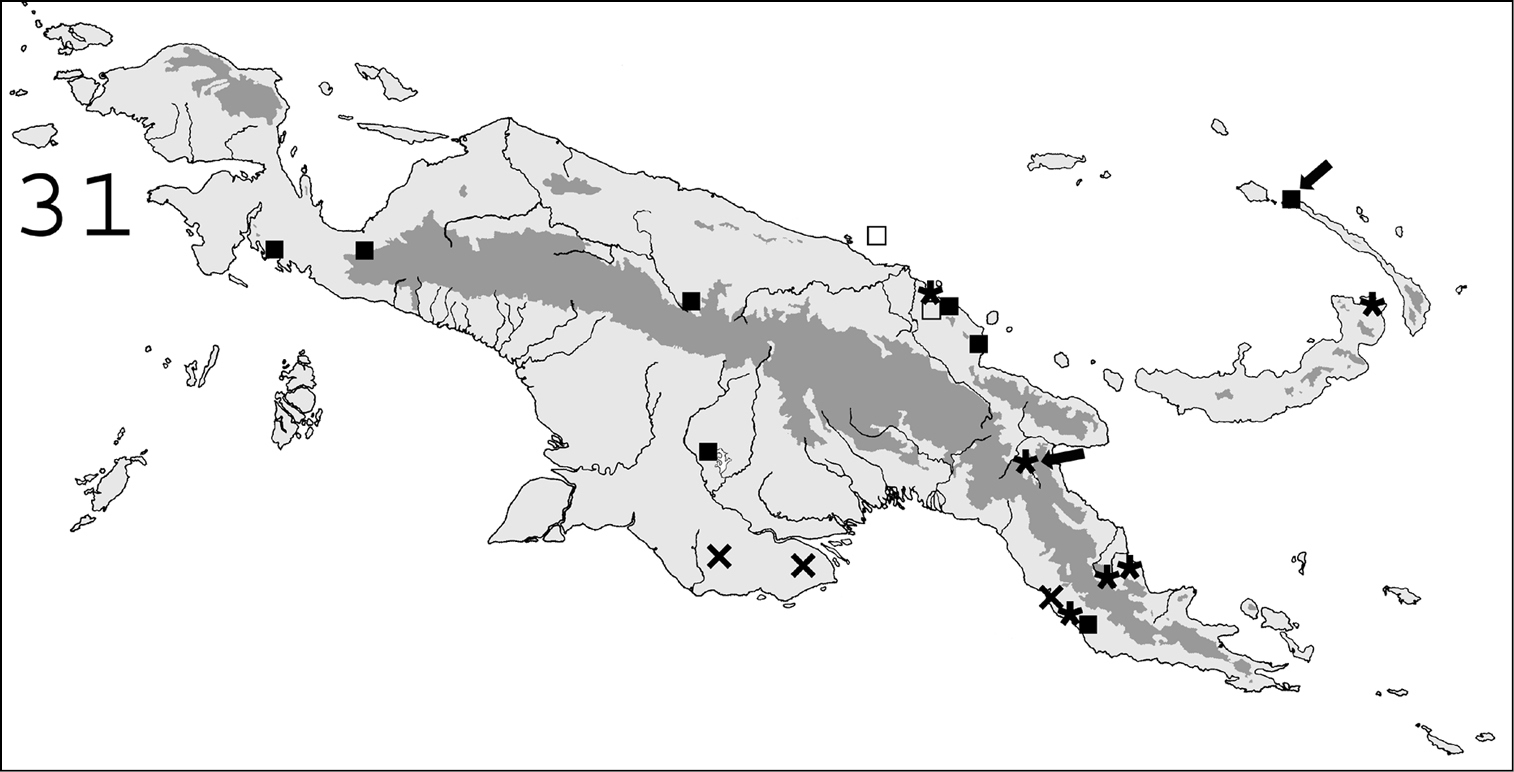
(Fig. 30).This species is widespread in New Guinean forests. It also occurs in New Ireland. It has also been reported from the following localities, but the relevant material was not examined: Bukaua (as Coptotermes hyaloapex:
Coptophysa obesa (Coleoptera: Staphylinidae) was found in colony #PNGT740 of this species in Sogeri (
http://species-id.net/wiki/Coptotermes_remotus
Figs 10–15, 31Syntypes: PAPUA NEW GUINEA: New Ireland: Kavieng (as Kaewieng), 4.x.1923 (H.G. Wallace) (NMVA T-18703, T-18704). Other material: PAPUA NEW GUINEA: Madang: Hansa Point, 08.vii.1984 (YR) (#PNGT719); Awar, 11.ix.1984 (YR) (#PNGT828); Nubia, 17.ii.1988 (YR) (#PNGT1154); Potsdam plantation, 22.xi.1988 (YR) (#PNGT1333); Hatzfeldhafen, 25.viii.1984 (YR) (#PNGT795); Baitabag, 17.x.1999 (L. Čižek) (#1, J. Šobotník’s collection);Sandaun: Yapsiei, 10–11.iii.1994 (YR & ML) (#PNGT1735, 1744). Central: Sirinumu Dam, 7.iii.1985 (JMP & YR) (#PNGT923); Fly: Lake Murray, 23.v.1990 (YR & ML) (#PNGT1569). INDONESIA: Papua: Pusppenssat-IrJa, 13.xi.1995 (YR) (#IRJT6, 10, 12); Pusppenssat-IrJa, 19.xi.1995 (YR) (#IRJT84, 85); Pusppenssat-IrJa, 30.xi.1995 (YR) (#IRJT207); Road Nabire-Mapia km 43, 15.xi.1995 (YR) (#IRJT40, 41); Road Nabire-Mapia km 43, 26.xi.1995 (YR) (#IRJT169, 170); Road Nabire-Mapia km 62, 18.xi.1995 (YR) (#IRJT74, 75); Topo, 28.xi.1995 (YR) (#IRJT189); Sanoba, 29.xi.1995 (YR) (#IRJT195); Coa, 22.xi.1995 (YR) (#IRJT131): Kaimana, 23.xi.1995 (YR) (#IRJT141). Samples included with doubt: PAPUA NEW GUINEA: East Sepik: Koil Island, 17.vi.1981 (JMP) (#PNGT75); Madang: Hansa Point, 23.ix.1988 (YR) (#PNGT1281).
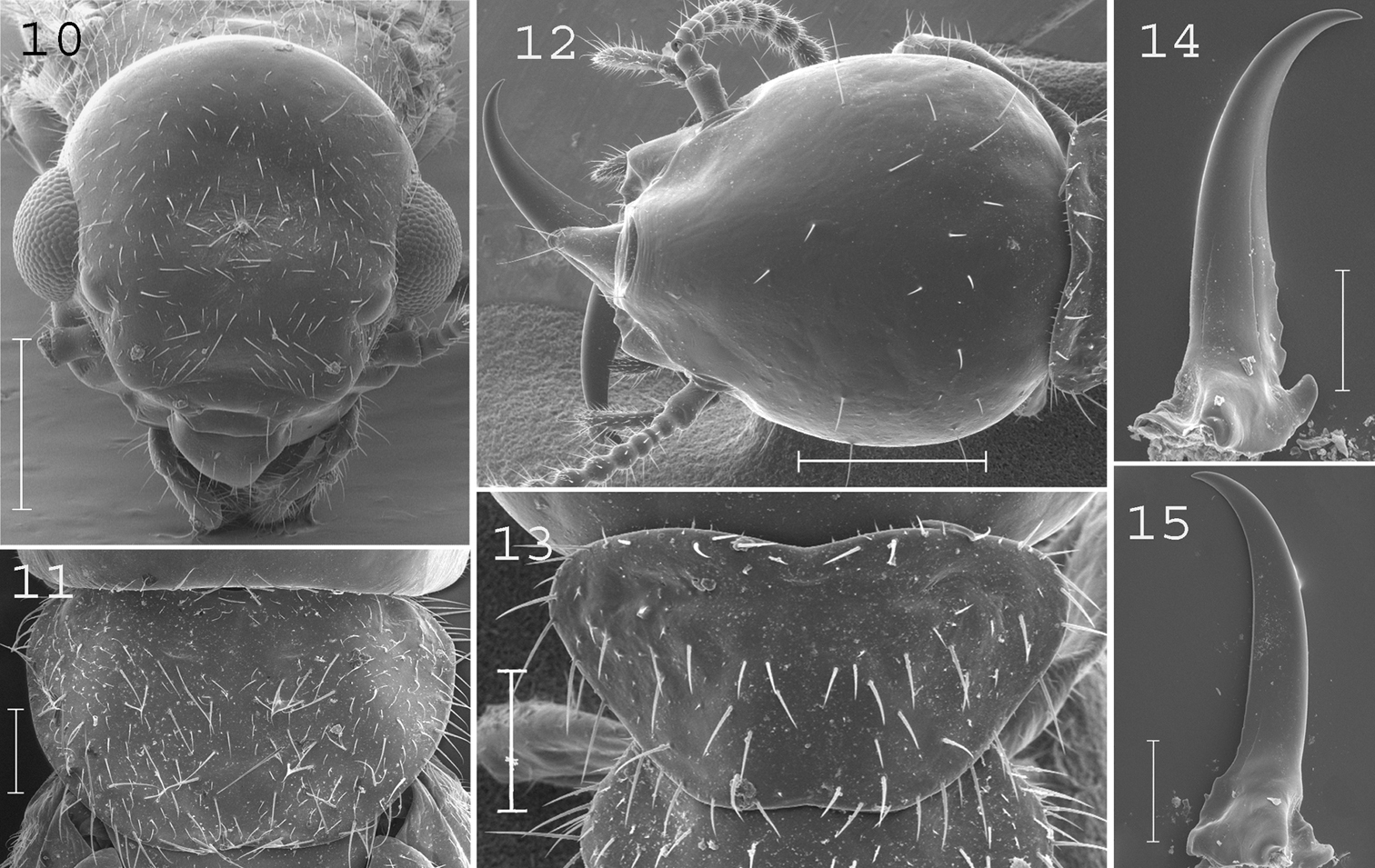
Coptotermes remotus. Imago: 10 head; 11 pronotum. Soldier: 12 head; 13 pronotum; 14 left mandible; 15 right mandible. Scale bars: 10, 12: 0.5mm; 11, 13, 14, 15: 0.2mm.
(Figs 10–11).Head densely hairy. Pronotum with many medium and large setae. Antennae with 17 articles. Measurements (mm) of 6 imagoes from 1 colony: TBL: 5.75–6.39; HLC: 1.09–1.18; HWE: 1.30–1.38; PL: 0.57–0.73; PW: 0.97–1.08; FWL: 9.92–11.09; ED: 0.33–0.44.
(Figs 12–15). Soldiers of small size. Head slightly longer than broad, covered by about 20 setae. Fontanelle with opening directed forward, not visible from above. Antennae generally with 14 articles, but occasionally with only 13 articles. Pronotum larger anteriorly than posteriorly, trapezoid-shaped, covered by about 50 setae. Mandibles with tips extremely curved. Right mandible with four serrations. Measurements (mm) of 5 soldiers from the type colony and 72 soldiers from 24 colonies (parentheses): HLF: 1.11–1.19 (1.01–1.29); HLL: 1.45–1.56 (1.23–1.63); HW: 1.00–1.04 (0.83–1.11); PL: 0.36–0.39 (0.29–0.46); PW: 0.67–0.70 (0.59–0.78); RML: 0.78–0.83 (0.69–0.85); MPW: 0.28–0.31 (0.26–0.38); T3L: 0.83–0.89 (0.68–0.99).
This species shows variation in size along its distribution range although no consistent characters allowed us to split it up. Coptotermes remotus most resembles the Australian species Coptotermes lacteus and the Malayan ones Coptotermes bentongensis and Coptotermes sepangensis. These last two species occur in sympatry and are morphologically undistinguishable, making them probable synonyms. Soldiers of Coptotermes remotus can be distinguished from other New Guinean species by their small size and mandibles curved at the tip.
This species, originally described from Kavieng, New Ireland, is widespread throughout New Guinean forests. It is abundant in the bird’s neck area of Indonesian Papua, but only a few specimens were collected from Papua New Guinea (Fig. 31).
Coptophysella pulposa (Coleoptera: Staphylinidae) was found in colony #PNGT795 of this species in Hatzfeldthafen (
http://species-id.net/wiki/Coptotermes_grandiceps
Figs 16–21, 30Topotype: SOLOMON ISLANDS: Tulaghi (as Tulagi), iii.1933 (R.A. Lever) (AMNH). Holotype of Coptotermes obiratus:PAPUA NEW GUINEA: Central: Waima, vii.1922 (G.F. Hill) (NMVA).
PAPUA NEW GUINEA: Oro: Mambare River, 27.iv.1922 (G.F. Hill) (NMVA). Fly: Morehead, 25.iii.1989 (YR & ML) (#PNGT1447); Wipim, 30.iii.1989 (YR & ML) (#PNGT1488); Lake Murray, 22.v.1990 (YR & ML) (#PNGT1561); Lake Murray, 25.v.1990 (YR & ML) (#PNGT1594); Nomad, 31.v.1990 (YR & ML) (#PNGT1627); Nomad, 02.vi.1990 (YR & ML) (#PNGT1651); Central: Subitana plantation, 24.xi.1962 (AE), in stump of rubber tree (AMNH); Sirinumu Dam, 06.iii.1985 (JMP & YR) (#PNGT916); Sirinumu Dam, 08.iii.1985 (JMP & YR) (#PNGT929); Varirata National Park, 06.xii.1988 (YR & Phille P. Daur) (#PNGT1346); National Capital: 2 km E Port Moresby, 18.xi.1962 (AE), from log in dry eucalypt savanna (AMNH); 8 km E Port Moresby, 16.xi.1962 (AE), under log in eucalypt savanna (AMNH); 8 km E Port Moresby, 18.xi.1962 (AE), from dead log in ravine woods in savanna (AMNH); 10 km NW Port Moresby, 20.xi.1962 (AE), from log on ground in eucalypt savanna (AMNH); 19 km NW Port Moresby, 22.xi.1962 (AE), from fallen log in savanna (AMNH); INDONESIA: Papua: Pusppenssat-IrJa, 13.xi.1995 (YR) (#IRJT17).

Coptotermes grandiceps. Imago: 16 head; 17 pronotum. Soldier: 18 head; 19 pronotum; 20 left mandible; 21 right mandible. Scale bars: 16, 17, 18: 0.5mm; 19, 20, 21: 0.2mm.
In his revision of termites from Australia,
(Figs 16–17).Head moderately hairy. Pronotum covered by many setae of medium size. Antennae with 20 articles. Measurements (mm) of 6 imagoes from one colony: TBL: 6.08–7.55; HLC: 1.19–1.27; HWE: 1.33–1.41; PL: 0.73–0.82; PW: 1.23–1.29; FWL: 9.97–10.61; ED: 0.31–0.42.
(Figs 18–21). Soldiers of large size. Head elongated, larger posteriorly than anteriorly, covered by about 30 setae. Fontanelle narrow, with opening directed frontally, not visible from above. Antennae generally with 15 articles, sometimes with 14 articles. Pronotum wide, moderately long, covered by about 60 setae. Mandibles elongated, curved at tip. Measurements (mm) of holotype of Coptotermes obiratus, 1 soldier from the type colony of Coptotermes grandiceps [brackets], and27 soldiers from 9 colonies (parentheses): HLF: 1.53 [1.52] (1.21–1.44); HLL: 2.17 [2.19] (1.89–2.20); HW: 1.38 [1.31] (1.17–1.33); PL: 0.51 [0.50] (0.40–0.56); PW: 0.97 [0.95] (0.79–1.00); RML: 1.17 [1.24] (1.03–1.15); MPW: 0.48 [0.42] (0.31–0.46); T3L: 1.22 [n.a.] (1.07–1.21).
This species is closely allied to Coptotermes pamuae from which it can be distinguished by its larger soldiers with more curved mandibles.
(Fig. 30). Coptotermes grandiceps, originally described from the Solomon Islands, occurs in southern New Guinea and the Papuan peninsula.
http://species-id.net/wiki/Coptotermes_pamuae
Figs 22–25, 31Paratype soldier from type colony: SOLOMON ISLANDS: San Cristobal: Pamua, viii.1916 (W.M. Mann) (AMNH). PAPUA NEW GUINEA: National Capital: UPNG campus, 04.xii.1988 (YR) (#PNGT1338); Central: Varirata National Park, 06.xii.1988 (YR & Phille P. Daur) (#PNGT1348); Fly: Morehead, 27.iii.1989 (YR & ML) (#PNGT1466); Wipim, 30.iii.1989 (YR & ML) (#PNGT1486).
Unknown.
(Figs 22–25). Soldiers of medium size. Head elongated, egg-shaped, slightly larger posteriorly than anteriorly, covered by about 30 setae. Fontanelle narrow, with opening directed forward. Antennae generally with 13 articles, sometimes with 14 articles. Pronotum short and narrow, covered by about 40 setae. Mandibles elongated, slightly curved at tip. Measurements (mm) of 1 paratype and 12 soldiers from 4 colonies (parentheses): HLF: 1.28 (1.11–1.27); HLL: 1.70 (1.58–1.79); HW: 1.12 (1.00–1.10); PL: 0.41 (0.31–0.44); PW: 0.79 (0.63–0.74); RML: 0.77 (0.89–0.98); MPW: 0.37 (0.28–0.32); T3L: 0.82 (0.92–1.08).
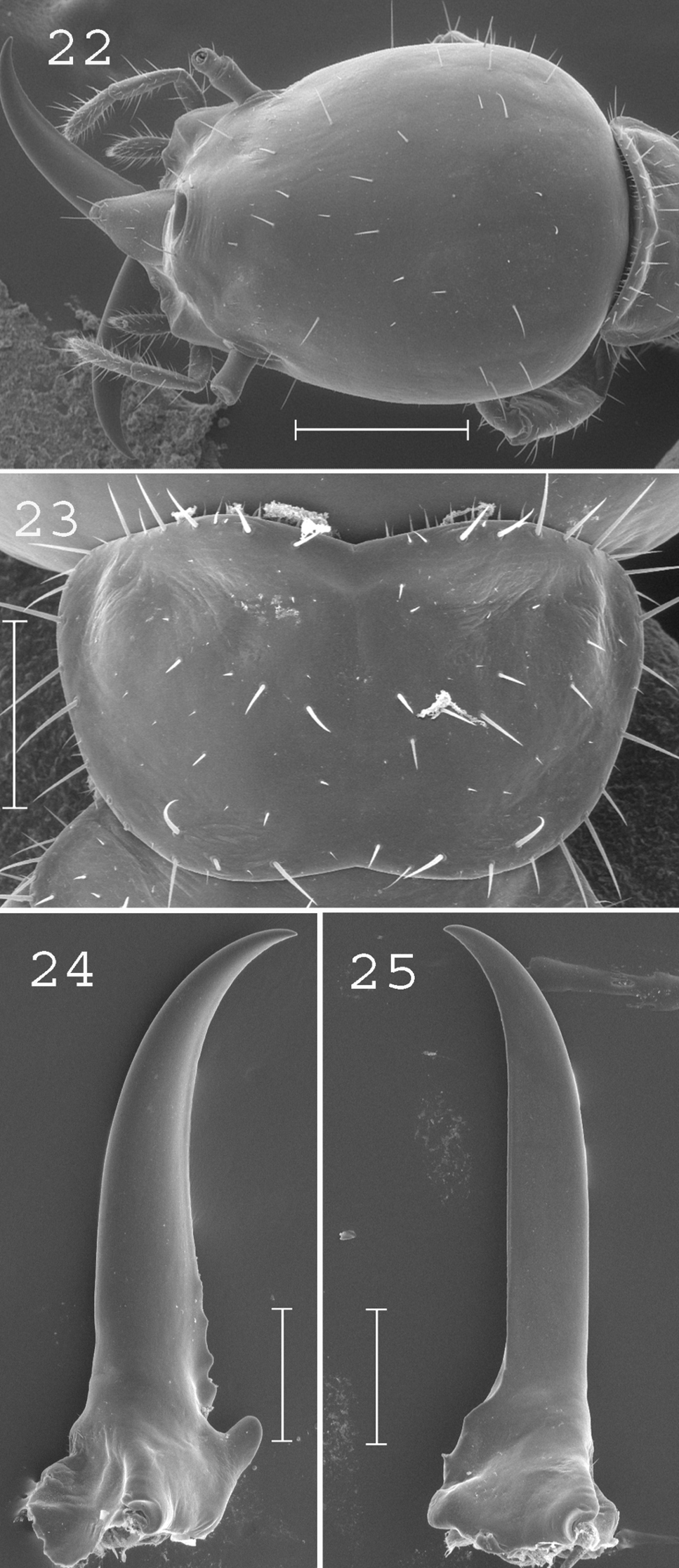
Coptotermes pamuae. Soldier: 22 head; 23 pronotum; 24 left mandible; 25 right mandible. Scale bars: 2:2 0.5mm; 23, 24, 25: 0.2mm.
This species is closely related and imperfectly separated from Coptotermes grandiceps. However, the size and morphological differences between the two species are high enough to retain them as distinct taxa, even though some individuals cannot be unambiguously assigned. The same was already observed in the related Australian species Coptotermes acinaciformis Froggatt, which is believed to form a species complex (
(Fig. 31). This species, originally described from the Solomon Islands, was only collected in southern Papua New Guinea (Fly savannas and the Port Moresby region).
urn:lsid:zoobank.org:act:8E27AAD1-29A7-471B-BD03-58F21AC53514
http://species-id.net/wiki/Coptotermes_gambrinus
Figs 26–29, 31Soldier: PAPUA NEW GUINEA: Morobe: Bulolo, 22.v.1987 (YR), from stump of klinkii pine (Araucaria hunsteinii) (#PNGT1077). Paratypes: PAPUA NEW GUINEA: Morobe: Soldiers and workers from same colony as holotype, same data (#PNGT1077); Mount Susu, 19.v.1988 (YR), in dead branch of klinkii pine on the ground (#PNGT1232); McAdam National Park, 20.v.1988 (YR) (#PNGT1238). Madang: Hatzfeldhafen, 23.ix.1984 (YR) (#PNGT847); Oro: Kokoda, 13.iii.1985 (JMP & YR) (#PNGT950); Popondetta, 18.iii.1985 (JMP & YR) (#PNGT986). Central: Sirinumu dam, 07.iii.1985 (JMP & YR) (#PNGT918); Brown River, 21.iii.1985 (JMP & YR) (#PNGT996); East New Britain: Warengoi, 19.v.1984 (JMP & YR) (#PNGT562, 567).
unknown.
(Figs 26–29). Soldiers of small size. Head twice broader posteriorly than anteriorly (at the level of mandibles), covered by about 10 setae. Antennae generally with 15 articles. Pronotum covered by about 15 setae, slightly larger anteriorly than posteriorly. Mandibles very short, with apex almost not curved. Measurements (mm) of 30 soldiers from 10 colonies: HLF: 0.92–1.07; HLL: 1.22–1.48; HW: 0.82–0.98; PL: 0.27–0.41; PW: 0.50–0.70; RML: 0.56–0.74; MPW: 0.23–0.36; T3L: 0.75–0.88.

Coptotermes gambrinus. Soldier: 26 head; 27 pronotum; 28 left mandible; 29 right mandible. Scale bars: 26: 0.5mm 27, 28, 29: 0.2mm.

Known distribution in New Guinea of: ✚ Coptotermes elisae; ∗ Coptotermes grandiceps. Arrow points to type locality.
Known distribution in New Guinea of: ∗ Coptotermes gambrinus; ✖ Coptotermes pamuae; ■ Coptotermes remotus; □ doubtful samples of Coptotermes remotus. Arrows point to type localities.
This species is allied to Coptotermes remotus from which it is easily recognisable by its shorter and less curved soldier mandible tips.
(Fig. 31). This species occurs in Eastern New Guinea and in New Britain.
We named this species in honor of Gambrinus, a legendary character from Flanders, famous for enjoying life.
http://species-id.net/wiki/Genus_Heterotermes
Heterotermes platycephalus Froggatt, 1897, by monotypy.
Imago head roughly oval, narrower anteriorly than posteriorly. Fontanelle small, located in the middle of the head. Eyes small and flat. Ocelli situated in front of the head, before eyes. Antennae generally with 15 to 19 articles. Pronotum elongated, narrower than head. Soldier head long and narrow, rectangular-shaped. Fontanelle small, circular, situated forward. Labrum short to medium-sized, about half as long as mandibles. Mandibles sabre-shaped, slightly curved at tips. Left mandible with a tooth and some serrations at the base. Right mandible without basal tooth and serrations. Antennae with 13 to 18 articles.
Most species of Heterotermes are tropical (
http://species-id.net/wiki/Heterotermes_vagus
Figs 32–36, 44Lectotype and paralectotype soldier: AUSTRALIA: Northern Territory: Darwin, 01.viii.1914 (G.F. Hill) (NMVA #T-10848, #T-18705) Lectotype of Heterotermes venustus: AUSTRALIA: Northern Territory: Stapleton, 4.xi.1914 (G.F. Hill) (NMVA #T-10850). Other material: PAPUA NEW GUINEA: Fly: Morehead, 23.iii.1989 (YR & ML) (#PNGT1419, 1420, 1422); Morehead, 25.iii.1989 (YR & ML) (#PNGT1441); Lake Murray, 25.v.1990 (YR & ML) (#PNGT1588, 1597).
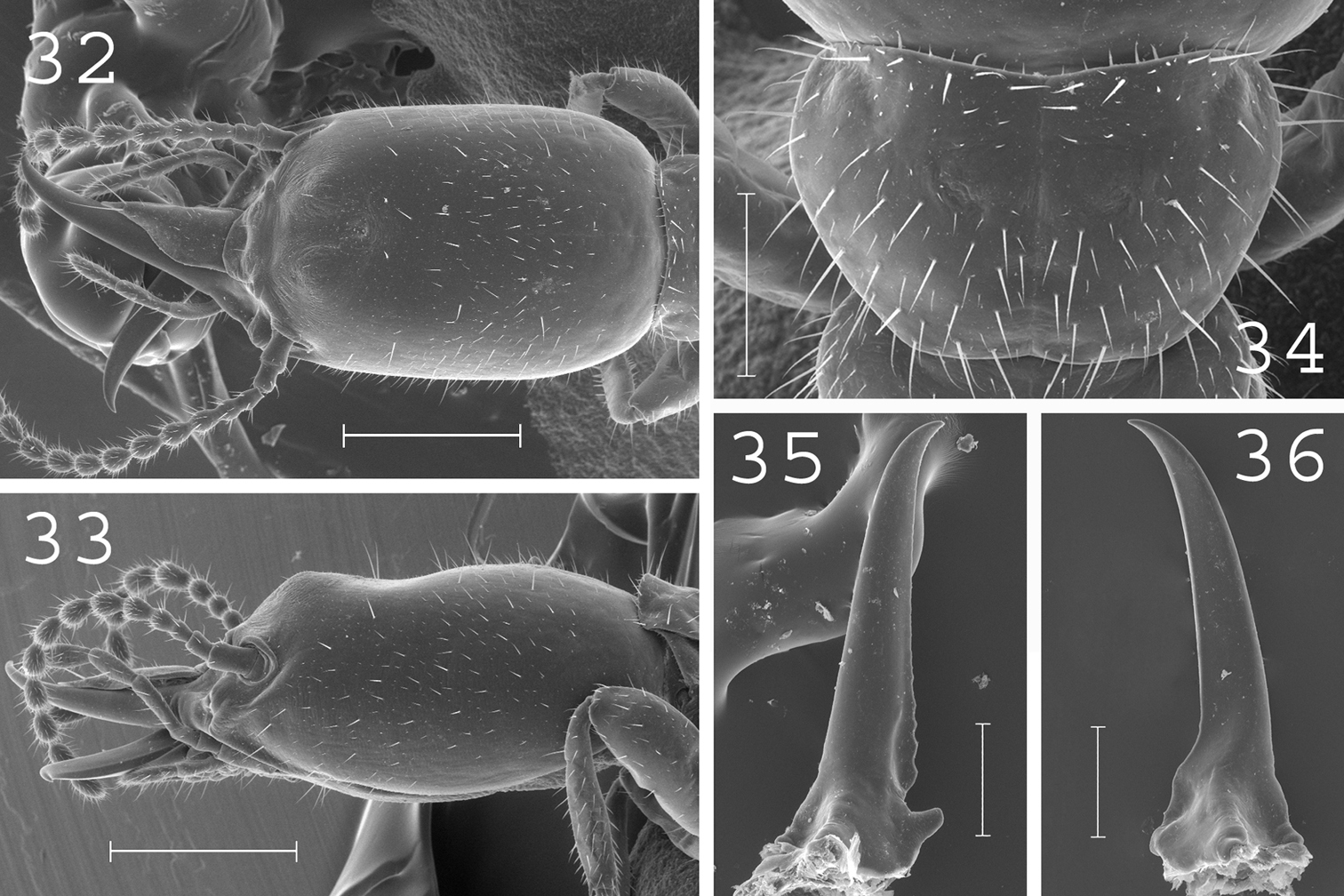
Heterotermes vagus. Soldier: 32 head in dorsal view; 33 head in lateral view; 34 pronotum; 35 left mandible; 36 right mandible. Scale bars: 32, 33: 0.5mm; 34, 35, 36: 0.2mm.
Unknown from New Guinea (see
(Figs 32–36). Soldiers of small size. Head elongated, with a prominent hump at front, covered by plenty of short setae. Labrum elongated with sharp tip. Antennae generally with 13 articles. Pronotum short and narrow, covered by about 50 setae. Postmentum slightly narrow, without setae. Mandibles slightly curved at tips. Measurements (mm) of lectotype of Heterotermes vagus, lectotype of Heterotermes venustus [brackets] and 18 soldiers from 6 colonies (parentheses): HLC: 1.32 [1.43] (1.12–1.37); HLL: 1.75 [1.83] (1.48–1.76); HW: 0.80 [0.82] (0.74–0.83); PL: 0.40 [0.41] (0.32–0.50); PW: 0.55 [0.62] (0.49–0.60); RML: 0.92 [0.89] (0.80–0.93); MPW: 0.34 [0.31] (0.29–0.33); T3L: 0.65 [0.66] (0.54–0.66).
(Fig. 44).This species was collected in Sourthern Papua New Guinea. It is also known from northernmost Queensland and Northern Territory (Australia) (
http://species-id.net/wiki/Heterotermes_paradoxus
Figs 37 -44AUSTRALIA: Northern Territory: 37 km SE Newcastle Waters, 16.vi.1936 (coll. det. G.F. Hill) (ANIC #10–2186); Queensland: Weipa mission, 05.x.1957 (F.J. Gay & J.H. Calaby) (ANIC #10–8659); PAPUA NEW GUINEA: Central: Sogeri, 14.vii.1984 (YR) (#PNGT731); Sogeri, 4.ii.1985 (YR) (#PNGT855); Sirinumu Dam, 6.iii.1985 (JMP & YR) (#PNGT912); Fly: Morehead, 24.iii.1989 (YR & ML) (#PNGT1431); Wipim, 29.iii.1989 (YR & ML) (#PNGT1471); Wipim, 2.iv.1989 (YR & ML) (#PNGT1517); Lake Murray, 22.v.1990 (YR & ML) (#PNGT1562); Lake Murray, 24.v.1990 (YR & ML) (#PNGT1577); Lake Murray, 25.v.1990 (YR & ML) (#PNGT1590a, ); Lake Murray, 27.v.1990 (YR & ML) (#PNGT1598, 1606). INDONESIA: Papua: Kaimana, 21.xi.1995 (YR) (#IRJT104, 111, 112); Kaimana, 23.xi.1995 (YR) (#IRJT142), two samples with alates (#IRJT143, 144); Pusppenssat-IrJa, 26.xi.1995 (YR) (#IRJT166).

Heterotermes paradoxus. Soldier: 37 head in dorsal view; 38 head in lateral view; 39 left mandible; 40 right mandible; 43 pronotum. Imago: 41 head; 42 pronotum. Scale bars: 37, 38: 1mm; 41: 0.5mm; 39, 40, 42, 43: 0.2mm.

Known distribution in New Guinea of: ✖ Heterotermes vagus; ● Heterotermes paradoxus.
(Figs 41–42).Head slightly elongated covered by several hundreds of setae. Pronotum elongated and moderately wide, covered by several hundreds of setae. Antennae generally with 18 articles, rarely with 17 articles. Eyes flat. Small ocelli in front of the eyes. Measurements (mm) of 12 imagoes from 1 colony: TBL: 5.16–6.31; HLC: 1.13–1.29; HWE: 0.98–1.14; PL: 0.59–0.64; PW: 0.76–0.85; FWL: 8.79–9.92; ED: 0.25–0.33.
(Figs 37–40, 43) Soldiers of large size. Head elongated, slightly rounded in the corners, with a well developed hump at front. Labrum with rounded apex. Antennae with 16 or 17 articles. Pronotum elongated, covered by numerous setae. Mandibles elongated, sligthly curved at tip. Measurements (mm) of 55 soldiers from 19 colonies: HLC: 1.34–1.98; HLL: 1.63–2.40; HW: 0.86–1.16; PL: 0.39–0.60; PW: 0.62–0.87; RML: 1.03–1.34; MPW: 0.35–0.48; T3L: 0.70–0.93.
Morphological variation occurs along the distribution range of Heterotermes paradoxus, though it appears insufficient to recognize distinct species. Heterotermes paradoxus differs from Heterotermes vagus by the larger size of its soldiers, its less elongated labrum with rounded end and its antennae with 16 or 17 articles.
(Fig. 44).This species is widespread in southern Papua New Guinea, and was found on both coasts of the “bird’s neck” in Indonesian Papua. It is also known from northern Australia, especially Queensland (Watson & Abbey 1993). It occurs in savanna as well as in forest.
http://species-id.net/wiki/Parrhinotermes
Termes aequalis Haviland, 1898, by monotypy (as Termes aqualis, incorrect spelling).
Imago head approximately circular, with fontanelle situated between eyes. Frons with a slightly visible groove. Labrum short, inclined downward, without groove. Antennae with 16 or 17 articles. Pronotum relatively short, generally of the same width or slightly narrower than head. Soldier head rectangular to ovoid. Frons and clypeus with a groove, from the narrow fontanelle to the beginning of the labrum. Labrum elongated, crossed by a groove in the middle, and garnished with an apical brush. Base of mandibles serrated. Left mandible with two subsidiary teeth. Right mandible with one subsidiary tooth. Antennae with 13 articles.
This genus is known from the Oriental region, northeastern India, the Papuan region and northern Australia (in northern Queensland) (
http://species-id.net/wiki/Parrhinotermes_browni
Figs 45–49, 55Paratypes, soldiers and workers: SOLOMON ISLANDS: Guadalcanal: Gold Ridge, 22.iii.1955 (E.S. Brown) (Brit. Mus. 1957–137, BMNH). Other material: PAPUA NEW GUINEA: East Sepik: Tsenap, 18.v.1929 (K.P. Schmidt), from log on ground (AMNH); Madang: Yagaum Hospital, 07.iv.1983 (YR) (#PNGT303, 306); Yagaum Hospital, 13.v.1983 (YR) (#PNGT340, 341); Bunapae, 18.ii.1984 (YR) (#PNGT546); Bunapae, 12.vi.1984 (YR) (#PNGT681); Guam bridge, 09.viii.1984 (YR), with royal pair (#PNGT782); Guam bridge, 12.ii.1985 (JMP & YR), with royal pair (#PNGT867); Guam bridge, 9.vi.1986 (YR), large colony with royal pair in rotten wood (#PNGT1023), and small sample (#PNGT1024); Sepen No.1, 29.vi.1986 (YR) (#PNGT1031); Hatzfeldthafen, 22.viii.1987 (YR), with primary king and ergatoid queen (#PNGT1130); Braham mission, 06.v.1988 (YR) (#PNGT1204); Wanuma, 05.viii.1969 (R. Zweifel) (AMNH); Morobe: Busu River, 17.xii.1962 (AE) (AMNH); 18 mi W Lae, 28.xi.1962 (AE), nest with royal pair in large log on forest floor (AMNH); Bulolo, 22.v.1987 (YR), in hoop pine (Araucaria cunninghamii) plantation (#PNGT1081); McAdam National Park, 20.v.1988 (YR) (#PNGT1236); Sandaun: Yapsiei, 11.iii.1994 (YR & ML) (#PNGT1742); Yapsiei, 12.iii.1994 (YR & ML), in standing dead wood, with alates (#PNGT1751); Central: Subitana plantation, xi.1962 (AE) (AMNH). INDONESIA: Papua: Pusppenssat-IrJa, 13.xi.1995 (YR) (#IRJT7, 9); Pusppenssat-IrJa, 14.xi.1995 (YR) (#IRJT28); Pusppenssat-IrJa, 15.xi.1995 (YR) (#IRJT39); Pusppenssat-IrJa, 29.xi.1995 (YR) (#IRJT203); road Nabire-Mapia km 43, 26.xi.1995 (YR) (#IRJT167).

Parrhinotermes browni. Imago: 45 head. Soldier: 46 head in dorsal view; 47 head in ventral view; 48 left mandible; 49 right mandible. Scale bars: 45, 46, 47: 0.5mm 48, 49: 0.2mm.
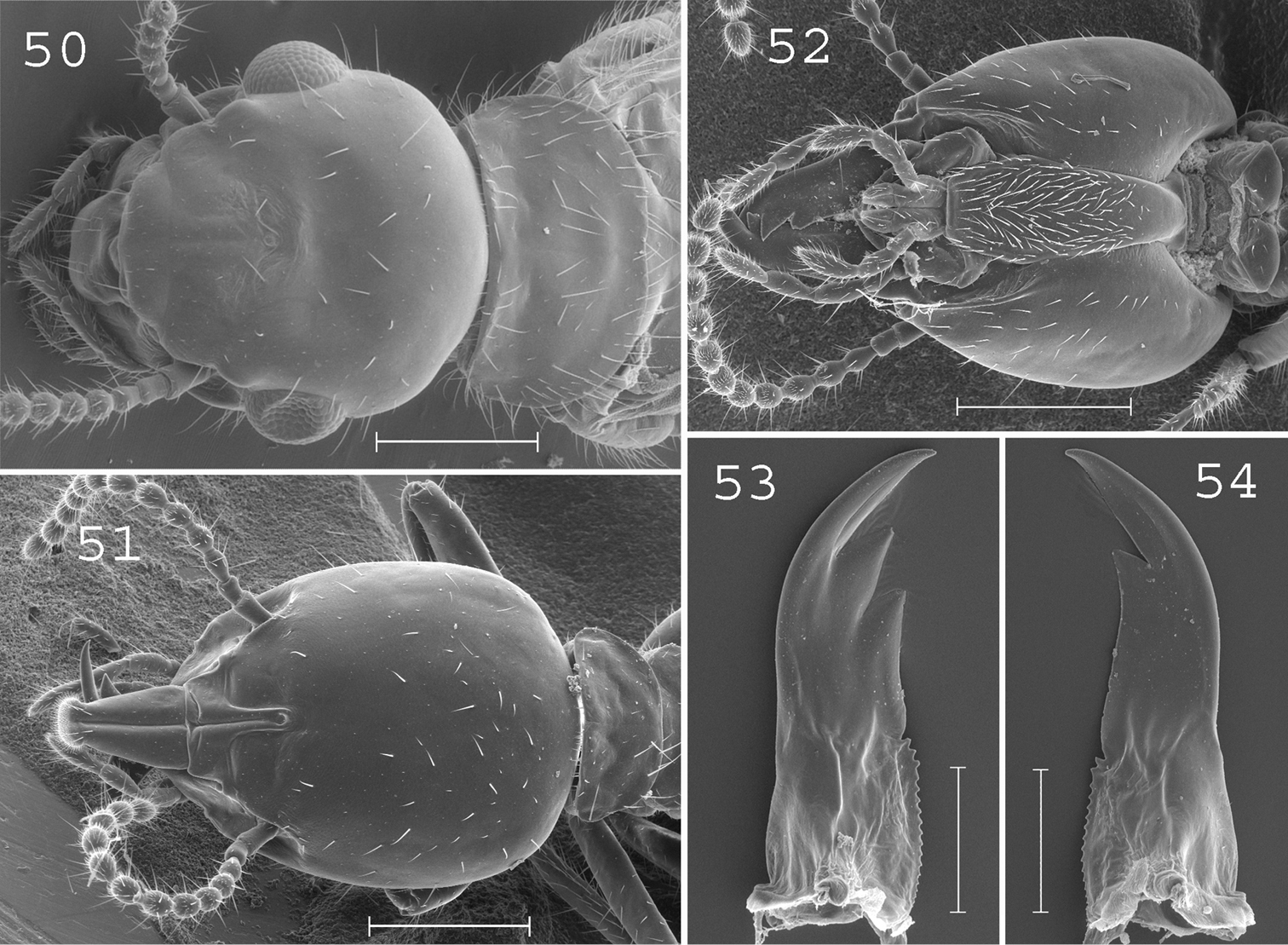
Parrhinotermes barbatus. Imago: 50 head. Soldier: 51 head in dorsal view; 52 head in ventral view; 53 left mandible; 54 right mandible. Scale bars: 50, 51, 52: 0.5mm; 53, 54: 0.2mm.
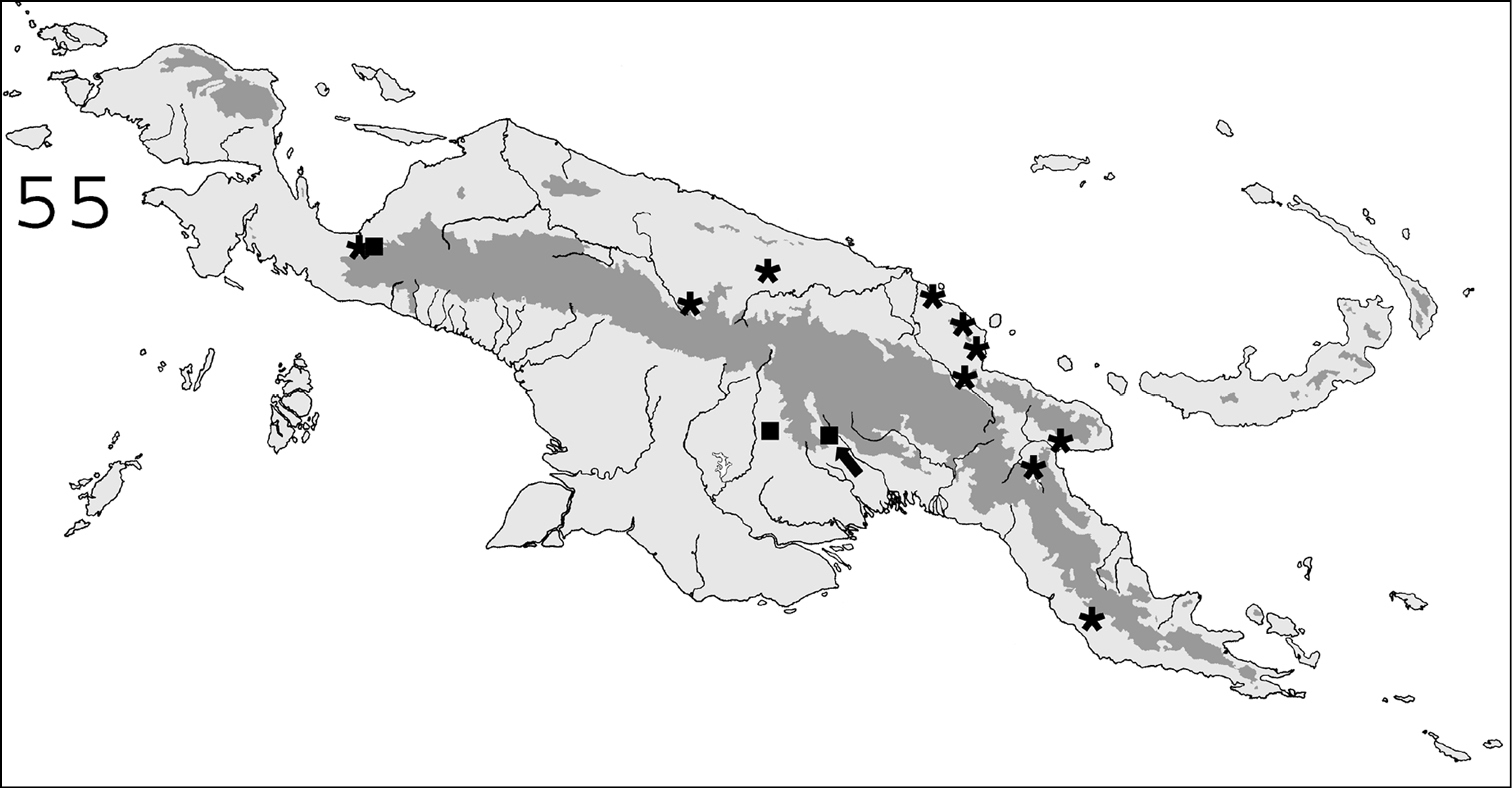
Known distribution in New Guinea of: ∗ Parrhinotermes browni; ■ Parrhinotermes barbatus. Arrow points to type locality.
(Fig. 45). Head covered by about 15 setae with posterior edges strongly rounded, frons and clypeus short. Pronotum covered by about 100 setae. Antennae with 17 articles. Measurements (mm) of 6 imagoes from 1 colony: TBL: 5.76–6.34; HLC: 1.04–1.18; HWE: 1.19–1.26; PL: 0.47–0.65; PW: 0.90–0.95; FWL: 8.34–8.93; ED: 0.27–0.36.
(Figs 46–49) Head rectangular-shaped with edges slightly rounded, covered by about 50 small setae and 10 large ones. Labrum elongated, reaching the tip of mandibles, with a rounded apical brush. Postmentum covered by setae only on its upper part. Antennae with 13 articles. Mandibles straight, slightly curved at tip, slightly serrated at base. Measurements (mm) of 6 soldiers of the type colony and 39 soldiers from 13 colonies (parentheses): HLC: 1.13–1.19 (1.01–1.33); HLL: 1.56–1.64 (1.40–1.79); HW: 0.90–0.94 (0.86–1.05); PW: 0.52–0.56 (0.51–0.68); RML: 0.69–0.73 (0.64–0.78); MPW: 0.27–0.31 (0.27–0.34); mPW: 0.16–0.18 (0.16–0.22); T3L: 0.75–0.81 (0.70–0.89).
(Fig. 55). This species, originally described from the Solomon Islands, is common in Northern New Guinea and in the bird’s neck area.
Several species of Trichopseniini (Coleptera, Staphylinidae) were reported from this species (
urn:lsid:zoobank.org:act:10EADEB0-140F-46D1-BEED-EFD6E6FA8C01
http://species-id.net/wiki/Parrhinotermes_barbatus
Figs 50– 55Soldier: PAPUA NEW GUINEA: Southern Highlands: Lake Kutubu, 11.x.1988 (YR) (#PNGT1285). Paratypes: PAPUA NEW GUINEA: Southern Highlands: Lake Kutubu, 11.x.1988 (YR), two colonies, one with alates, the other one with late nymphs (#PNGT1285, 1286); Lake Kutubu, 12.x.1988 (YR), with late nymphs (#PNGT1292); Lake Kutubu, 13.x.1988 (YR), with late nymphs (#PNGT1296); Pimaga, 16.x.1988 (YR) (#PNGT1306); Pimaga, 17.x.1988 (YR), with late nymphs (#PNGT1315); Fly: Nomad, 31.v.1990 (YR & ML), two colonies in dead wood, with queen (#PNGT1628, 1630); Nomad, 31.v.1990 (YR & ML) (#PNGT1650). INDONESIA: Papua: Road Nabire-Mapia km 62, 18.xi.1995 (YR) (#IRJT71).
(Fig. 50). Head covered by about 15 setae with posterior margin strongly rounded, frons and clypeus of medium size. Pronotum covered by about 100 setae (Fig. 52). Antennae with 17 articles. Measurements (mm) of 6 imagoes from 1 colony: TBL: 5.23–6.02; HLC: 1.06–1.10; HWE: 1.24–1.27; PL: 0.44–0.65; PW: 0.85–0.98; FWL: 8.02–8.84; ED: 0.27–0.39.
(Figs 51–54). Head ellipsoid, covered by about 50 small setae. Labrum elongated, reaching the tip of mandibles, ending in a rounded brush. Postmentum covered by setae from the base to the upper part. Antennae with 13 articles. Mandibles straight, slightly curved at tip, slightly serrated at base. Measurements (mm) of 30 soldiers from 10 colonies: HLC: 1.11–1.33; HLL: 1.51–1.75; HW: 0.95–1.12; PW: 0.53–0.65; RML: 0.66–0.79; MPW: 0.25–0.34; mPW: 0.16–0.22; T3L: 0.74–0.90.
This species is distinguishable from Parrhinotermes browni and Parrhinotermes queenslandicus Mjöberg, 1920 by the postmentum of soldiers, completely covered by setae.
(Fig. 55). This species is common in southern New Guinean forests, and was also collected once in Indonesian Papua.
The four species of Parrhinopsenius found with Parrhinotermes browni were also found with this species, previously referred to as Parrhinotermes nr. queenslandicus (
We named this species after the latin “barba”, referring to the postmentum of its soldiers fully covered by setae.
http://species-id.net/wiki/Schedorhinotermes
Rhinotermes intermedius Brauer, 1865, by original designation.
Imagoes very similar to those of Parrhinotermes. Head approximately circular in shape. Fontanelle situated in the middle of the head. Frons with a slightly visible groove. Labrum short, inclined downward, without groove. Soldiers generally dimorphic and sometimes trimorphic. All species described here have dimorphic soldiers, excepted Schedorhinotermes seclusus in which the minor soldiers can sometimes be further separated into two morphs (
This genus is known from Africa, Southeast Asia, the Papuan region and Australia (
http://species-id.net/wiki/Schedorhinotermes_seclusus
Figs 56–61, 82Lectotype: AUSTRALIA: Queensland: Babinda, 06.i.1925 (G.F. Hill) (NMVA #T-10854). Other material: PAPUA NEW GUINEA: Southern Highlands: Lake Kutubu, 13.x.1988 (YR) (#PNGT1295); Fly: Morehead, 25.iii.1989 (YR & ML) (#PNGT1438); Morehead, 26.iii.1989 (YR & ML) (#PNGT1453); Wipim, 15.viii.1962 (R. Zweifel), witth nymphs (AMNH); Wipim, 30.iii.1989 (YR & ML) (#PNGT1480, 1482); Wipim, 31.iii.1989 (YR & ML) (#PNGT1495); Wipim, 01.iv.1989 (YR & ML) (#PNGT1504); Lake Murray, 22.v.1990 (YR & ML) (#PNGT1558, 1559, 1560); Lake Murray, 23.v.1990 (YR & ML) (#PNGT1573, 1576); Lake Murray, 24.v.1990 (YR & ML) (#PNGT1579); Lake Murray, 25.v.1990 (YR & ML) (#PNGT1596); Lake Murray, 27.v.1990 (YR & ML) (#PNGT1604); Nomad, 31.v.1990 (YR & ML) (#PNGT1633).
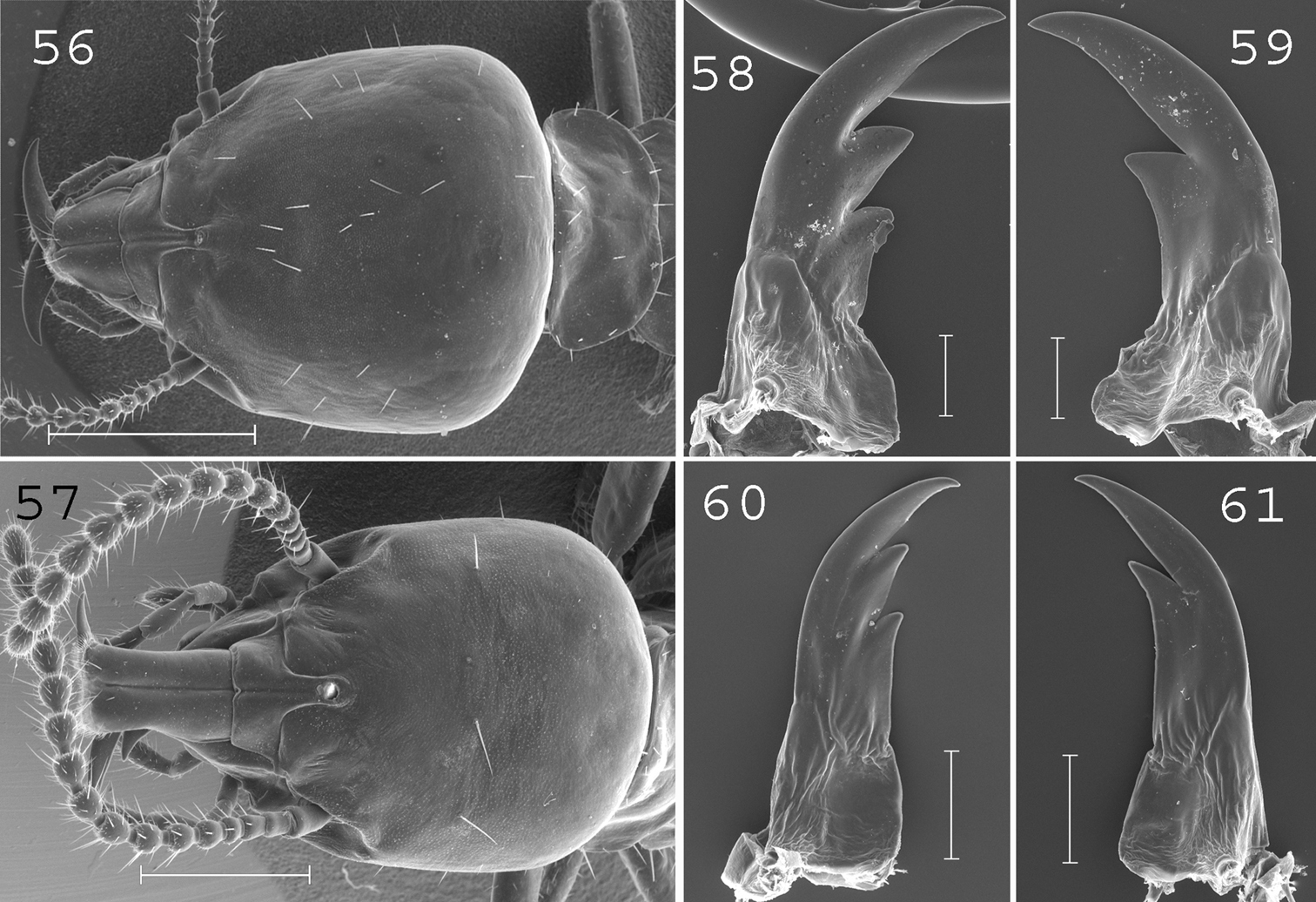
Schedorhinotermes seclusus. Major soldier: 56 head; 58 left mandible; 59 right mandible. Minor soldier: 57 head; 60 left mandible; 61 right mandible. Scale bars: 56, 57: 0.5mm; 58, 59, 60, 61: 0.2mm.
Unknown.
(Figs 56, 58–59) Soldiers of large size. Head covered by about 30 setae. Labrum short and large, not reaching the tip of mandibles. Antennae generally with 17 articles, sometimes with 16 or 18 articles. Pronotum large, covered by about 15 setae principally situated on the edges. Abdomen densely hairy, with 20 to 40 setae per segment. Mandibles moderately curved. Left mandible with the first subsidiary tooth shorter than the second and with a small hump at the base. Right mandible with a slight hump at the base. Measurements (mm) of 1 major soldier from the type colony and 30 major soldiers from 10 colonies (parentheses): HLC: 2.07 (1.82–2.33); HLL: 2.52 (2.24–2.74); HW: 1.96 (1.69–2.05); PW: 1.27 (1.08–1.42); RML: 1.27 (1.00–1.36); mPW: 0.36 (0.22–0.38); T3L: 1.76 (1.43–1.73).
(Figs 57, 60–61). Soldiers of large size. Head covered by about 15 setae. Labrum 1.5 times longer than wide, reaching the tip of mandibles. Antennae with 16 articles. Pronotum of large size with about 10 setae on the edges. Abdomen abundantly hairy, with 10 to 15 setae per segment. Large mandibles strongly curved. Measurements (mm) of 1 minor soldier from the type colony and 10 minor soldiers from 10 colonies (parentheses): HLC: 1.11 (1.10–1.41); HLL: 1.52 (1.59–1.97); HW: 0.86 (0.95–1.19); PL: 0.44 (0.38–0.48); PW: 0.66 (0.68–0.86); RML: 0.75 (0.79–0.97); MPW: 0.35 (0.34–0.39); T3L: 1.09 (1.01–1.32).
This species is easily distinguishable from others by its large size and its densely hairy abdomen in the minor and major soldier castes. Minor soldiers are highly variable in size, indicating the likely presence of two developmental subcategories (
(Fig. 82). This species is widespread throughout southern Papua New Guinea. It is also known from Queensland, Australia.
Three species of Trichopseniini (Coleoptera, Staphylinidae, Aleocharinae) were reported by
http://species-id.net/wiki/Schedorhinotermes_malaccensis
Figs 62–67, 82Paratype: NHRS collections. Other material: INDONESIA: Papua: Pusppenssat-IrJa, 13.xi.1995 (YR) (#IRJT15); Road Nabire-Mapia km 48, 15.xi.1995 (YR) (#IRJT43).
Unknown.
(Figs 62, 64–65). Soldiers of large size. Head rounded, as long as large, covered by about 20 setae. Labrum short and large. Antennae generally with 16 articles. Pronotum large, covered by about 15 setae situated on the edges. Mesonotum and metanotum covered by about 10 setae on posterior margin. Abdomen covered by about 10 setae per segment. Mandibles very large. Right mandible with large hump at the basis. Measurements (mm) of 6 major soldiers from 2 colonies: HLC: 1.70–2.09; HLL: 2.03–2.43; HW: 1.95–2.04; PW: 1.17–1.25; RML: 1.07–1.15; mPW: 0.26–0.34; T3L: 1.54–1.64.

Schedorhinotermes malaccensis. Major soldier: 62 head; 64 left mandible; 65 right mandible. Minor soldier: 63 head; 66 left mandible; 67 right mandible. Scale bars: 62, 63: 0.5mm; 64, 65, 66, 67: 0.2mm.
(Figs 63, 66–67). Head covered by 5 to 10 setae. Labrum moderately elongated, almost reaching the tip of mandibles. Fronto-clypeus short and wide. Antennae with 15 or 16 articles. Pronotum with about 10 setae placed on the edges. Mesonotum and metanotum with about 8 setae on the posterior edge. Mandibles elongated, slender, with short subsidiary teeth. Measurements (mm) of 1 minor soldier from the type colony and 6 minor soldiers from 2 colonies (parentheses): HLC: 1.28 (1.01–1.22); HLL: 1.77 (1.52–1.78); HW: 1.00 (0.81–0.95); PL: 0.50 (0.37–0.49); PW: 0.70 (0.59–0.73); RML: 0.85 (0.72–0.87); MPW: 0.36 (0.30–0.35); T3L: 1.18 (1.03–1.14).
This species is easily distinguished from other New Guinean species by the large rounded head and stout mandibles of its major soldiers.
(Fig. 82). This species is widespread throughout Sundaland (
http://species-id.net/wiki/Schedorhinotermes_longirostris
Figs 68–74, 82Syntypes, minor soldier and workers: INDIA: Nicobar Islands: Kondul Island (NHRS). Syntypes of Schedorhinotermes dimorphus, major soldiers, minor soldiers and workers: PAPUA NEW GUINEA: Madang: Madang (as Friedrich-Wilhelmshafen), 8.i.1901 (L. Biró) (IRSNB). Other material: PAPUA NEW GUINEA: Sandaun: Vanimo, 08.iii.1994 (YR & ML) (#PNGT1718); East Sepik: Marangis, 07.iii.1983 (JMP & YR) (#PNGT253); Madang: Road Madang-Lae km 30, 04.vii.1981 (JMP) (#PNGT120, 121); Bunapas road, 07.vii.1981 (JMP) (#PNGT132, 141); Potsdam plantation, 23.iii.1983 (YR) (#PNGT275); Nubia, 18.v.1983 (YR) (#PNGT353), Potsdam plantation, 20.vi.1983 (YR) (#PNGT386); Bunapae, 25.x.1983 (YR) (#PNGT415); Sepen No.1, 29.x.1983 (YR) (#PNGT425); Potsdam plantation, 25.xi.1983 (YR), with nymphs (#PNGT479); Bunapae, 12.vi.1984 (YR) (#PNGT680); Potsdam plantation, 20.vii.1984 (YR) (#PNGT743); Bunapae, 23.vii.1984 (YR) (#PNGT748); Potsdam plantation, 24.vii.1984 (YR) (#PNGT753); Hansa point, 22.viii.1984 (YR) (#PNGT786); Hansa point, 08.ix.1984 (YR) (#PNGT823); Sepen No.1, 16.ii.1985 (JMP & YR), with alates (#PNGT876); Sepen No.1, 01.iii.1988 (YR) (#PNGT1165, 1166); Yagaum hospital, 10.iv.1983 (YR) (#PNGT304); Gilagil River bridge, 12.iii.1988 (YR) (#PNGT1170); Baitabag, 15.v.1999 (L. Čižek) (#2, J. Šobotník’s collection); Tabobo, 07.i.1989 (ML) (#PNGT1383); Road Kausi-Bundi, 07.v.1988 (YR) (#PNGT1207); Bundi, 10.v.1988 (YR) (#PNGT1210); Morobe: Wampit, 06.ii.1983 (JMP & YR) (#PNGT178); Kaiapit, 19.ii.1983 (JMP & YR) (#PNGT192); Sirasira, 15.v.1988 (YR) (#PNGT1219, 1220). Bulolo, 14.ii.1983 (JMP & YR) (#PNGT168); Mount Susu, 23.v.1987 (YR), in branch of klinkii pine on the ground (#PNGT1085); Mount Susu, 19.v.1988 (YR), in branch of klinkii pine on the ground (#PNGT1233); Oro: Kokoda, 13.iii.1985 (JMP & YR), in tree stump in cocoa plantation (#PNGT949); Koiasi, 14.iii.1985 (JMP & YR) (#PNGT957); Kokoda, 19.iii.1985 (JMP & YR), in dead wood in rubber plantation (#PNGT987); East New Britain: Warengoi, 19.v.1984 (JMP & YR) (#PNGT561); Ataliklikun Bay, 30 km W of Keravat, 23.v.1984 (JMP & YR) (#PNGT595); New Ireland: Lelet plateau, 27.v.1984 (JMP & YR) (#PNGT609, 610); Konos, 29.v.1984 (JMP & YR) (#PNGT623); Manus: Road Lorengau-Yiringo km 32, 04.vi.1984 (JMP & YR) (#PNGT659); Central: Sogeri area, 23.xi.1962 (AE), 2 samples with alates, from logs in rubber grove (AMNH); Sirinumu Dam, 06.iii.1985 (JMP & YR) (#PNGT913); Sirinumu Dam, 08.iii.1985 (JMP & YR) (#PNGT926, 931); Southern Highlands: Lake Kutubu, 11.x.1988 (YR) (#PNGT1287, 1288); Pimaga, 16.x.1988 (YR) (#PNGT1304); Fly: Tabubil, 20.v.1990 (YR & ML) (#PNGT1548); Lake Murray, 22.v.1990 (YR & ML) (#PNGT1557); Lake Murray, 24.v.1990 (YR & ML) (#PNGT1583); Nomad, 31.v.1990 (YR & ML) (#PNGT1626). INDONESIA: Papua: Road Nabire-Mapia km 43, 26.xi.1995 (YR) (#IRJT168); Kaimana, 21.xi.1995 (YR), three samples, the last one with large nymphs (#IRJT103, 109, 113); Kaimana, 23.xi.1995 (YR) (#IRJT142).
We compared the type material of Schedorhinotermes longirostris and Schedorhinotermes dimorphus with our material and were not able to find any relevant morphological characters to distinguish the two species. Therefore, we consider Schedorhinotermes dimorphus as a junior synonym of Schedorhinotermes longirostris.
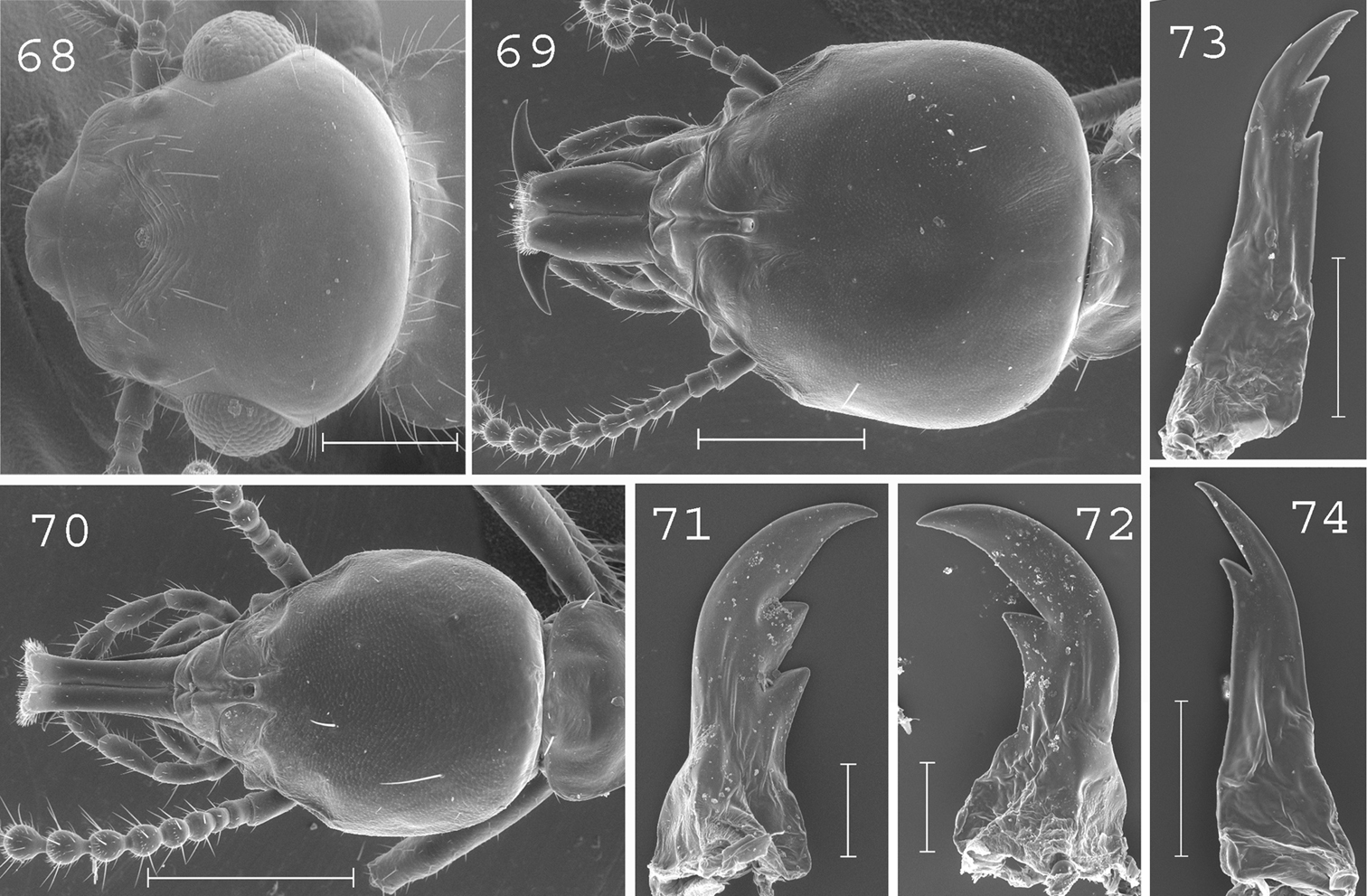
Schedorhinotermes longirostris. Imago: 68 head. Major soldier: 69 head; 71 left mandible; 72 right mandible. Minor soldier: 70 head; 73 left mandible; 74 right mandible. Scale bars: 68, 69, 70: 0.5mm; 71, 72, 73, 74: 0.2mm.
(Fig. 68). Head covered by 10–15 setae. Eyes of medium size. Pronotum covered by about 50 setae mainly located on the edges. Antennae with 20 articles. Measurements (mm) of 6 imagoes from 1 colony: TBL: 7.39–8.79; HLC: 1.37–1.58; HWE: 1.51–1.62; PL: 0.73–0.79; PW: 1.23–1.36; FWL: 8.70–9.80; ED: 0.31–0.33.
(Figs 69, 71–72). Soldiers of small size. Head, excluding labrum, square-shaped, slightly longer than wide, covered by about 10 setae. Labrum slightly longer than wide, reaching the tip of mandibles. Antennae with 15 or 16 articles. Pronotum covered by 6 long setae disposed in each corners. Mesonotum and metanotum with 4 long setae. Abdomen with 6 long setae per segment, plus sometimes 1 or 2 smaller ones. Mandibles strongly curved at tip with short subsidiary teeth. Left mandible with 2 subsidiary teeth of the same length. Right mandible without hump at the basis. Measurements (mm) of 1 major soldier from the type colony of Schedorhinotermes longirostris, 2 major soldiers from the type colony of Schedorhinotermes dimorphus [brackets], and 77 major soldiers from 29 colonies (parentheses): HLC: 1.56 [1.50–1.56] (1.35–1.95); HLL: 2.05 [1.92–1.99] (1.65–2.21); HW: 1.32 [1.31–1.35] (1.17–1.51); PW: 0.74 [0.73–0.75] (0.65–0.86); RML: 0.98 [0.89–0.97] (0.75–1.06); mPW: 0.25 [0.24] (0.19–0.30); T3L: 1.19 [1.09–1.16] (1.11–1.36).
(Figs 70, 73–74). Soldiers of small size. Head elongated, covered by 5–10 setae. Labrum very elongated, 3 times longer than wide, reaching beyond mandibles. Fronto-clypeus of medium size. Antennae with 15 or 16 articles. Pronotum covered by 6 large setae disposed in each corner. Mesonotum and metanotum with 4 long setae. Abdomen with 6 long setae per segment, plus sometimes 1 or 2 smaller ones. Mandibles elongated, hardly curved, with short subsidiary teeth. Measurements (mm) of 10 minor soldiers from 10 colonies of Schedorhinotermes longirostris, plus 6 minor soldiers of the type colony of Schedorhinotermes dimorphus [brackets]: HLC: 0.82–1.05 [0.90–0.94]; HLL: 1.18–1.54 [1.30–1.37]; HW: 0.60–0.77 [0.66–0.69]; PL: 0.28–0.36 [0.30–0.33]; PW: 0.40–0.53 [0.44–0.51]; RML: 0.52–0.70 [0.51–0.64]; MPW: 0.25–0.31 [0.24–0.28]; T3L: 0.74–0.94 [0.71–0.87].
This species can be distinguished from other New Guinean species by the number of setae on the pronotum (6 per segment), mesonotum (4), metanotum (4) and abdomen (6) in the major soldier caste.
(Fig. 82).This species is common in forested areas throughout New Guinea. It probably also occurs in Indonesia, since it was described from the Nicobar islands.
Several Aleocharinae (Coleoptera, Staphylinidae) were found with this species. Schedotermoecia kaimanensis (Coptotermoeciina) was described from colony #IRJT142, and its congener Schedotermoecia papuana from #PNGT1165 (
http://species-id.net/wiki/Schedorhinotermes_translucens
Figs 75–81, 83Syntypes, all castes: MALAYSIA: Sarawak: Kuching, xi.1894 (G.D. Haviland) (type No. 299, B.M.1899–41, BMNH, collection data from
Schedorhinotermes celebensis was described by
Schedorhinotermes marjoriae was described by
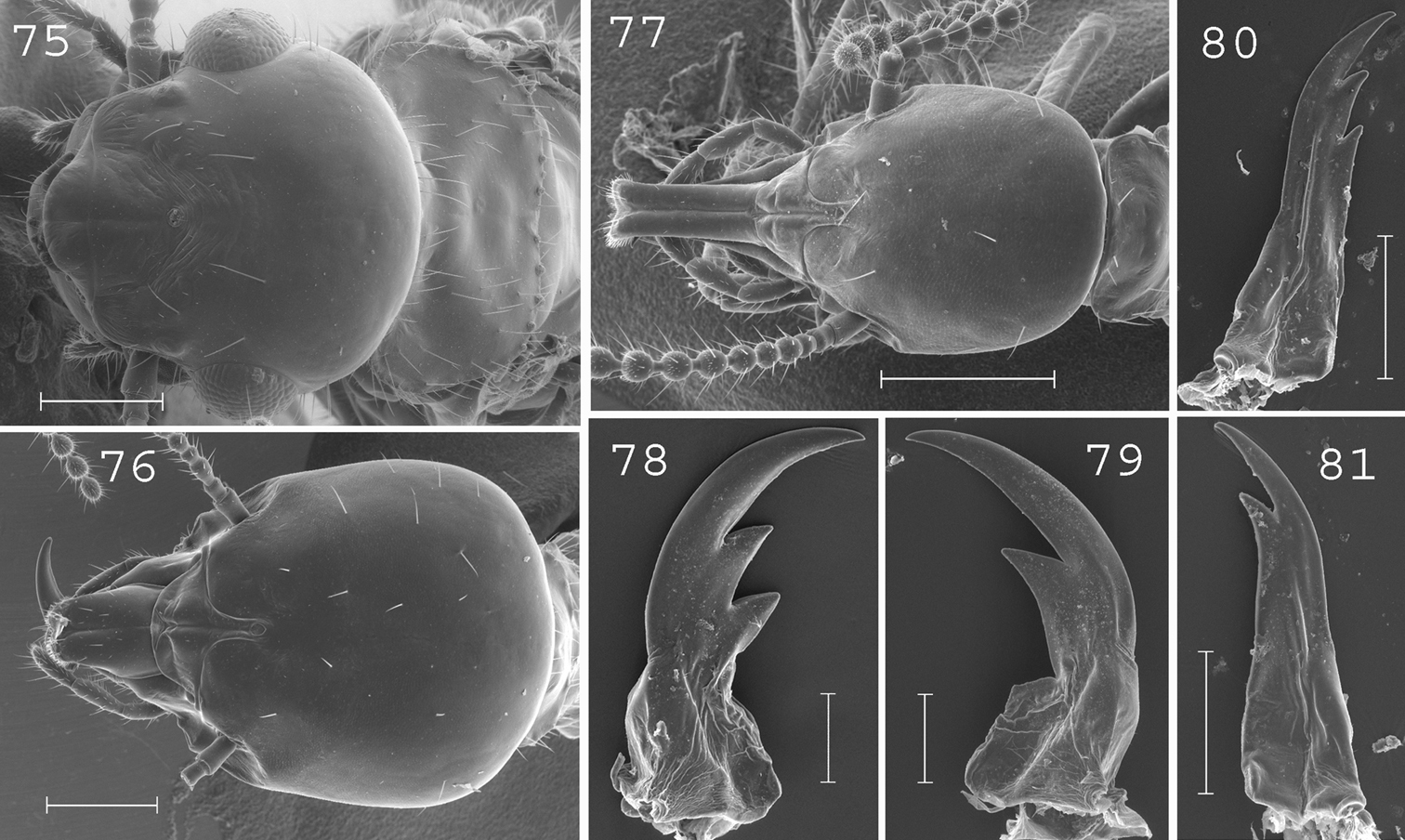
Schedorhinotermes translucens. Imago: 75 head. Major soldier: 76 head; 78 left mandible; 79 right mandible. Minor soldier: 77 head; 80 left mandible; 81 right mandible. Scale bars: 75, 76, 77: 0.5mm; 78, 79, 80, 81: 0.2mm.
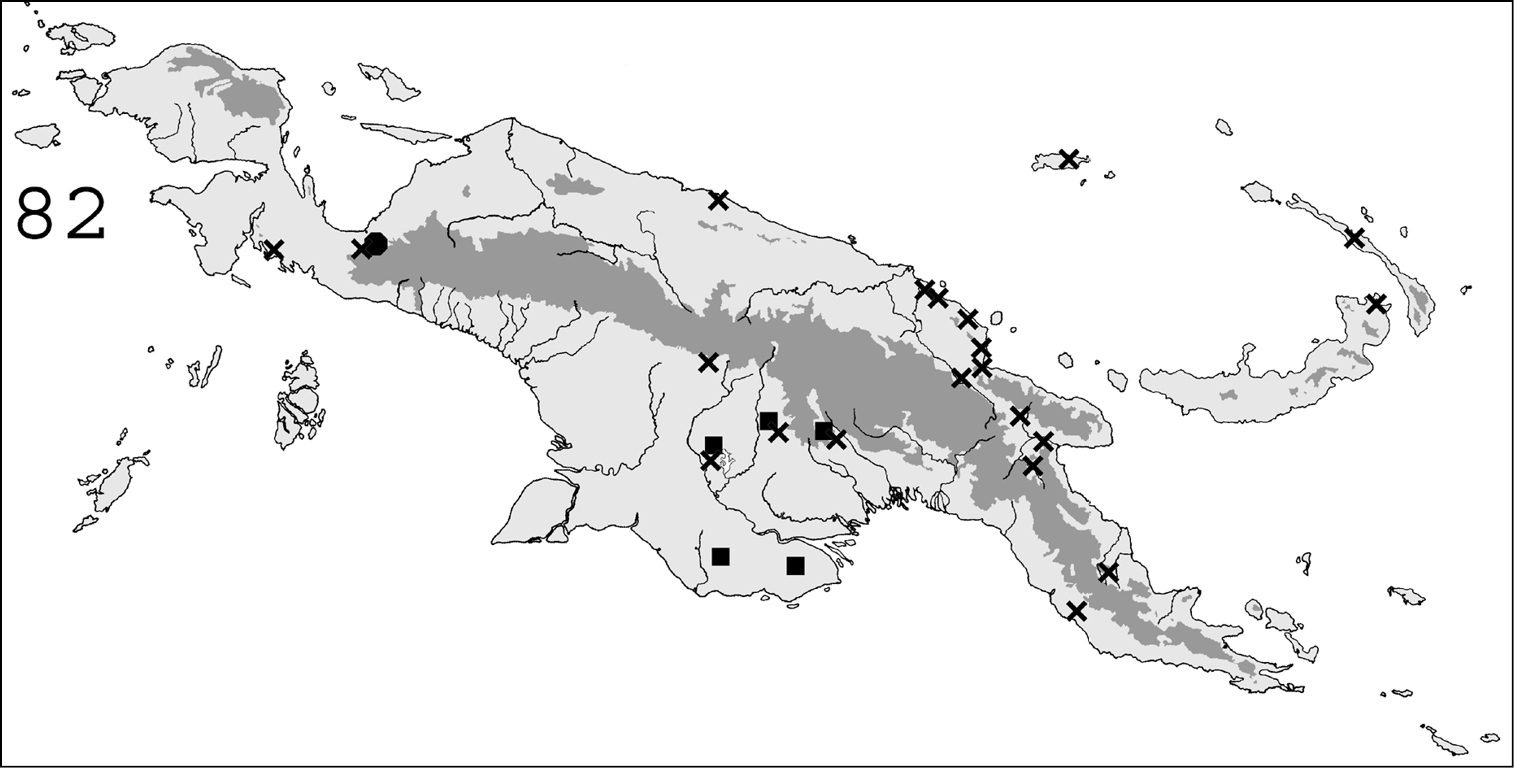
Known distribution in New Guinea of: ■ Schedorhinotermes seclusus; ● Schedorhinotermes malaccensis; ✖ Schedorhinotermes longirostris.
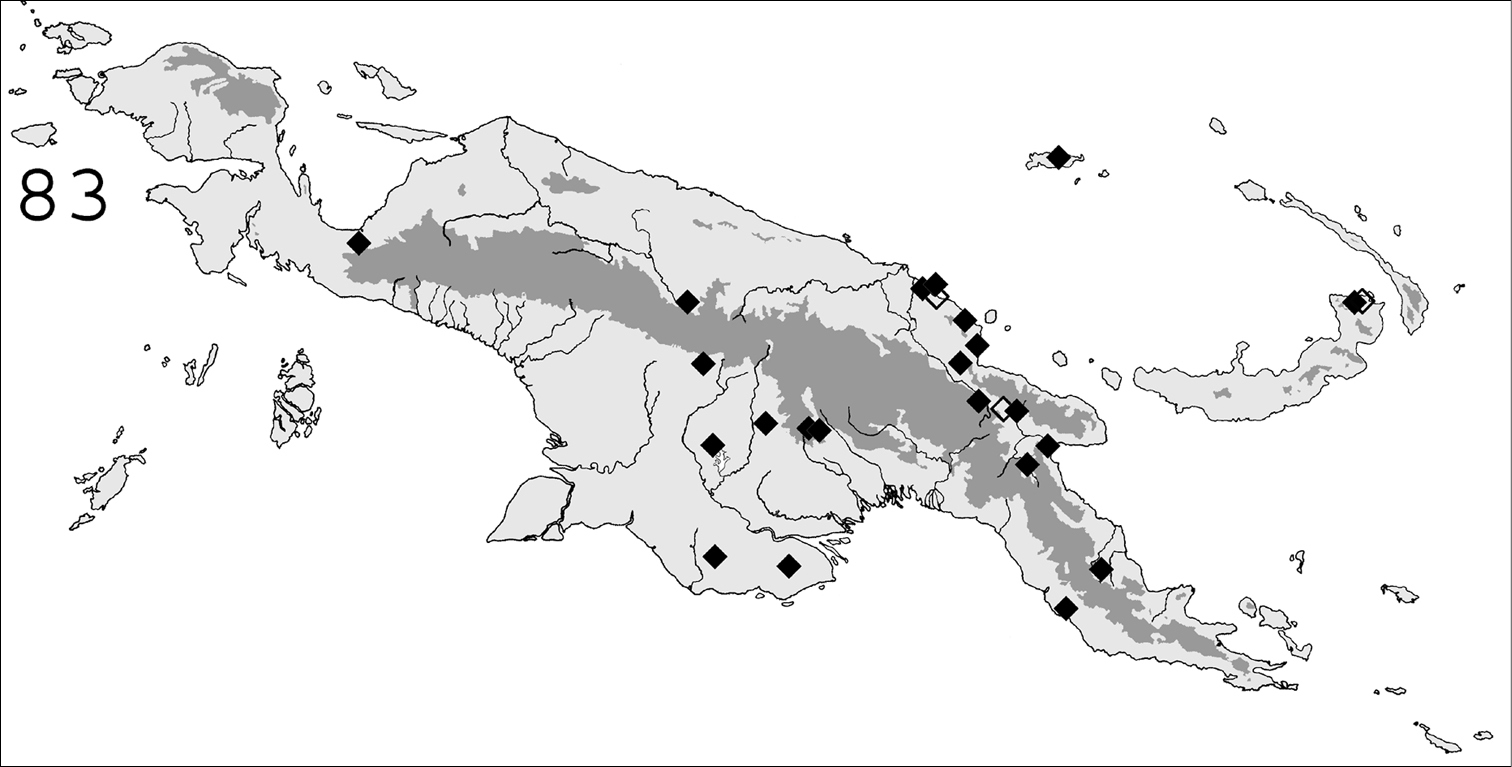
◆ Known distribution in New Guinea of Schedorhinotermes translucens; ◇ dubious samples.
(Fig. 75). Head slightly rounded posteriorly, covered by about 15 setae. Eyes relatively large. Pronotum bearing about 50 setae, principally located on edges. Antennae with 20 articles. Measurements (mm) of 3 imagoes from the type colony of Schedorhinotermes translucens, 1 imago from the type colony of Schedorhinotermes celebensis [brackets], and 26 imagoes from 5 colonies (parentheses): TBL: 7.31–8.95 [7.18] (7.02–8.95); HLC: 1.62–1.68 [1.54] (1.36–1.67); HWE: 1.72–1.76 [1.66] (1.63–1.84); PL: 0.88–0.98 [0.80] (0.74–0.89); PW: 1.48–1.55 [1.39] (1.30–1.56); FWL: 10.21–10.70 [n.a.] (9.60–11.40); ED: 0.51–0.54 [0.41] (0.31–0.48).
(Figs 76, 78–79). Soldiers of medium size. Head slightly longer than wide, covered by about 30 setae. Labrum not reaching the tip of mandibles. Antennae with 16 articles. Pronotum large, covered by about 15 setae. Mesonotum and metanotum covered by about 10 setae on the posterior margin. Abdomen with 15 to 20 setae per segment. Left mandible with the first subsidiary tooth slightly longer than the second. Right mandible with well developed outgrowth on interior side of base. Measurements (mm) of 3 major soldiers from the type colony of Schedorhinotermes translucens, 6 major soldiers from one determined sample of Schedorhinotermes marjoriae [brackets], and 87 major soldiers from 29 colonies (parentheses): HLC: 1.91–1.97 [1.95–2.06] (1.45–2.10); HLL: 2.38–2.42 [2.40–2.58] (1.82–2.62); HW: 1.67–1.79 [1.75–1.89] (1.32–1.84); PW: 1.02–1.08 [1.04–1.12] (0.74–1.17); RML: 1.11–1.14 [1.12–1.19] (0.88–1.23); MPW: 0.27–0.30 [0.29–0.34] (0.20–0.34); T3L: 1.54–1.57 [1.42–1.53] (1.15–1.63).
(Figs 77, 80–81). Head elongated, rounded posteriorly, covered by about 10 setae. Labrum 2.5 times longer than wide, reaching the tip of mandibles. Fronto-clypeus elongated. Antennae with 15 articles. Pronotum with about 10 setae on the edges. Mesonotum and metanotum with about 10 setae on the posterior edge. Abdomen with about 8 to 10 setae per segment. Mandibles slender. Measurements (mm) of 5 minor soldiers from the type colony of Schedorhinotermes translucens, 6 minor soldiers of one determined sample of Schedorhinotermes marjoriae [brackets] and 10 minor soldiers from 10 colonies (parentheses): HLC: 0.99–1.19 [0.98–1.20] (0.90–1.11); HLL: 1.44–1.72 [1.49–1.67] (1.34–1.68); HW: 0.78–0.93 [0.78–0.87] (0.71–0.90); PL: 0.40–0.48 [0.35–0.46] (0.28–0.43); PW: 0.59–0.69 [0.52–0.63] (0.45–0.65); RML: 0.65–0.78 [0.60–0.72] (0.55–0.77); MPW: 0.30–0.35 [0.28–0.32] (0.24–0.34); T3L: 1.04–1.24 [0.89–1.02] (0.87–1.04).
This species is related to Schedorhinotermes longirostris, from which it can be distinguished by its more hairy pronotum, mesonotum, metanotum and abdomen of major soldiers.
(Fig. 83). Schedorhinotermes translucens is widespread throughout New Guinea, both in savannas and forests. The following additional records are from the literature (samples not examined):
Myrmedonota termitophila (Coleoptera, Staphylinidae, Aleocharinae, Lomechusini) was discovered in colony #PNGT163 (
http://species-id.net/wiki/Prorhinotermes
Prorhinotermes inopinatus Silvestri, 1909, by original designation.
Imago head oval to circular-shaped, with ocelli located before the well developed eyes. Fontanelle situated in the middle of the head. Antenna with 19 to 22 articles. Pronotum narrower than head. Soldier head variable in shape, often larger posteriorly than anteriorly. Fontanelle narrow, placed at anterior third of the head. Frons with a groove in the middle from opening of fontanelle to clypeus. Eyes present as hyaline spots, more or less developed. Antennae with 13 to 20 articles. Pronotum generally wide. Mandibles elongated, left one with a short marginal tooth at the basis, right one without marginal teeth. Soldiers and workers very variable in size (
Prorhinotermes has an insular distribution. It is found in the West Indies, Pacific islands, East Indies and islands of the Indian Ocean including Madagascar. It is not reported from continents excepted in Central America, Southern Florida and Northern Australia (
http://species-id.net/wiki/Prorhinotermes_inopinatus
Figs 84–87, 93Syntype: TONGA: Niua Fo’ou (as Insulae Samoa, Ninafoon) (B. Friedländer) (AMNH). Other material: PAPUA NEW GUINEA: Madang: Bunapas road, 26.vi.1981 (JMP), with neotenic reproductives (#PNGT111); Road Bogia-Tangu km 10, 07.vii.1981 (JMP) (#PNGT129); Potsdam, 18.iv.1983 (YR) (#PNGT325); Nubia, 18.v.1983 (YR) (#PNGT354); Sepen No.1, 29.x.1983 (YR), with nymphs and neotenics (#PNGT426); Bunapae, 12.vi.1984 (YR) (#PNGT687); Bunapae, 23.vii.1984 (YR), with queen in dead wood (#PNGT749); Potsdam, 24.vii.1984 (YR), with neotenics (#PNGT754); Hansa point, 05.ix.1984 (YR) (#PNGT808); Tabele (Manam Is.), 19.ix.1984 (YR) (#PNGT840); Laing Island, 06.ii.1985 (JMP & YR), with alates and neotenics (#PNGT857); Nubia, 25.v.1986 (YR), two colonies, the former dissected completely, with two primary queens and one neotenic male, and nymphs (#PNGT1012, 1013); Potsdam, 03.vi.1986 (YR), two colonies, the latter with many alates (#PNGT1019, 1020); Sepen No.1, 06.vii.1986 (YR) (#PNGT1035); Nubia, 12.vii.1986 (YR), two small colonies (#PNGT1039, 1040); Road Potsdam-Makarup km?, 30.viii.1987 (YR) (#PNGT1137); Sepen No.1, 26.iv.1988 (YR), with two neotenic females (#PNGT1192); Laing Island, 12.iii.1989 (ML), in log on sea shore, with alates (#PNGT1405); Hatzfeldthafen, 20.v.1983 (YR) (#PNGT362); Tabobo, 23.ii.1983 (JMP & YR), with royal pair (#PNGT219); Yagaum Hospital, 07.ii.1983 (JMP & YR), with neotenics (#PNGT149); Yagaum Hospital, 12.iv.1983 (YR), two colonies, the former with alates (#PNGT310, 311); Yagaum Hospital, 13.v.1983 (YR) (#PNGT346); Baitabag, xi.1999 (L. Čižek) (#12, J. Šobotník’s collection). Morobe: 21 km NW Lae, 08.xii.1962 (AE), from log on ground in lowland forest (AMNH); Lae, 11.xii.1962 (P. Aloma & AE), rather wet log on forest floor in Botanic Garden (AMNH); 5 km S Lae, 21.xii.1962 (P. Aloma), 2 samples in mangrove branches and stump in water (AMNH); 8 km NW Lae, 27.xii.1962 (AE) (AMNH); 32 km NW Lae (Markham road), 27.xii.1962 (AE), in standing dead tree on edge of Pandanus forest (AMNH); 32 km SW Lae, 29.xii.1962 (AE), in sago forest (AMNH); Oomsis, 24.v.1988 (YR) (#PNGT1241); Sandaun: Yapsiei, 12.iii.1994 (YR & ML) (#PNGT1753); Fly: Tabubil, 20.v.1990 (YR & ML) (#PNGT1539); Nomad, 29.v.1990 (YR & ML), two colonies, the former with neotenics (#PNGT1608, 1612); Nomad, 01.vi.1990 (YR & ML), (#PNGT1635). INDONESIA: Papua: Pusppenssat-IrJa, 13–15.xi.1995 (YR), three colonies, the first two with neotenics (#IRJT16, 27, 44); Pusppenssat-IrJa, 18.xi.1995 (YR) (#IRJT76); road Nabire-Mapia km 43, 26.xi.1995 (YR) (#IRJT172); Pusppenssat-IrJa, 01.xii.1995 (YR) (#IRJT213). Kaimana, 23.xi.1995 (YR), in dead log on limestone hill (#IRJT145).
(Fig. 85). Head slightly rounded, covered by about 10 setae. Eyes variable in size. Pronotum with large setae mainly situated on the edges. Measurements (mm) of 12 imagoes from 2 colonies: TBL: 4.38–6.72; HLC: 1.09–1.33; HWE: 1.20–1.47; PL: 0.65–0.81; PW: 1.09–1.27; FWL: 6.67–8.42; ED: 0.24–0.37.

Prorhinotermes inopinatus. Soldier: 84 head; 86 left mandible; 87 right mandible. Imago: 85 head and pronotum. Scale bars: 84, 85: 0.5mm; 86, 87: 0.2mm.
(Figs 84, 86–87). Head slightly elongated, covered by about 15 setae. Eyes of medium size. Pronotum, mesonotum and metanotum long and wide. Abdomen covered by 6 to 12 long setae per segment. Large mandibles, slightly curved at tip. Measurements (mm) of syntype and 94 soldiers from 32 colonies (parentheses): HLC: 1.79 (1.39–1.85); HLL: 2.02 (1.65–2.16); HW: 1.10 (1.13–1.53); PL: 0.56 (0.45–0.69); PW: 1.17 (0.86–1.24); RML: 1.22 (0.95–1.40); MPW: 0.51 (0.37–0.57); T3L: 1.35 (1.02–1.48).
(Fig. 93). This species occurs throughout New Guinean forests, or forested swamps (
http://species-id.net/wiki/Termitogeton
Termes planus Haviland, 1898, by monotypy.
Imagoes densely hairy. Head larger posteriorly than anteriorly. Fontanelle very narrow, placed in the middle of the head. Eyes small. Antennae with 10 to 15 articles. Pronotum very small, half as broad as head. Wings without median and radial vein (
Termitogeton is a rainforest-dwelling wood feeder known from Sri Lanka, Borneo, Peninsular Malaysia (
http://species-id.net/wiki/Termitogeton_planus
Figs 88– 93Syntypes: MALAYSIA: Sarawak: Santubong, 16.ix.1894 (G.D. Haviland) (type No. 164, CUMZ, collection data from
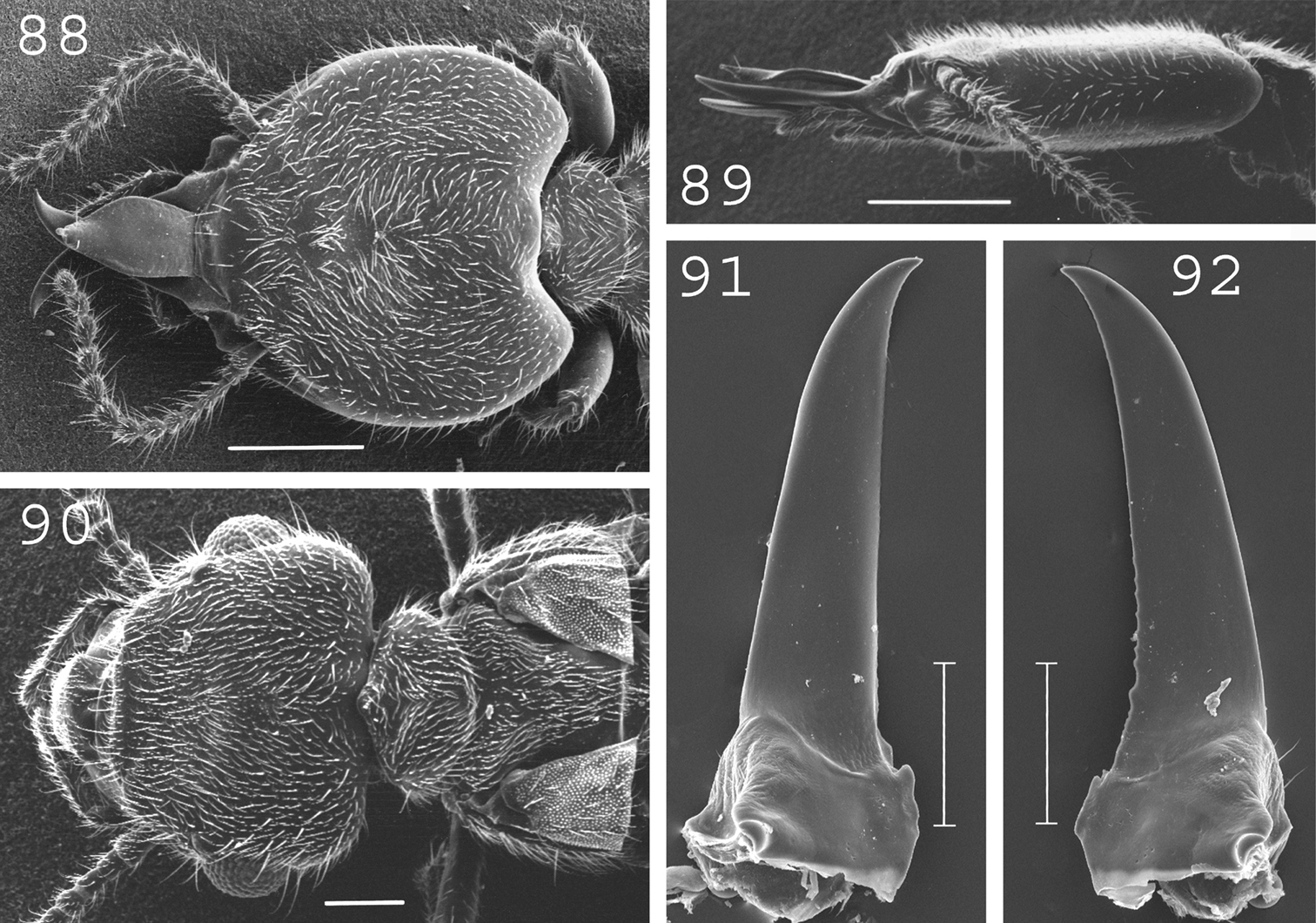
Termitogeton planus. Soldier: 88 head and pronotum in dorsal view; 89 head in lateral view; 91 left mandible; 92 right mandible. Imago: 90 head, pro- and mesonotum. Scale bars: 88, 89: 0.5mm; 90, 91, 92: 0.2mm.
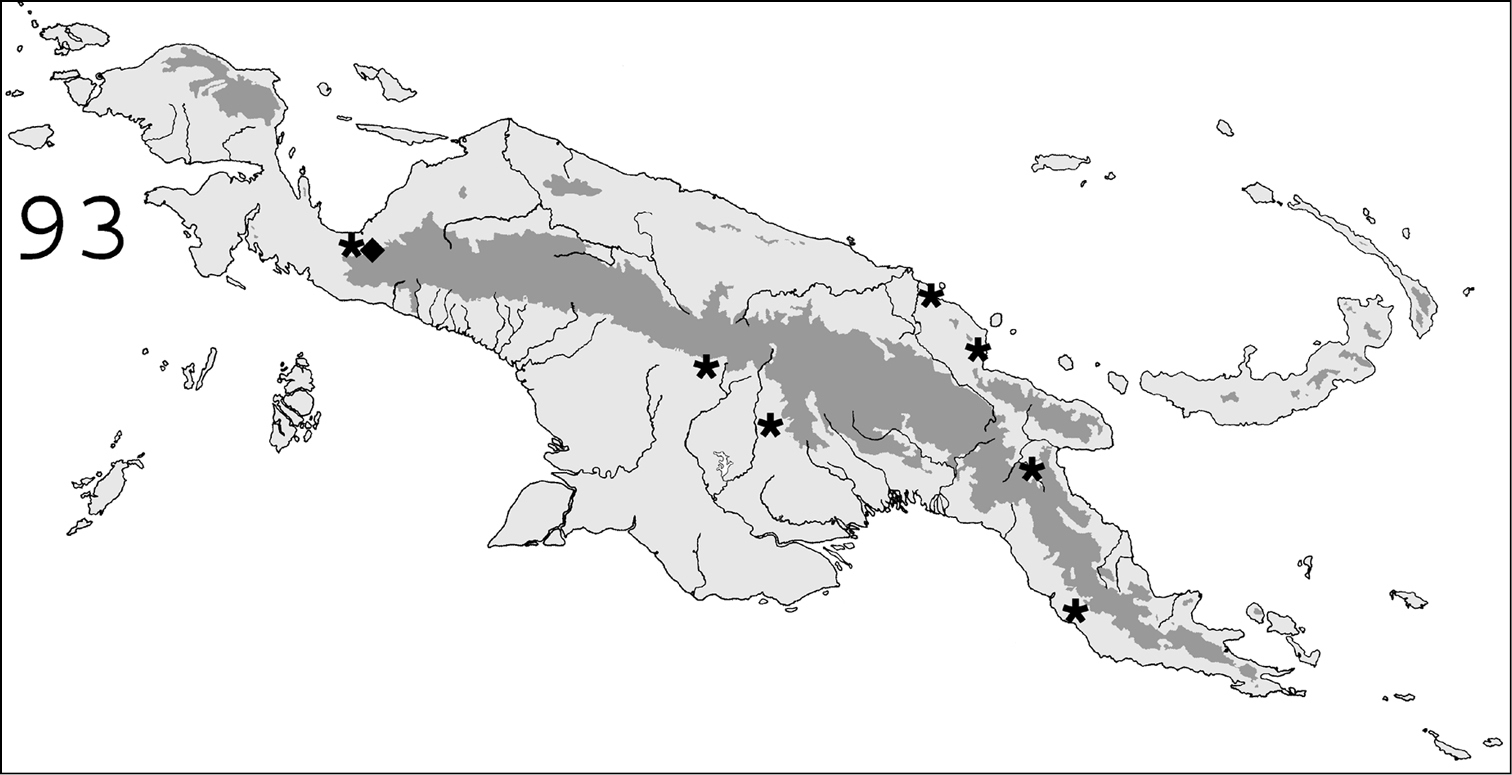
Known distribution in New Guinea of: ∗ Prorhinotermes inopinatus; ◆ Termitogeton planus.
(Fig. 90). Very small. Overall body covered by many small setae. Head narrower anteriorly than posteriorly, heart-shaped. Large eyes. Ocelli in front of eyes. Antennae generally with 9 to 15 articles. Pronotum approximately half as broad as head, with a small projection forward. Measurements (mm) of 4 imagoes from the type colony and 10 imagoes from 2 colonies (parentheses): TBL: 3.23–3.94 (3.19–3.94); HLC: 0.59–0.63 (0.59–0.70); HWE: 0.82–0.85 (0.85–0.93); PL: 0.27–0.28 (0.28–0.30); PW: 0.39–0.45 (0.43–0.51); FWL: n.a. (4.93–5.23); ED: 0.14–0.18 (0.18–0.23).
(Figs 88–89, 91–92). Overall body covered by many small setae. Head heart-shaped. Labrum elongated, pointed at the tip, narrow at the basis, broadest apical one-third. Antennae with 11 to 15 articles. Pronotum half as broad as head, elongated anteriorly in the middle. Mandibles short, without marginal teeth, slightly curved at tip. Measurements (mm) 4 soldiers from the type colony and 32 soldiers from 8 colonies (parentheses): HLC: 1.16–1.27 (1.10–1.37); HW: 1.21–1.31 (1.12–1.48); PW: 0.51–0.55 (0.45–0.58); LML: 0.71–0.73 (0.75–0.86); PML: 0.87–0.94 (0.77–0.97); mPW: 0.10–0.13 (0.11–0.15); T3L: 0.58–0.62 (0.51–0.65).
(Fig. 93).In New Guinea, this species was only collected in the bird’s neck area (western Indonesian Papua). Originally described from from Sarawak (
| 1 | Mandibles sabre-like, without subsidiary teeth (Figs 14–15) | 2 |
| – | Left and right mandibles with 2 and 1 subsidiary teeth respectively (Figs 53–54) | 10 |
| 2 | Head rounded to ovoid, with broad fontanelle opening in front (Fig. 12) | Coptotermes 3 |
| – | Head of different shape, with dot-like fontanelle distant from clypeal border | 7 |
| 3 | Head rounded, fontanelle well visible from above, mandibles strongly curved at tip (Figs 6, 7, 9) | Coptotermes elisae |
| – | Soldier head of different shape, fontanelle directed forwards | 4 |
| 4 | Head distincly narrowing anteriorly (Figs 18, 22) | 5 |
| – | Head ovoid (Figs 12, 26) | 6 |
| 5 | Small-sized soldier (HW < 1.10 mm), mandibles slightly curved (Figs 22–25) | Coptotermes pamuae |
| – | Large-sized soldier (HW > 1.15) mm, mandibles distinctly curved (Figs 18–21) | Coptotermes grandiceps |
| 6 | Mandibles curved at tips (Figs 14, 15) | Coptotermes remotus |
| - | Mandibles almost not curved at tips (Figs 28, 29) | Coptotermes gambrinus |
| 7 | Soldier flattened with heart-shaped head (Fig. 88) | Termitogeton planus |
| – | Soldier not flattened, head ovoid or with parallel sides, with posterior margin not indented (Figs 32, 37, 84) | 8 |
| 8 | Soldier head with sparse setae (Fig. 84) | Prorhinotermes inopinatus |
| – | Soldier head covered by hundreds of setae (Figs 32, 37) | Heterotermes 9 |
| 9 | Head with a small hump anteriorly (Fig. 33); labrum long and pointed, reaching well beyond half length of mandibles | Heterotermes vagus |
| – | Head with a large hump anteriorly (Fig. 38); labrum rounded and short, ending well before half length of mandibles | Heterotermes paradoxus |
| 10 | Monomorphic soldiers with serrated mandible base (Figs 48, 49) | Parrhinotermes 11 |
| - | Polymorphic soldiers, mandible base not serrated | Schedorhinotermes 12 |
| 11 | Postmentum completely covered by setae (Fig. 52) | Parrhinotermes barbatus |
| – | Postmentum covered by setae only in the anterior part (Fig. 47) | Parrhinotermes browni |
| 12 | Pronotum and abdomen of major soldiers with 6 setae per segment | Schedorhinotermes longirostris |
| – | Pronotum and abdomen of major soldiers with 10 or more setae per segment | 13 |
| 13 | Major soldier head rounded, with stout mandibles (Fig. 62) | Schedorhinotermes malaccensis |
| – | Major soldier head somewhat flattened on sides and posterior margin (Figs 56, 76) | 14 |
| 14 | Abdomen of major soldier with more than 20 setae per segment | Schedorhinotermes seclusus |
| – | Abdomen of major soldier with less than 20 setae per segment | Schedorhinotermes translucens |
This study reveals that the richness of Rhinotermitidae in New Guinea is much higher than the 8 species previously recognized. Overall, we found 6 genera and 15 species, a diversity which appears slightly lower than that of neighboring areas such as Peninsular Malaysia (6 genera, 24 species (
The distribution patterns of rhinotermitid species match those found in termitids (
We expect the rhinotermitid diversity found in this study to reflect the overall richness of the island, although it is inevitable that some rare or locally distributed taxa escaped detection. Two regions are particularly likely to host undiscovered taxa: (i) southern Papuan savannas, in which our collecting effort was limited, possibly host additional taxa of Australian affinities; (ii) as stated above, the western half of New Guinea (Indonesian Papua) was also poorly explored and is therefore likely to conceal further taxa of Oriental origin, in the vein of Termitogeton planus and Schedorhinotermes malaccensis.
Thanks are due to museum curators who kindly loaned us type material, with a special mention to Kumar Krishna (AMNH) who also allowed us access to samples from the Emersons’ journey through Lae and Port Moresby in 1962–1963. J.M. Pasteels and M. Leponce contributed to several collection campaigns. J. Cillis provided technical assistance with SEM operations. Financial support was provided by the Belgian National Fund for Scientific Research (F.R.S.–FNRS), through several grants to YR and a predoctoral fellowship to TB.
Situation of collecting localities in Papua New Guinea and Indonesian Papua