





(C) George E. Ball. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Using for comparison with, and as outgroups for, supertribe Cicindelitae, we describe and illustrate the mandibles and labrum-epipharynx of the basal geadephagans Trachypachus gibbsii LeConte, 1861 (family Trachypachidae), and family Carabidae: Pelophila rudis (LeConte, 1863) (supertribe Nebriitae, tribe Pelophilini) and Ceroglossus chilensis (Eschscholtz, 1829) (supertribe Carabitae, tribe Ceroglossini). The range and pattern of variation in structure of mandibles and labrum-epipharynx within the supertribe Cicindelitae was assessed using scanning-electron (SEM) images of these structures in nine exemplar taxa: Amblycheila baroni (Rivers, 1890), Omus californicus (Eschscholtz, 1829) and Picnochile fallaciosa (Chevrolat, 1854) (representing the Amblycheilini); Manticora tuberculata (DeGeer, 1778) (representing the Manticorini): Tetracha carolina (Linnaeus, 1767) (representing the Megacephalini); Pogonostoma chalybeum (Klug, 1835) (representing the Collyridini); and Therates basalis Dejean, 1826, Oxycheila species, and Cicindela longilabris Say, 1824 (representing the Cicindelini). An evolutionary transformation series was postulated for the mandibles and labrum-epipharynx, based on a reconstructed phylogenetic sequence, which, in turn, was based on morphological and DNAevidence.Principal features of the transformation series for the mandibles included development of a densely setose basal face; wide quadridentate retinaculum; a lengthened incisor tooth; a multidentate terebra (one to five teeth; two-three most frequent), followed by subsequent loss of one or more such teeth; development of a diastema in the occlusal surface; development and subsequent loss of scrobal setae, and reduction and loss of the scrobe. Principal features of the transformation series for the labrum included evolution of form from transverse, sub-rectangular to elongate almost square, to triangular; position and number of setae evolved from dorsal to insertion on the apical margin, the number increased from 8-10 to as many as 36, and decreased to as few as four. The epipharynx broadened evolutionarily, the pedium evolving in form from narrow, triangular and nearly flat, to broad, palatiform, and markedly convex; anterior parapedial setae both increased and decreased in number, and in orientation, from a row parallel to the parapedial ridge to a setal row extended forward at about a right angle to the latter.
comparative morphology, mouthparts, pre-oral mill, evolution, Coleoptera, Trachypachidae, Carabidae, Carabitae, Nebriitae, Cicindelitae
In their review of variation in mandibular structure within the coleopteran suborder Adephaga,
Our initial investigation of tiger beetle mandibles revealed for the Geadephaga a combination of unique features and in them, substantial variation. To understand these aspects, we sought an orienting principle in the relatively recent studies and postulates of tiger beetle evolution, admirably summarized by Pearson and Vogler (2001: 43-51). We turned to analyses (Maddison et al 1999) of related basal stocks of geadephagans to seek the antecedents of the tiger beetle mandibles and labrum-epipharynx. Tiger beetles have been placed as a supertribe (Cicindelitae) within the Carabidae (
In this paper, we illustrate and describe the mandibles and labrum-epipharynx of tiger beetles representing the five tribes here recognized (Amblycheilini, Manticorini, Megacephalini, Collyridini, and Cicindelini). We arrange them in a phylogenetic sequence, based on a postulated evolutionary pattern (
We are pleased to dedicate this study in comparative morphology to Ross and Joyce Bell, Department of Biology, University of Vermont, Burlington, Vermont, in recognition of their contributions to the study of tiger beetles, the use of mandibles as character systems, and more generally to the field of adephagan systematics (e.g.,
We examined 12 specimens with SEM, and an additional 37 with light (Wild M5 and M3 stereoscope) microscopy, (Table 1, Appendix). These specimens are housed in the E. H. Strickland Entomological Museum, University of Alberta (UASM), Royal Alberta Museum, Edmonton, Alberta (RAMC), California Academy of Science, San Francisco, California (CASC), John H. Acorn Collection, Edmonton, Alberta (JHAC), and Ronald L. Huber Collection, Bloomington, Minnesota (RLHI).
Names, sex, and classification of exemplar individuals and species with SEM-illustrated mandibles and labrum-epipharynx.
| Family TRACHYPACHIDAE |
| Tribe TRACHYPACHIDINI |
| Trachypachus Motschulsky |
| Trachypachus gibbsii LeConte, 1861 (male) |
| Family CARABIDAE |
| Supertribe CARABITAE |
| Tribe CEROGLOSSINI |
| Ceroglossus Solier |
| Ceroglossus chilensis (Eschscholtz, 1829) (male) |
| Supertribe CICINDELITAE |
| Tribe AMBLYCHEILINI |
| Amblycheila Say |
| Amblycheila baroni (Rivers, 1890) (female) |
| Omus Dejean |
| Omus californicus (Eschscholtz, 1829) (male) |
| Picnochile Motschulsky |
| Picnochile fallaciosa (Chevrolat, 1835) (female) |
| Tribe MANTICORINI Fabricius |
| Manticora Fabricius |
| Manticora tuberculata (DeGeer, 1778) (female) |
| Tribe MEGACEPHALINI |
| Tetracha Hope |
| Tetracha carolina (Linnaeus, 1767) (male) |
| Tribe COLLYRIDINI |
| Pogonostoma Klug |
| Pogonostoma chalybeum (Klug, 1835) (male) |
| Tribe CICINDELINI |
| Therates Latreille |
| Therates basalis Dejean, 1826 (female) |
| Oxycheila Dejean |
| Oxycheila species, male |
| Cicindela Linnaeus |
| Cicindela longilabris Say, 1824 (male) |
| Supertribe NEBRIITAE |
| Tribe PELOPHILINI |
| Pelophila Dejean |
| Pelophila rudis (LeConte, 1863) (female) |
Taxon and specimen selection. For the cicindelites, we chose exemplar specimens to represent the five currently recognized tribes: Amblycheilini, Manticorini, Megacephalini, Collyridini and Cicindelini. For comparative purposes, we chose representatives from basal geadephagan lineages: Trachypachidae (Trachypachus gibbsii (LeConte, 1861)), the putative adelphotaxon of the caraboid stock; and Carabidae— Carabitae-Ceroglossini (Ceroglossus chilensis (Eschscholtz, 1829)), and Nebriitae-Pelophilini (Pelophila rudis (LeConte, 1863)). These groups of Carabidae represent different feeding types (carabites, like cicindelites, primarily predatory fluid feeders; and nebriites, particulate feeders; see Evans and Forsythe (1985: 115).
Specimen preparation. Standard techniques were used to prepare specimens for examination with the SEM. Each specimen was relaxed and cleaned in warm water and the sclerites of interest were removed from the head capsule and sputter coated with gold before SEM images were prepared. For light microscopy, specimens were relaxed, cleaned, and the mandibles spread to their fullest extent. For some but not all taxa, the labrum epipharynx was excised and point-mounted.
SEM image preparation. Three aspects for both left and right mandibles are shown: dorsal, occlusal, and ventral; lateral aspects were noted, but not illustrated. For the labrum-epipharynx, the dorsal surface is the labrum, while the ventral surface is the epipharynx, keeping in mind that the epipharynx can also be thought of as the dorsal surface of the preoral cavity.
Photographs. The mouthparts of 12 taxa examined are also illustrated in situ, with the mandibles widely spread, hopefully providing better perspective on the relative coverage of the adducted mandibles by the labrum-epipharynx for each taxon, as well as some sense of potential occlusal relationships among mandible features, and bilateral asymmetry. Photographs were taken hand-held with a Nikon D300s camera set at ISO 800 and 1/200th of a second, equipped with an AF-S Micro-Nikkor 105mm lens set at f32, a TC-17EII (1.7X) teleconverter, and the Nikon R1C1macro flash system with two flash heads, each equipped with hand made, double-layered, frosted Mylar light diffusers. These images were presented on two color plates.
Identification of structural elements. For tiger beetles (specifically Cicindela hybrida (Linnaeus, 1767))
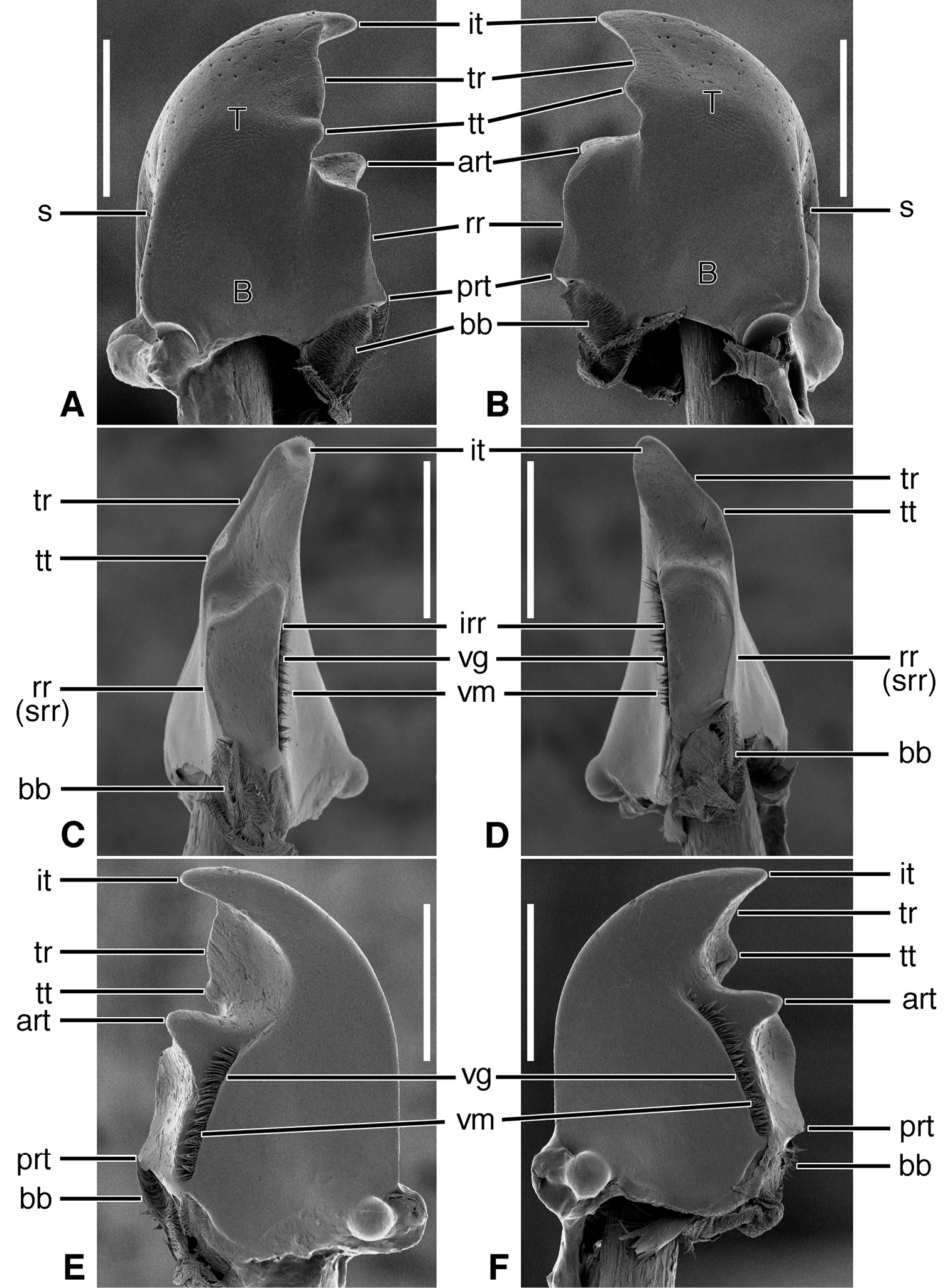
Terms and abbreviations for geadephagan mandibles (modified from
| Abbreviations | Terms |
|---|---|
| MANDIBLES | |
| art | anterior retinacular tooth |
| B | base of mandible |
| bb | basal brush |
| bfb | basal face brush |
| irr | inferior retinacular ridge |
| it | incisor tooth |
| mss | multiple scrobal setae |
| mt | molar tooth |
| od | occlusal diastema |
| prt | posterior retinacular tooth |
| rc 1 | retinacular tooth, cusp 1= art, in part |
| rc 2 | retinacular tooth, cusp 2= art, in part |
| rc 3 | retinacular tooth, cusp 3= prt, in part |
| rc 4 | retinacular tooth, cusp 4= prt , in part |
| ret | retinaculum |
| rr | retinacular ridge |
| s | scrobe |
| srr | superior retinacular ridge |
| srt | supplementary retinacular tooth |
| ss | single scrobal seta |
| T | terebra |
| tr | terebral ridge |
| tt | terebral tooth |
| tt 1 | terebral tooth 1 |
| tt 1-1 | terebral tooth 1, cusp 1 |
| tt 1-2 | terebral tooth 1, cusp 2 |
| tt 1-3 | terebral tooth 1, cusp 3 |
| tt 2 | terebral tooth 2 |
| tt 3 | terebral tooth 3 |
| tt 4 | terebral tooth 4 |
| vg | ventral groove |
| vm | ventral microtrichia |
| LABRUM-EPIPHARYNX | |
| aps | anterior parapedial setae |
| epd | epipharynx, dorsal aspect |
| las | labral apical seta |
| ped | pedium |
| pp | parapedial projection |
| pps | posterior parapedial setae |
| pr | parapedial ridge |
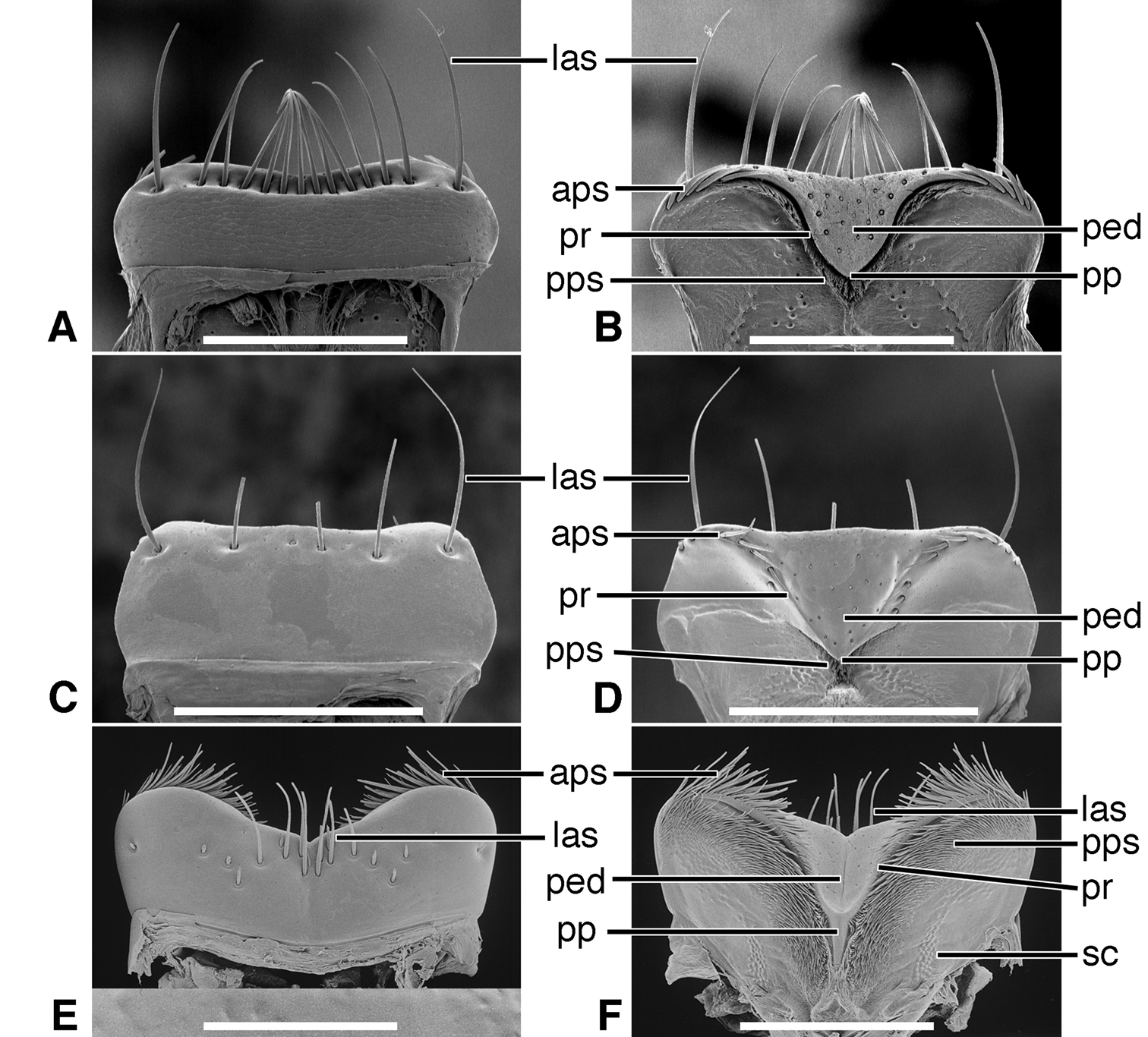
| sc | sensillum coeloconicum |
Measurements. To assist in characterizing mandibles, four measurements (Figure 1) were taken and used to make ratios (Table 2). Being based on the figures of single specimens, differences in values of these ratios have no statistical significance. They are simply a means of standardizing descriptive statements.
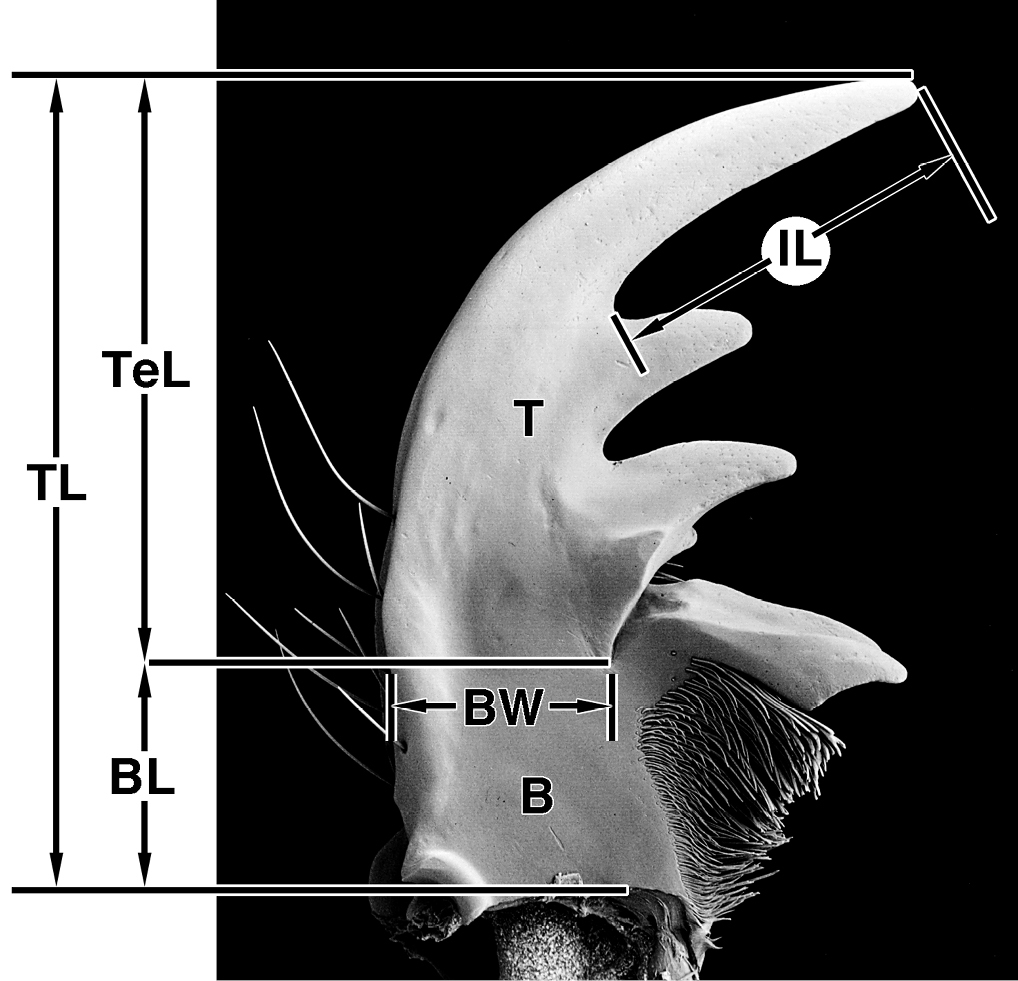
Measurements, plotted on photographic image of dorsal aspect of left mandible of Amblycheila baroni Rivers. Legend: B basal area; BL length of basal area; BW width of basal area; IL length of incisor tooth; T terebra; TeL length of terebra; TL total length.
Descriptions. Descriptions are brief, arranged in putative phylogenetic sequence, as reflected in the suprageneric taxa referred to in this paper.
Descriptions and Comparisons Family TrachypachidaeClassification. Ranked as a family, this group of two genera and fewer than 10 species may be regarded as the adelphotaxon of the Carabidae (
Exemplar taxon. Trachypachus gibbsii LeConte, 1861.
Figures 2F, 5A–B
SEM photographs of mandibles of Trachypachus gibbsi LeConte. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 0.2 mm.
Structural features. Mandibles (Figures 2A-F).Trigonal in dorso-ventral aspect, robust, basal width one half total length; curved downward slightly (Figures 2C-D). Lateral surface basally with scrobe broad, asetose, delimited by a dorsolateral and ventrolateral ridge. Terebra (T) short (terebral length one half total length), in occlusal aspect broad, with short incisor tooth (it) and with small terebral tooth (tt). Retinaculum slightly posteriad terebral tooth; short, broad, with two ridges (rr and irr), and two teeth (art and prt), anterior tooth near ventral margin (2C-D), posterior tooth dorsal. Basal face not evident. Basal brush (bb) small. Ventral surface smooth except for the ventral groove (Figures 2E-F, vg), extended anteriad anterior retinacular tooth, and with rather short and sparse microtrichia (vm).
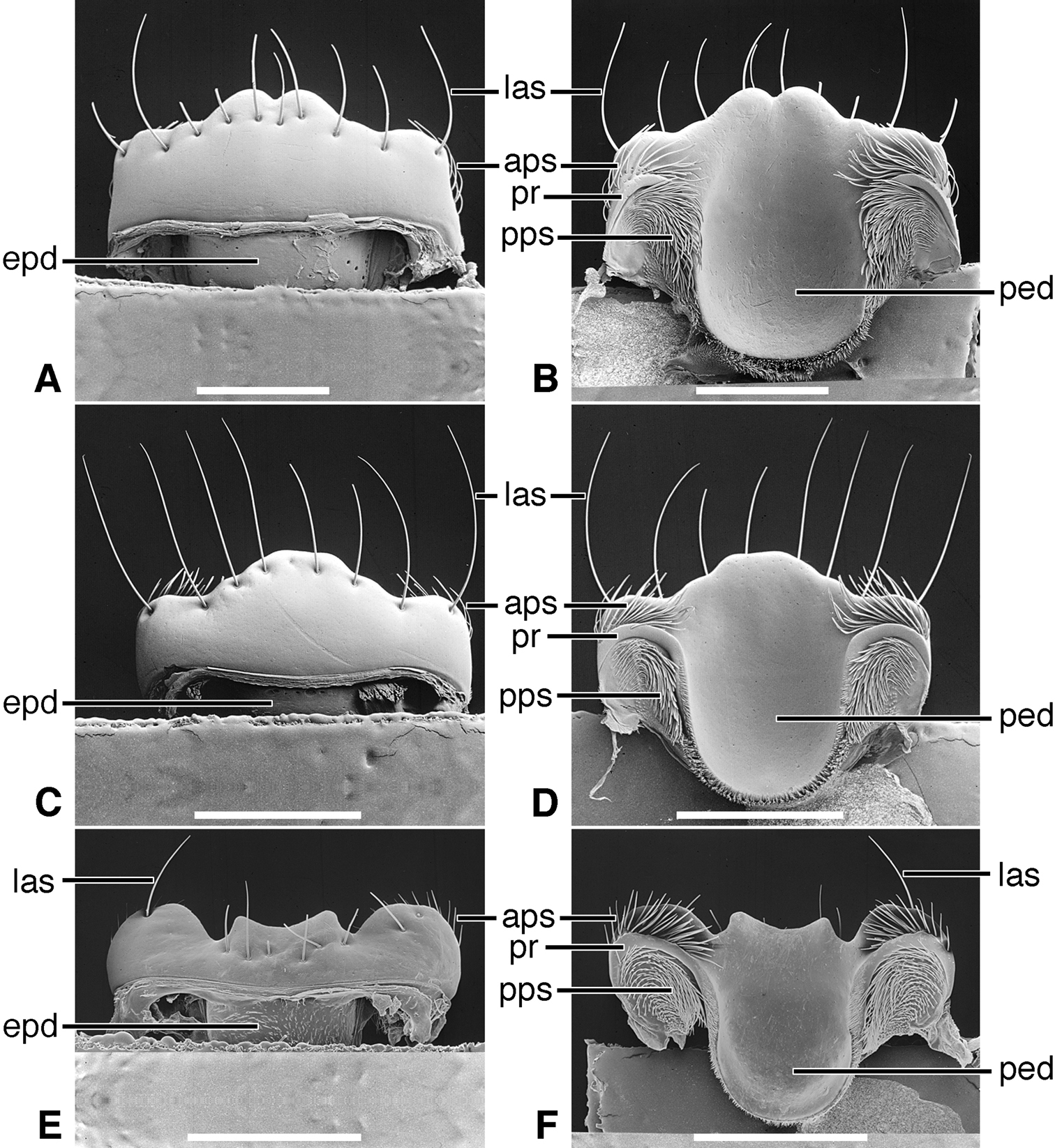
Labrum (Figure 5A). Articulated with, not immovably attached to, clypeus, labral-clypeal suture evident. Distinctly transverse, sub-rectangular, row of 14 tactile setae (las) near anterior margin, on dorsal surface (Figure 5A). Form of anterior margin subtruncate, shallowly emarginate.
Epipharynx (Figure 5B). Pedium (ped) trianguloid, apex posterior, slightly arched, with a short parapedial projection (pp), margined laterally each side by a thin parapedial ridge (pr); parapedial ridge anteriorly each side curved to lateral margin of labrum as a short lateral arm. Anterior parapedial setae (aps) in a row anterior and parallel to lateral arms of parapedial ridge. Posterior parapedial setae (pps) in a row.
Family CarabidaeTwo major types of mandibles occur among the basal carabid lineages: the fluid-feeding Ceroglossus type, and the particulate-feeding Pelophila type, evidently depending upon manipulation of food (
We identified two major types of epipharynx: the general one, shared with the Trachypachidae– pedium triangular in form, as described above; and the type confined to the tiger beetles– pedium broad, palatiform, markedly convex. For details, see below, under “Cicindelitae”.
Supertribe Nebriitae, Tribe PelophiliniClassification. This monogeneric group of two extant species is basal to the Supertribe Nebriitae, which in turn is a basal assemblage of the Carabidae (
Exemplar taxon. Pelophila rudis (LeConte, 1863).
Figures 3A-F, 5C-D

SEM photographs of mandibles of Pelophila rudis LeConte. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 0.5 mm.
Structural features. Mandibles (Figures 3A-F).Trigonal in dorso-ventral aspect, robust, basal width one third total length; curved downward (Figures 3C-D). Lateral surface basally with scrobe broad, delimited by a dorsolateral and ventrolateral ridge; scrobe triangular, moderately deep, with single seta (ss). Terebra (T) long (terebral length two thirds total length), in occlusal aspect narrow, with short incisor tooth (it) and with small terebral tooth (tt). Retinaculum slightly posteriad terebral tooth, in dorso-ventral aspect (Figures 3A-B) narrow with single ridge (rr), and two teeth (art and prt), one at each end, in line with one another. Basal face not evident. Basal brush (bb) small. Ventral surface smooth except for the ventral groove (Figures 3E-F, vg), extended anteriad anterior retinacular tooth, and with rather short and dense microtrichia (vm).
Labrum (Figure 5C). Articulated with, but not immovably attached to clypeus, labral-clypeal suture evident. Distinctly transverse, sub-rectangular, row of five tactile setae (las) near anterior margin, on dorsal surface (Figure 5C). Form of anterior margin subtruncate.
Epipharynx (Figure 5D). Pedium (ped) broadly trianguloid, apex posterior, slightly arched, with a short parapedial projection (pp); margined laterally each side by a thin parapedial ridge (pr); parapedial ridge anteriorly each side curved to lateral margin of labrum as a short lateral arm. Anterior parapedial setae (aps) in a row anterior and parallel to lateral arms of parapedial ridge. Posterior parapedial setae (pps) few, in row along parapedial ridge.
Comparisons. The prominent retinaculum seems to be a feature of the Nebriitae (see
Classification. The Supertribe Carabitae is a basal lineage (Maddison et al 1999: 104, Figure 1;
The Carabitae includes 14genera, arranged in four tribes: Cychrini; Ceroglossini; Pamborini; and Carabini. The tribe Ceroglossini is monogeneric, including eightspecies.
Exemplar taxon. Ceroglossus chilensis (Eschscholtz, 1829).
Figures 4A-F, 5E-F, Plate 1A

SEM photographs of mandibles of Ceroglossus chilensis Eschscholtz. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.
SEM photographsof labrum and epipharynx of: Trachypachus gibbsi LeConte (A labrum, dorsal aspect; B epipharynx, ventral aspect); Pelophila rudis LeConte (C labrum, dorsal aspect; D epipharynx, ventral aspect); Ceroglossus chilensis Eschscholtz (E labrum, dorsal aspect; F epipharynx, ventral aspect). Legend: see Table 2. Scale bars: A–B 0.2 mm; C–D 0.5 mm; E–F 1.0 mm).
Structural features. Mandibles (Figures 4A-F, Plate 1A).Trigonal in dorso-ventral aspect (Figures 4A-B), robust, basal width one quarter total length; curved downward (Figures 4C-D). Lateral surface basally with scrobe broad, triangular, moderately deep, asetose, delimited by a dorsolateral and ventrolateral ridge. Terebra (T) long (terebral length only one half total length, but basal area exceptionally long), in occlusal aspect (Figures 4C-D) broad; with short incisor tooth (it) and with small terebral tooth (tt). Retinaculum slightly posteriad terebral tooth, short, oriented obliquely to long axis of mandible (Figures 4C-D), (broad single ridge (rr), and two teeth (art and prt), one at each end of terebral ridge, offset from one another. Basal face long, densely setose (bfb), setae seemingly continuous with microtrichia of ventral groove (Figures 4C-D). Basal brush (bb) small. Ventral surface smooth except for the ventral groove (Figures 4E-F, vg), extended anteriorly nearly to base of the incisor tooth, and with long and dense microtrichia (vm).
Labrum (Figure 5E, Plate 1A). Immovably attached to clypeus, labral-clypeal suture evident. Distinctly transverse, sub-rectangular, transverse cluster of numerous tactile setae (las) near anterior margin, on dorsal surface (Figure 5E). Form of anterior margin moderately deeply emarginate.
Epipharynx (Figure 5F, Plate 1A). Pedium (ped) rather narrowly trianguloid, apex posterior; slightly arched, with a long parapedial projection (pp); margined laterally each side by a thin parapedial ridge (pr); parapedial ridge anteriorly each side curved gradually to lateral margin of labrum. Anterior parapedial setae (aps) in a row anterior and parallel to lateral portion of parapedial ridge. Posterior parapedial setae (pps) rather numerous, in row along parapedial ridge.
Comparisons. The form of the mouthparts in Ceroglossus is in many ways reminiscent of that in cicindelites, and a more extensive survey of the Carabitae may well uncover additional shared features.
Another group of fluid feeders, the scaritines, was examined briefly, with inconclusive results. Some, (e.g. Pasimachus Bonelli, Mouhotia Laporte de Castelnau) appear not to possess a retinaculum, whereas in Scarites the retinaculum is either reduced and confluent with the base of the terebra, or the basal portion of the terebra is multidentate and vaguely reminiscent of what we interpret here as the retinaculum of cicindelites. Some evidence exists for a relationship between tiger beetles and scaritines (the “CPRS quartet” of
Classification. Based principally on the phylogenetic conclusions of Pearson and Vogler (2001: 46, Figure 3.5) the tiger beetles are arranged here in five tribes: Amblycheilini; Manticorini; Megacephalini; Collyridini; and Cicindelini. Gálian et al. (2002: 1794, Figure 1) indicate the Megacephalini as polyphyletic, based on their study of multiple sex chromosomes in the cicindelites, the problem taxa being the oxycheiline genera Oxycheila and Cheiloxya. Based on their 18sRNA evidence and the DNA evidence of Vogler and Barraclough (1998: 255, Figure 1), these genera (and presumably Pseudoxycheila) belong in the tribe Cicindelini, and such a transfer obviates the taxonomic problem.
Structural Features. Mandibles (Figures 6A-F–8A-F, 10A-F–12A-F, and 14A-F–16A-F).Trigonal in dorso-ventral aspect, robust to slender, basal width one fifth to one third total length; planar to curved downward, ventral curvature simple to complex. Lateral surface basally with scrobe broad, delimited by a dorsolateral and ventrolateral ridge, or with ventrolateral ridge only distinct, or lateral surface convex, without a scrobe; lateral surface asetose or multisetose (ss). Terebra (T) elongate (terebral length one half to three quarters total length), in occlusal aspect broad to slender, with long incisor tooth (it) (one third to one half total length), and with or without terebral teeth (tt) (in most taxa teeth two to three, variously prominent, unicuspidate or tricuspidate; in most taxa, sexually dimorphic). Retinaculum slightly posteriad basal terebral tooth or more widely separated by a distinct gap (the occlusal diastema, od; cf. Figures 12A, B, E, F). short, broad, with four or more cusps (rc 1–4); cusps of various sizes, in some taxa as large and prominent as terebral teeth. Basal face (bf) posteriad retinaculum, with long setae in form of a dense brush (bfb). Ventral surface smooth except for the ventral groove (vg) of various lengths, and with rather long and dense microtrichia (vm).
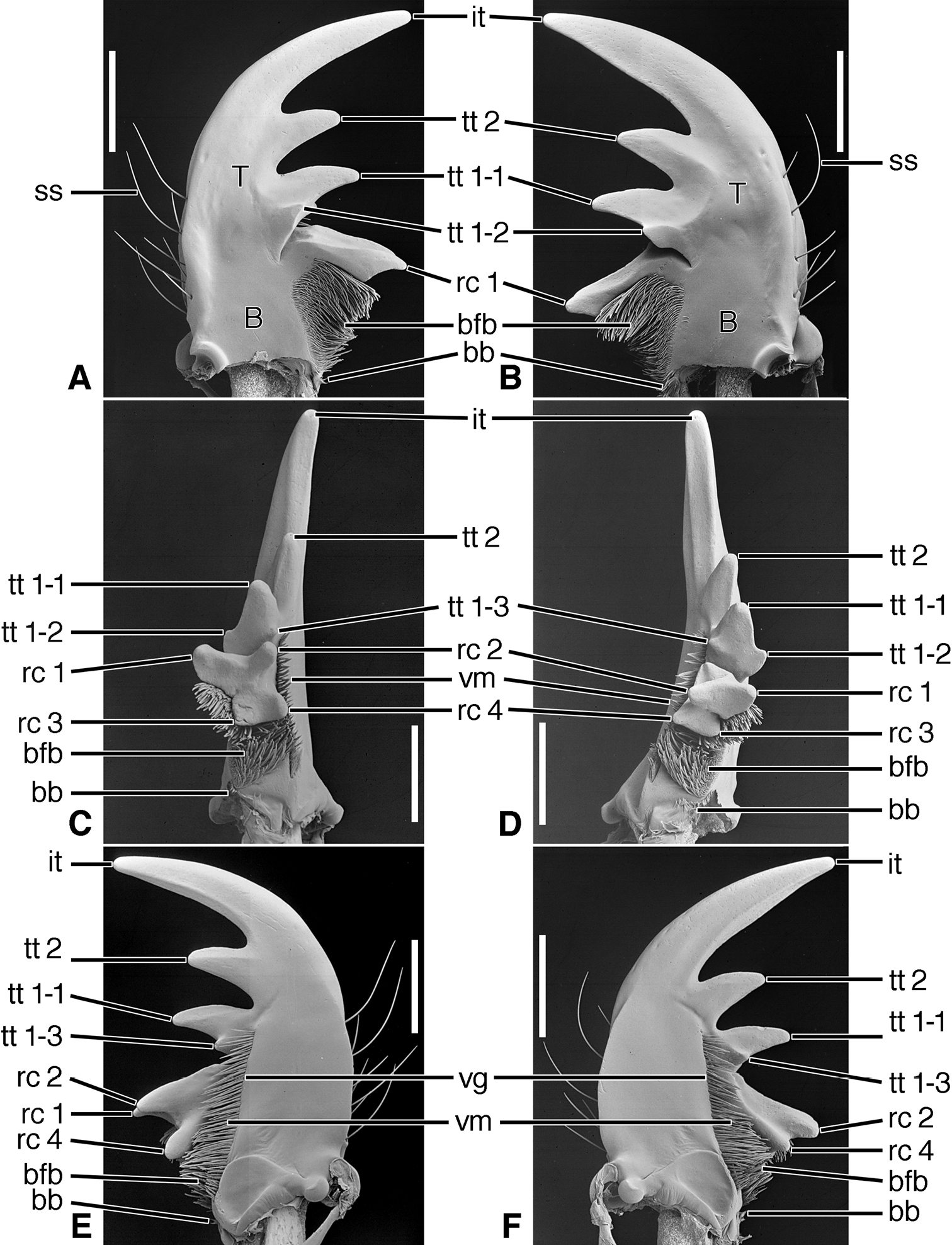
SEM photographs of mandibles of Amblycheila baroni Rivers. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars 1.0 mm.
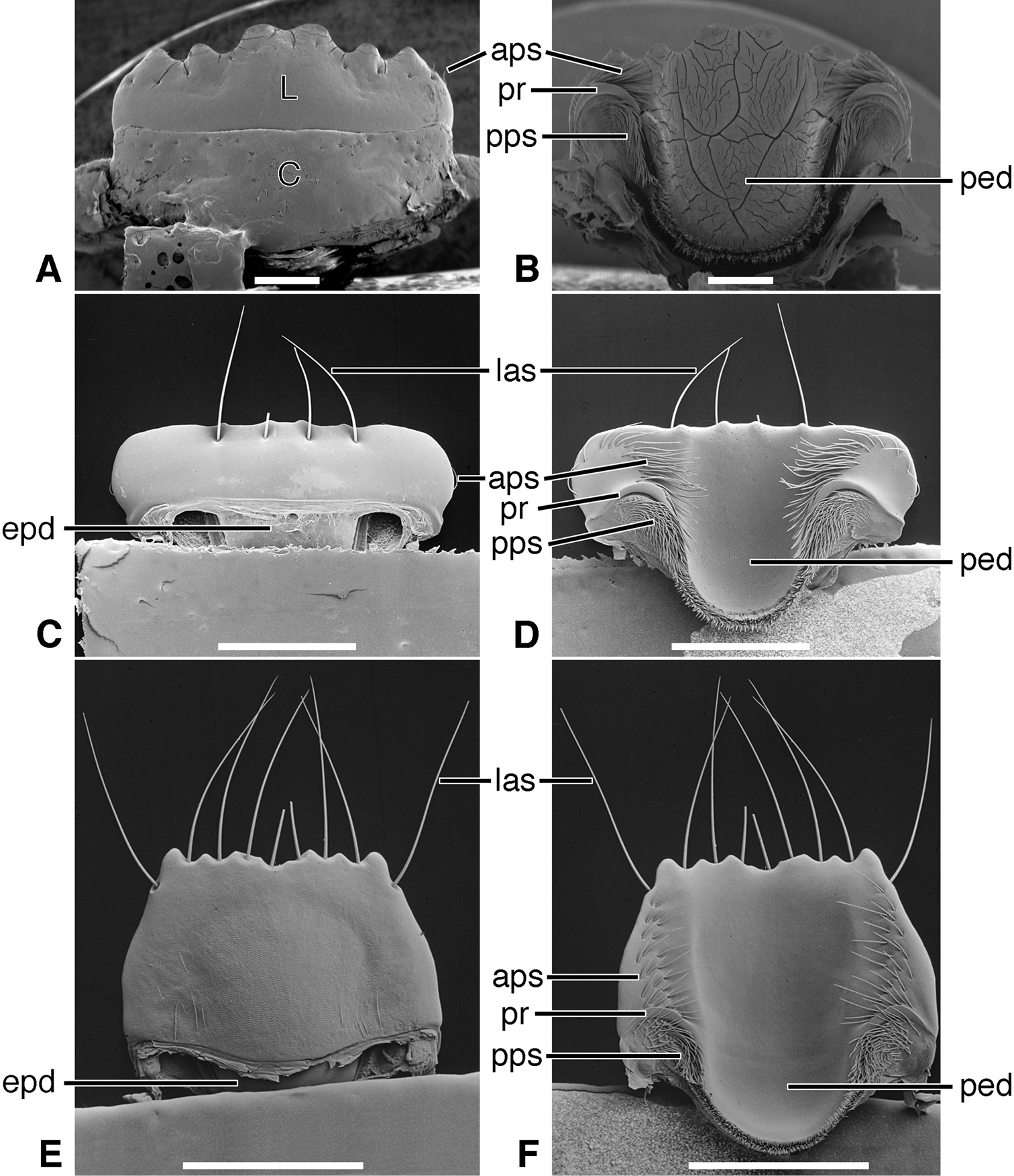
Labrum (Figures 9A, C, E; 13A, C, E; and 17A, C, E). Attached immovably to clypeus, although labral-clypeal suture evident. Of various proportions (Length/ Width 0.20-1.02), i.e., distinctly transverse, sub-rectangular, to slightly longer than wide and trapezoid or trianguloid; row of tactile setae either near anterior margin, on dorsal surface (Figure 9A), or on apical margin (Figure 13E). Form of anterior margin various, from subtruncate (Figure 13C) to simply projected medially (Figure 9C), to markedly projected (Figure 17C); projection one or several denticles, or broadly rectangular (Figure 9C). See also Cazier (1954: 306-307, Figures 124-169).
Epipharynx (Figures 9B, D, F; 13B, D, F; and 17B, D, F).Pedium (ped) palatiform, markedly arched, ventral surface concave; posteriorly broadly rounded, without a parapedial projection, margined laterally each side by a thin parapedial ridge (pr); parapedial ridge anteriorly each side curved to lateral margin of labrum as a short lateral arm. Parapedial setae in a row or cluster anterior (aps) to or posterior (pps) to lateral arms of parapedial ridge.
Comments. Cicindelite mandibles are readily distinguishable from those of other geadephagans by a combination of complex and distinctive retinacular structure (essential to the “pre-oral mill” of
Synonymic note. Commonly known as the Omini W.
Classification. This tribe includes three Western Hemisphere genera: the western Nearctic Amblycheila Say and Omus Dejean; and the southern Neotropical Picnochile Motschulsky (female). If the southern Afrotropical monobasic genus Platychile Macleay is placed in this group (e.g.
Exemplar taxa. Amblycheila baroni (Rivers, 1890) (female); Omus californicus (Eschscholtz, 1829) (male); Picnochile fallaciosa (Chevrolat, 1854) (female).
Also examined but not treated in detail were two males of Platychile pallida (Fabricius, 1801).
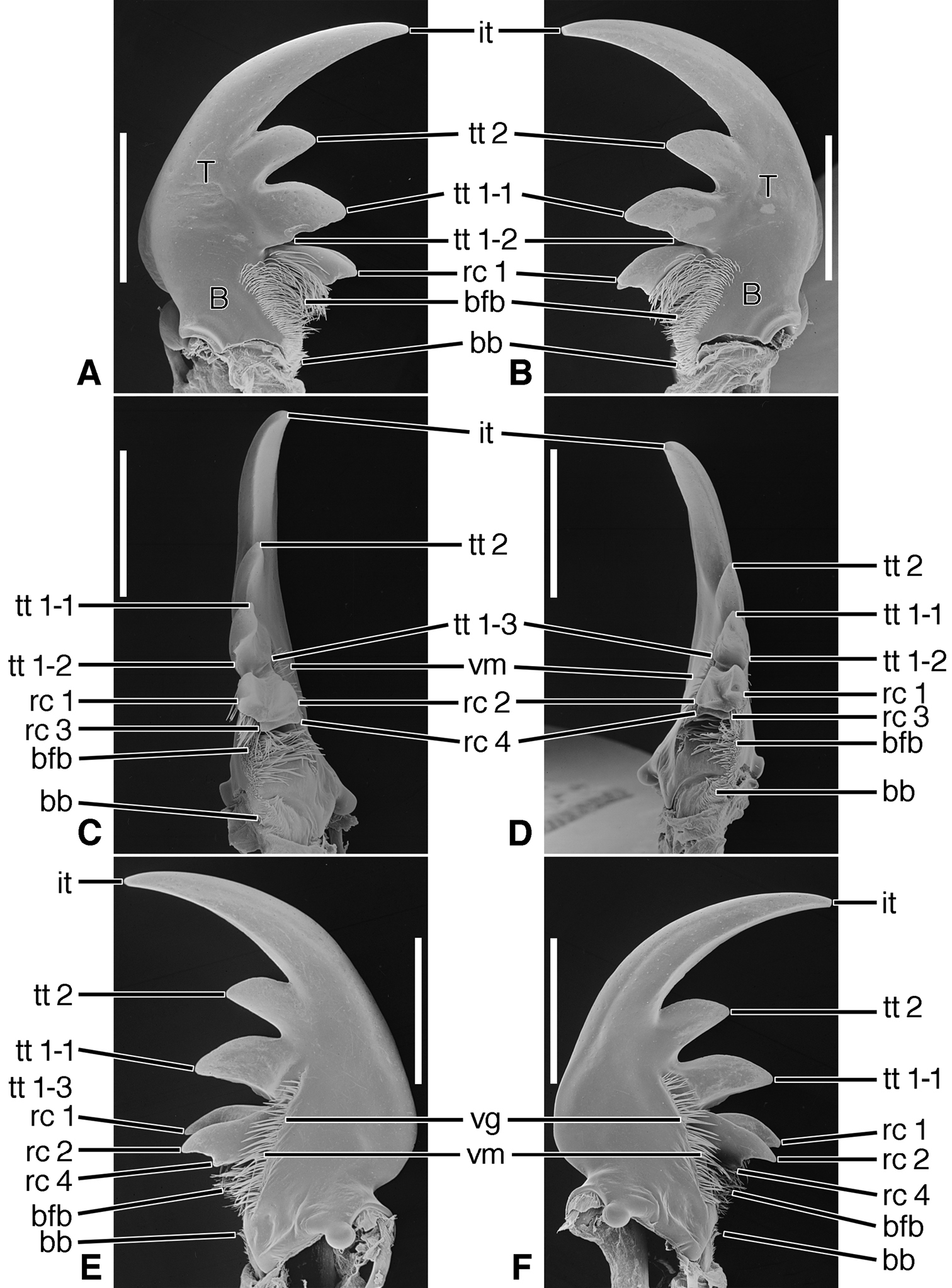
Structural features. Mandibles (Figures 6A-F – 8A-F; Plate 1B).— With mandibular features of Cicindelitae, restricted as follows. Values for ratios BW/TL. TeL/TL, and IT/TL as in Table 3; planar (Figures 6C-D) to moderately curved ventrad (Figures 7C-D). Lateral surface basally with scrobe broad, delimited by a dorsolateral and ventrolateral ridge;scrobe multisetose (Figures 6A-F, ss) or glabrous. Diastema absent. Terebral teeth two, terebral tooth 1 tricuspidate (Figures 6A-F, tt 1-1, tt 1-2, tt 1-3). Retinacular cusps (Figures 6C-D) rc 1 and rc 2 directly opposite one another, also rc 3 and rc 4 directly opposite one another.Ventral groove (Figures 6E-F and 10E-F, vg)moderately long, extended about to middle of terebral tooth 1.
Ratios for features of left mandible of exemplar specimens of Trachypachus, Pelophila, Ceroglossus, and nine genera of Cicindelitae, representing Tribes Amblycheilini, Manticorini, Megacephalini, Collyridini, and Cicindelini.
| Taxon | BW2/TL1 | TeL3/TL | IL4/TL |
|---|---|---|---|
| Trachypachus gibbsii LeConte | 0.47 | 0.53 | 0.12 |
| Pelophila rudis LeConte | 0.32 | 0.76 | 0.10 |
| Ceroglossus chilensis Eschscholtz | 0.24 | 0.54 | 0.12 |
| Amblycheila baroni (Rivers) | 0.27 | 0.77 | 0.46 |
| Omus californicus | 0.27 | 0.59 | 0.57 |
| Picnochile fallaciosa (Chevrolat) | 0.32 | 0.71 | 0.43 |
| Manticora tuberculata (DeGeer) | 0.27 | 0.68 | 0.33 |
| Tetracha carolina (Linnaeus) | 0.30 | 0.70 | 0.34 |
| Pogonostoma chalybeum (Klug) | 0.20 | 0.69 | 0.42 |
| Therates basalis Dejean | 0.19 | 0.66 | 0.37 |
| Oxycheila species | 0.24 | 0.70 | 0.39 |
| Cicindela longilabris Say | 0.17 | 0.68 | 0.45 |
1 Total Length; 2 Basal width; 3 Terebral Length; 4 Incisor Length

SEM photographs of mandibles of Omus californicus Eschscholtz. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.
SEM photographs of mandibles of Picnochile fallaciosa Chevrolat. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.
SEM photographsof labrum and epipharynx of: Amblycheila baroni Rivers (A labrum, dorsal aspect; B epipharynx, ventral aspect); Omus californicus Eschscholtz (C labrum, dorsal aspect; D epipharynx, ventral aspect); Picnochile fallaciosa Chevrolat (E labrum, dorsal aspect; F epipharynx, ventral aspect). Legend: see Table 2. Scale bars: 1.0 mm.

SEM photographs of mandibles of Manticora latipennis Waterhouse. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 5.0 mm.
Labrum (Figures 9A, C, E; Plate 1B). Transverse (L/W 0.20-0.36), in form rectanguloid. Anterior margin distinctly projected medially, otherwise various: projection truncate (Figure 9C) or narrowly notched (Figure 9A); or anterior margin with two paramedial notches, anterior margin of medial projection broadly emarginate (Figure 9E). Single preapical row of 8-10 setae (las).
Epipharynx (Figures 9B, D, F). Row of anterior parapedial setae (aps) extended laterally, parallel to lateral extensions of parapedial ridge (pr).
Comments.
The genus Platychile deserves special comment here. We examined two males of Platychile pallida (Fabricius) using light microscopy. A number of mandibular features (planar, markedly curved; terebra bidentate; retinaculum quadridentate) place the genus among the more basal lineages. On the other hand, the mandibles are so flat as to virtually eliminate the scrobe (and are thus reminiscent of the more derived tiger beetles), and they possess a single cusp terebral tooth 1 instead of the three cusps characteristic of the Western Hemisphere genera Plate 1F). The rectangular labrum (Plate 1F) has an anterior margin with two paramedial dentiform projections, flanked each side by two more short and blunt projections (six, in all) and six setae on the anterior (not apical) surface. The labrum, in fact, in form and setation, is strongly reminiscent of that of the tribe Megacephalini. Further, the body size and form is not unlike that of what could be expected in the megacephaline genus Phaeoxantha. The color pattern is also megacephaline-like, and a careful reading of Pearson and Vogler (2001: 53-57) indicates that such a feature may have importance in tiger beetle evolution. The unusual habitus of Platychile may also derive in part from convergence or mimicry, since these nocturnal beetles show a consistent ecological association with the diurnal Eurymorpha cyanipes (Hope, 1838) (
Classification. This tribe includes the southern Afrotropical genera Manticora Fabricius, 1792 and Mantica Kolbe, 1896.
Exemplar taxon. Manticora tuberculata (DeGeer, 1778) (female).
Other specimens examined: see Appendix.
Structural features. Mandibles (Figures 10A-F). With mandibular features of Cicindelitae, restricted as follows. Values for ratios BW/TL. TeL/TL, and IT/TL as in Table 3. Robust; markedly curved ventrad (Figures 10C-D). Lateral surface basally with scrobe broad, delimited by a dorsolateral and ventrolateral ridge;scrobe multisetose (Figures 10A-F, ss). Diastema absent. Terebral teeth two (some female Manticora) or three (male Manticora, somefemale Manticora, andboth sexes of Mantica), terebral tooth 1 monocuspidate (Figures 10A-F, tt 1); or bicuspidate, tt 1-2 (appearing separate from tt 1-1 in genusMantica. Retinacular cusps (Figures 10C-D) rc 1 and rc 2 directly opposite one another, also rc 3 and rc 4 directly opposite one another.Ventral groove (Figures 10E-F, vg)moderately long, extended about to middle of terebral tooth 1.
The mandibles of Manticora are pronouncedly sexually dimorphic, larger in males and asymmetric in form, the right mandible typically exhibiting greater elongation of the incisor region than the left. In Mantica, sexual dimorphism is slight, and some but not all males show larger left than right mandibles (
Labrum (Figure 13A). Transverse (L/W 0.20-0.36), in form rectanguloid. Anterior margin distinctly crenate, Manticora with six teeth, Mantica with four teeth (
Epipharynx (Figure 13B).Row of anterior parapedial setae (aps) extended laterally, parallel to lateral extensions of parapedial ridge (pr).
Comments. See also Plate 1C. In structure of mandibles and labrum-epipharynx, the Manticorini seems most similar to the Amblycheilini. However, the marked ventrad curvature of the mandibles is suggestive of the more derived megacephalines, cicindelines and collyrines. With the genera Amblycheila and Pogonostoma, members of Manticora share multisetose scrobes.
For details about way of life, classification and relationships of Manticora, see
Classification. This tribe includes more than 100 species, arrayed in eight genera (
Exemplar Taxon. Tetracha carolina (Linnaeus, 1767) (male).
Other specimens examined: see Appendix.
Structural Features. Mandibles (Figures 11A-F).With mandibular features of Cicindelitae, restricted as follows. Values for ratios BW/TL. TeL/TL, and IT/TL as in Table 3. Robust; markedly curved ventrad (Figures 11C-D). Lateral surface basally with scrobe broad, delimited by a dorsolateral and ventrolateral ridge;scrobe glabrous (Figures 11A-F). Diastema absent. Terebral teeth three, terebral tooth 1 monocuspidate (Figures 11A-F, tt 1). Retinaculum with supplementary tooth anteriorly (srt) in Tetracha and Megacephala, very small in Phaeoxantha. Retinacular cusps (Figures 11C-D) rc 1 and rc 2 directly opposite one another, also rc 3 and rc 4 directly opposite one another.Ventral groove (Figures 11E-F, vg)moderately long, extended about to base of terebral tooth 2.
SEM photographs of mandibles of Tetracha carolina Linnaeus. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.

SEM photographs of mandibles of Pogonostoma chalybeum Klug. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.
SEM photographsof labrum and epipharynx of: Manticora latipennis Waterhouse (A labrum, dorsal aspect; B epipharynx, ventral aspect); Tetracha carolina Linnaeus (C labrum, dorsal aspect; D epipharynx, ventral aspect); Pogonostoma chalybeum Klug (E labrum, dorsal aspect; F epipharynx, ventral aspect). Legend: see Table 2. Scale bars: 1.0 mm.
Labrum (Figure 13C). Transverse (L/W 0.28), in form rectanguloid. Anterior margin shallowly crenate, medially, median projection short, broad, emarginate anteriorly). Single preapical row of four setae (las).
Epipharynx (Figure 13D). Row of anterior parapedial setae (aps) extended anteriorly, at about right angle to lateral extensions of parapedial ridge (pr).
Comments. See also Plate 1D, E. The long terebral tooth 3 illustrated for the specimen of Tetracha carolina seems to be characteristic of males of that genus, contrasting markedly with the shorter t3 of the corresponding females. Number of labral setae in Megacephalini ranges from four to seven.
Supertribe Cicindelitae: Tribe CollyridiniClassification. This group is the equivalent of and co-extensive with Walther Horn’s (1908) “Phylum” Alocosternalia. This tribe includes seven genera arranged in two subtribes: Ctenostomatina, and Collyridina (
Exemplar taxon. Pogonostoma chalybeum (Klug, 1835) (male).
Other collyridines examined include representatives of subtribes Ctenostomatina and Collyridina. See appendix.
Structural features. Mandibles (Figures 12A-F). With mandibular features of Cicindelitae, restricted as follows. Values for ratios BW/TL, TeL/TL, and IT/TL as in Table 3. Slender; markedly curved ventrad (Figures 12C-D). Lateral surface basally with scrobe broad, delimited by a dorsolateral and ventrolateral ridge;scrobe multisetose (Figures 12A-F, ss). Diastema (od) present. Terebral teeth various in size and number (see “Variation” below, for details). Retinaculum without or with (Collyris only) supplementary tooth anteriorly (srt). Retinacular complex large, cusps (Figures 12C-D) diagonally arranged, number of cusps various (see below for details)Ventral groove (Figures 12E-F, vg)moderately long, extended about to base of terebral tooth 2.
Labrum (Figure 13E). Elongate (L/W 0.63), in form trapezoidal. Anterior margin shallowly crenate. Single apical row of nine setae (las), each seta inserted in base of crenulation. For details, See “Variation”, below.
Epipharynx (Figure 13F). Pedium (ped) markedly concave. Row of anterior parapedial setae (aps) extended anteriorly, at about right angle to lateral extensions of parapedial ridge (pr). For details, See “Variation”, below.
Variation. Terebral teeth two (each mandible), tt 1 monocuspidate (Figures 12A-F) (Pogonostoma), or only one tooth on each mandible (Collyris); or terebral teeth asymmetric, with two on right mandible and one on left mandible (Ctenostoma), or one on left mandible, two on right mandible (Tricondyla). Retinacular cusps five each mandible (Pogonostoma), or seven on left mandible, five on right mandible (Ctenostoma), or five on left mandible, four on right mandible (Tricondyla and Collyris).
Although the labrum-epipharynx is consistently elongate and dorsally convex, the number of anterior marginal teeth varies from five to eight, some taxa with medial crenulation and an odd number of teeth; some with median notch and an even number of teeth; lateral pair of teeth generally acute, median teeth in form of rounded crenulations; shallow grooves present, in some taxa on labrum and/ or epipharynx, extended posteriad notch separating lateral and medial teeth. Number of labral setae various, from six to 14. Anterior parapedial setae extended almost to anterior margin in Pogonostoma, but less so in other collyridine taxa. It is not clear how medial teeth, notches, and/or setae have evolved from their paired bilateral homologues.
Comments. See Plate 2A for illustrations of the mandibles and labrum of Ctenostoma ichneumoneum Dejean, 1833. Clearly, although the collyridines present a diversity of mouthpart configurations, there is no obvious reason to doubt the use of Pogonostoma as an exemplar for the group, likely to exhibit a more or less plesiotypic structural condition. The ant-like body form of most collyridines may have constrained the head shape and therefore mouthpart structure of these beetles to some extent, but this is merely conjecture on our part.
Supertribe Cicindelitae: Tribe CicindeliniClassification. This tribe includes more than 1500 species, arranged in five subtribes: Theratina; Oxycheilina; Iresina; Prothymina; and Cicindelina (Vogler and Barraclough, 1998).
Exemplar taxa. Therates basalis Dejean (Theratina); Oxycheila species (Oxycheilina); and Cicindela longilabris Say (Cicindelina).
Other cicindelines examined: see Appendix for names;plus numerous species of Nearctic Cicindela, principallyfor form, and for number of labral setae.
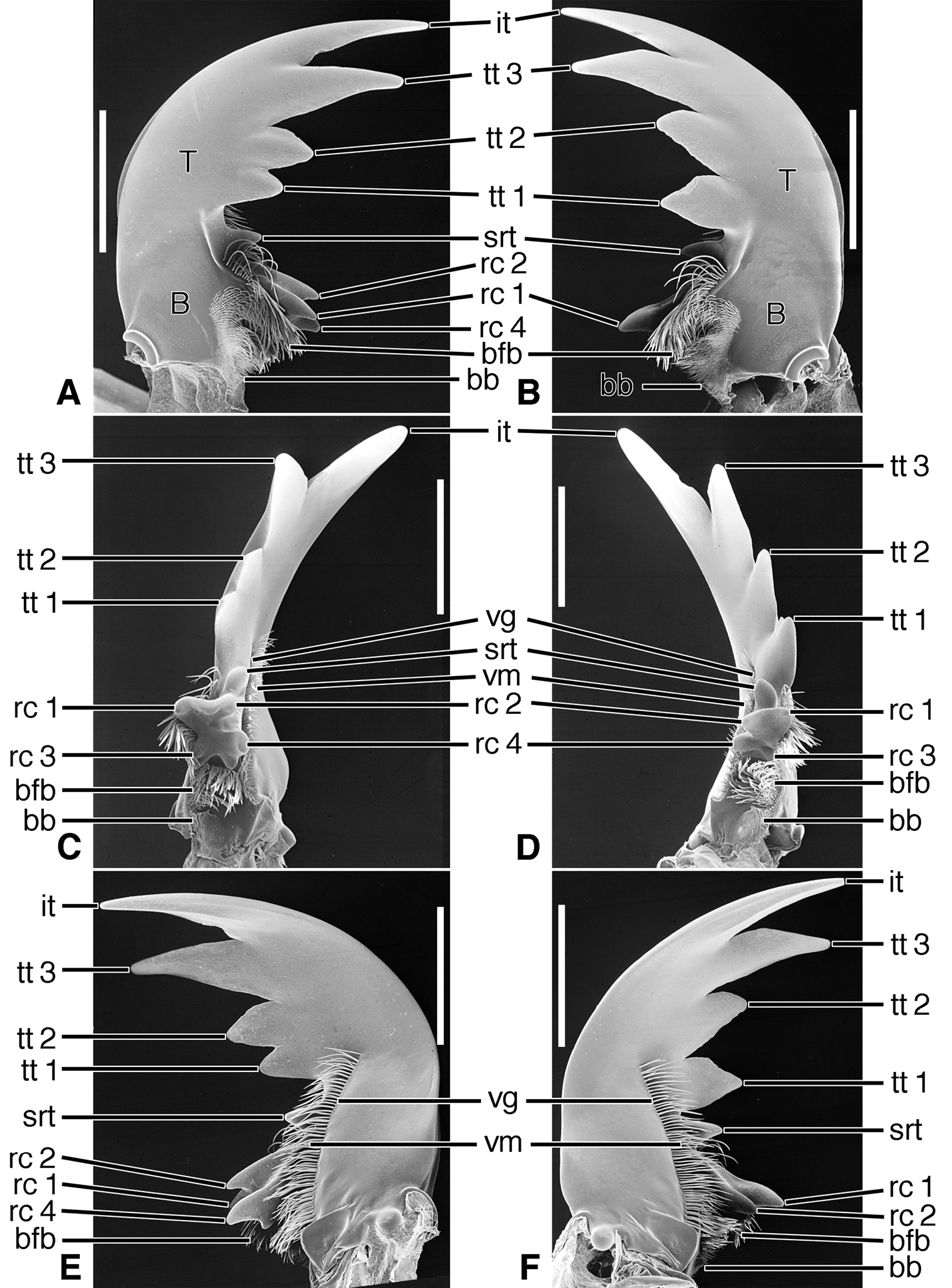
Structural features. Mandibles (Figures 14A-F- 16A-F; Plate 2F). With mandibular features of Cicindelitae, restricted as follows. Values for ratios BW/TL, TeL/TL, and IT/TL as in Table 3. Slender; markedly curved ventrad (Figures 14C-D). Lateral surface basally without scrobe, or scrobe very narrow, confined to lateroventral surface (Dromica), and without setae. Diastema present (Figures 14A-F and 16A-F, od) or absent (Figures 15A-F). Terebral teeth various in size and number: one, (Figures 14A-F, tt 2) to four (Figures 15A-F, tt 1 – tt 4), with a maximum of five, observed in Oxygonia gloriola Bates, 1872. Retinaculum without or with (Therates, Figures 14A-F) supplementary tooth anteriorly (srt). Retinacular complex large, cusps diagonally arranged, number of cusps four on each mandible (rc 1- rc 4).Ventral groove (Figures 15E-F, vg)moderately long, extended about to base of terebral tooth 2, or shorter, extended only to base of retinacular complex (Figures 16E-F).

SEM photographs of mandibles of Therates basalis Dejean. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.

SEM photographs of mandibles of Oxycheila species.Eschscholtz. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.

SEM photographs of mandibles of Cicindela longilabris Say. A, C, E left mandible, dorsal, occlusal, ventral aspects, respectively; B, D, F right mandible, dorsal, occlusal, ventral aspects, respectively. Legend: see Table 2. Scale bars = 1.0 mm.
Labrum (Figures 17A, C, E). Markedly varied. Illustrations as follows. Form transverse (L/W 0.41, Figure 17E), as long as wide (L/W 1.00, Figure 17A), or slightly longer than wide (L/W 1.02, Figure 17C). Trapezoidal to trianguloid. Anterior margin nearly smooth or distinctly crenate. Labral setae (las, plls) four to 12, on dorsal or apical surface, inserted at base of crenations.

SEM photographsof labrum and epipharynx of: Therates basalis Dejean (A labrum, dorsal aspect; B epipharynx, ventral aspect); Oxycheila species (C labrum, dorsal aspect; D epipharynx, ventral aspect); Cicindela longilabris Say (E labrum, dorsal aspect; F epipharynx, ventral aspect). Legend: see Table 2. Scale bars: 1.0 mm.
Epipharynx (Figures 17B, D, F). Anterior parapedial setae (aps) in row parallel to lateral arm of parapedial ridge (Figure 17D), extended in short row anterior and obliquely to lateral arm of parapedial ridge (Figure 17E) or only very few setae laterally (Figure 17B).
Variation. For illustrations of mandibles and labra of additional cicindeline taxa, see Plate 2C-E. The mandibles of three exemplar taxa described above, each representing a different cicindeline subtribe, differ strikingly from one another, seeming to indicate an appreciable level of divergence in this tribe. Within the subtribe Cicindelina,
In their remarkable study of geographical variation in Cicindela dorsalis Say, 1817,
Intensive investigation of mandibular length (
Our three exemplar taxa differ markedly from one another in form and setation of the labrum. We note that the insertion of the labral setae on the dorsal surface of the labrum in Cicindela longilabris Say, 1824 (Figure 17E) is a relatively basal condition.
The epipharynges of the three exemplar taxa are basically similar to one another, but differ in the anterior parapedial setation. The small number of such setae exhibited by Therates basalis Dejean, 1826 (Figure 17B) is the most derived, and is similar to that of the collyridine, Ctenostoma metallicum Laporte de Castelnau, 1834 (not illustrated).
EvolutionThe following hypothesis of mandibular and labral-epipharyngeal evolution is illustrated in the reconstructed phylogeny (Figure 18) based on
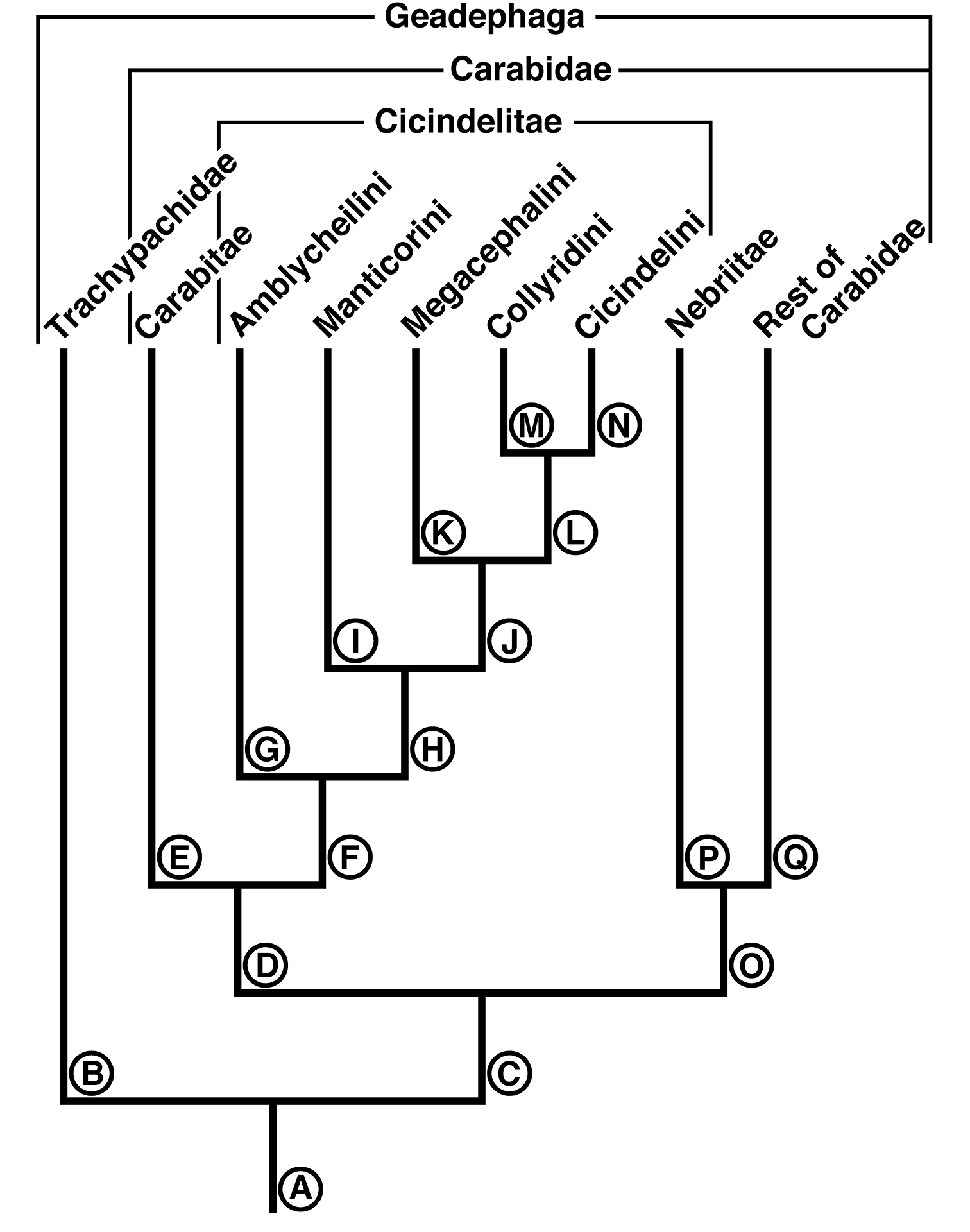
Diagram of a reconstructed phylogeny of the cicindelite tribes, related higher carabid taxa, and Trachypachidae. Reference points on this diagram are the lineages indicated by the capital letters A to Q.

Digital images of head capsule, labrum, and mandibles dorso-frontal aspect, of: A Ceroglossus chilensis (Eschscholtz); B Amblycheila baroni (Rivers); C Manticora mygaloides Thomson; D Megacephala regalis Boheman; E Phaeoxantha tremolerasi (W. Horn); F Platychile pallida (Fabricius). Scale bars: A, B, E, F = 5 mm; C, D = 10 mm.

Digital images of head capsule, labrum, and mandibles dorso-frontal aspect, of: A Ctenostoma ichneumoneumDejean; B Therates erinnysBates; C Cheiloxya binotata Laporte de Castelnau; D Pseudoxycheila species?; E Dromica junodi Péringuey; F Cicindela longilabris Say. Scale bars: A = 3 mm; B, E =4 mm; C, D, F = 5 mm.
Lineage A represents the common ancestor of the Geadephaga, in which we assume the mandibles possessed a distinct terebra and retinaculum, of unknown form, but probably Pelophila-like. The labrum-epipharynx was likely movably articulated with the head capsule.
Lineage B represents the evolution of the Trachypachidae, with mandible (Figures 2A-F) and labral-epipharynx (Figures 5A-B) features as follows: mandibles, short, broad, slightly curved ventrally, terebra relatively short, with short incisor and short terebral teeth, a very short basal face, and the basal brush (bb) serving as the basal face setae. The retinaculum includes two long retinacular ridges (rr, srr), and is bicuspidate, teeth (art, prt) short.
The labrum-epipharynx is movably articulated with the head capsule: labrum transverse, rectanguloid, anterior margin subtruncate, dorsal surface with a row of numerous setae (14, more or less); epipharynx with short, trianguloid pedium, with few anterior and posterior parapedial setae, and a small parapedial projection. Diet yet to be determined: probably partially fluid and partially particulate matter.
Lineage C The stem of the Carabidae. Evolution of the mandibles includes only simplification of the retinaculum to a single broad ridge and possibly lengthening of the terebra. The diet was probably generalized, including both solid and fluid food (the “mixed feeders” of
Lineage D Evolution of carabite-cicindelite mandibles as the diet became fluid, only, the food principally soft-bodied invertebrates. Includes development of a somewhat enlarged and densely setose basal face for retention of the fluid component of prey tissues for extra-oral digestion, and the labrum-epipharynx becoming immovably attached to the head capsule.
Lineage O Evolution of mandibles (Figures 3A-F) of the nebriite- “rest of Carabidae” Lineage includes development of a scrobal seta (ss), and narrowing of the single ridged retinaculum (rr). (Not followed further here, but see
Lineage E Evolution of mandibles of Carabitae (Figures 4A-F) includes marked lengthening of the basal face (bf) shortening of the retinaculum and its diagonal orientation (rr). The ventral groove (Figures 4E-F, vg) became markedly lengthened to nearly the base of the incisor tooth, and the ventral microtrichia (vm) became markedly lengthened. The anterior margin of the labrum (Figure 5E) became markedly emarginate.
Lineage F Evolution of mandibles of Cicindelitae includes hypertrophy of the incisor tooth, development of a second terebral tooth, hypertrophy of the terebral teeth, and the scrobes becoming multisetose. But the most striking mandibular development is seen in the retinaculum, which becomes markedly enlarged and quadricuspidate – an important element of the preoral mill.
The labrum becomes lengthened, its anterior margin sinuously arched and sparsely setose, the number of anterior setae reduced to 8-10. The epipharynx is extensively modified, becoming palatiform, dorsally arched, and widened posteriorly, with loss of the parapedial projection. Evans and Forsythe (1985: 116) describe the mode of feeding, unique among fluid-feeding carabids, in which prey is held, punctured and sheared by the incisor and terebral teeth, then passed posteriorly by the maxillary lacinia to the crushing teeth of the retinaculum. Within the preoral mill (bordered dorsally by the epipharynx, ventrally by the setose labium and laterally by the setose retinacular region of the mandibles), a food bolus is rotated posterio-dorsally and anterio-ventrally, bathed in midgut enzymes. Partially digested fluid is drawn through the mouth by a powerful pharyngeal pump until all but fragments of cuticle have been ingested, at which point the bolus is ejected.
Nodes and stems G-N map the evolution of the mandibles and labrum-epipharynx in the Cicindelitae.
Lineage G Evolution of the amblycheiline mandibles involves principally enlargement of the terebral tooth 1 (Figures 6C-D, tt 1-1), which becomes tricuspidate (tt 1-1 – tt 1-3). Within the Amblycheilini, the scrobal setae are retained in Amblycheila, but lost in the other genera (Figures 7A-B and 8A-B), and the slight down-curvature of the terebra is reduced (Figures 6C-D). The anterior margin of the labrum is variously modified, the median extension truncate (Figure 9A), or narrowly notched (Figure 9C), or broadly emarginate (Figure 9E).
Node H Evolution of the remaining lineages (H-N) of the Cicindelitae. The basic mandibular and labral-epipharyngeal features are those of the Amblycheilini, outlined above, except terebral tooth 1 consists only of a single large cusp.
Lineage I Evolution of the manticorine mandibles (Lineage I) involves marked ventral curvature of the terebra (Figures 10C-D). The mandibles of Mantica and female Manticora are otherwise very amblycheiline-like, but those of males are remarkably hypertrophied, with an especially elongate incisor tooth. Mandibular sexual dimorphism developed in numerous lineages throughout the phylogenetic history of tiger beetles, but became most pronounced in the lineage leading to Manticora, in which the males developed tremendously elongate mandibles, especially in the incisor region. Mandibular dimorphism likely developed in concert with prolonged copulation, and mate-guarding and the fitting of the male mandible to the female metathoracic copulatory sulcus (
Lineage J Evolution of the Megacephalini and Collyridini + Cicindelini (Lineages K- N). Mandibles of this lineage develop a more or less extensive diastema between terebral tooth 1 and anterior margin of the retinaculum (Figures 11A-F – 13A-F, od), and retain the scrobal setae in a few genera (e.g. Megacephala, Pogonostoma). The epipharynx undergoes slight differentiation with the row of the anterior parapedial setae extended anteriorly (Figures 13D, F; 17F).
Lineage K Evolution of the mandibles of Megacephalini includes development of a supplementary retinacular tooth (Figures 11A-F, srt), and hence a longer retinaculum. Within this lineage, the terebral teeth differentiate in number (one to three) among taxa, and between sexes of the same taxon (cf.
Lineage L The common ancestor of the tribes Collyridini and Cicindelini. Compared to those of its adelphotaxon (the Megacephalini), the mandibles (Figures 12A-F – 16A-F) become slender and labrum-epipharynx is extensively enlarged. The labral anterior margin becomes crenate, with the setae inserted apically rather than dorsally (Figures 13E and 17A, C). Since the arboreal members of the group evolved from ground-dwelling ancestors, tiger beetles serve as an example of the taxon pulse hypothesis (
Lineage M The Collyridini. The mandibular retinaculum becomes more complex, with one or more additional cusps (Figures 12B, D, F). Within the tribe, the mandibles become markedly varied. We illustrate only what must be a relatively basal lineage– the genus Pogonostoma, with only two terebral teeth on both left and right mandible, and only one additional retinacular cusp on only one (right) mandible. Scrobal setae are lost in all collyridine lineages examined with the exception of Pogonostoma. Terebral teeth are reduced in number in, or lost from, the genera Ctenostoma Klug, Tricondyla Latreille, and Collyris Fabricius.
Lineage N The Cicindelini. (Observations based principally on exemplar specimens representing the genus Therates Latreille, Oxycheila Dejean, and Cicindela Linnaeus, arranged in evolutionary order of appearance, according to the reconstructed phylogeny of Vogler and Barraclough). Also examined: Oxygonia gloriola Bates.
Within this tribe, and as in the Collyridini, the mandibles become markedly varied, as shown in Figures 14A-F, 15A-F and 16A-F. The scrobe, narrow in the more basal lineages, is lost from the more highly derived Cicindela. The ancestral number of terebral teeth was probably three as in Cicindela (Figures 16A-F), becoming four as in Oxycheila (Figures 15A-F, tt 1 - tt 4), and reducing to one, as in Therates (Figures 14A-F, tt 2). The supplementary retinacular tooth re-evolves in Therates (Figures 14A-F, srt). The ventral groove, normally quite long (Figures 15E-F, vg), becomes shortened in Cicindela longilabris (Figures 16E-F, vg), and in other species of this genus (
Building on increasingly sophisticated phylogenetic hypotheses for the Geadephaga, and the Cicindelitae (e.g.
Further research would benefit from the examination of additional taxa, both within the Cicindelitae and among other relatively basal lineages of the Carabidae, in order to better address questions such as: is the Carabitae indeed the sister group of the Cicindelitae, and is Platychile more closely related to the amblycheilines or to the megacephalines? Incorporation of mouthpart features as character systems in phylogenetic analyses is recommended, as are further studies of the biomechanics of tiger beetle feeding and the use of mandibles during mating and mate-guarding, since the evolutionary changes we hypothesize are difficult to interpret without an appreciation of the functional consequences of changes in mouthpart configuration. In this light, further studies are also needed to adequately characterize sexual dimorphism, asymmetry, and chirality among tiger beetle mouthparts.
Through the good offices of Terry Thormin and Matthias Buck, The Royal Alberta Museum, Edmonton, Alberta, loaned to us the specimens used to launch our study of tiger beetle mouthparts. We are grateful to these colleagues for arranging this loan. We thank George Braybrook and De-ann Rollings (Department of Earth and Atmospheric Sciences, University of Alberta) for capturing the digital SEM images of our exemplar specimens. For a prompt response to our last-minute loan request for specimens of Platychile pallida, we thank David H. Kavanaugh (California Academy of Sciences, San Francisco, California); and for providing names for several Neotropical cicindeline species used in the present study, we thank Ronald L. Huber (Bloomington, Minnesota). Finally, we thank James K. Liebherr (Department of Entomology, Cornell University, Ithaca, New York) and David L. Pearson (Department of Zoology, Arizona State University, Tempe, Arizona) for insightful comments on earlier drafts of this paper. Publication was made possible by Natural Sciences and Engineering Research Council of Canada, NSERC Research Grant OGP 1399, to GEB.
List of names of tiger beetle taxa examined with Wild M3 and M5 stereomicroscopes for mandibular and/or labral-epipharyngeal features.
incertae sedis
Platychile pallida (Fabricius, 1801) (2 males)
Tribe Manticorini
Manticora mygaloides Thomson, 1859 (2 males, 2 females)
Manticora latipennis Waterhouse, 1837 (1 male, 1 female)
Manticora tuberculata (DeGeer, 1778) (3 males, 2 females)
Tribe Megacephalini
Megacephala regalis Boheman, 1848 (1 male)
Phaeoxantha tremolerasi (W. Horn, 1909) (1 female)
Phaeoxantha wimmeri Mandel, 1958(1 male, 1 female)
Tetracha annuligera Lucas, 1857 (1 male, 1 female)
Tetracha virginica (Linnaeus, 1767)(1 male, 1 female)
Tribe Collyridini
Ctenostoma metallicum (Laporte de Castelnau, 1834) (1 male)
Ctenostoma unifasciatum Dejean, 1831 (1 male, 1 female)
Ctenostoma ichneumoneum Dejean, 1826 (1 female)
Collyris dohrnii Chaudoir. 1860(1 female)
Tricondyla species? (3 males)
Tribe Cicindelini
Therates erinnys Bates, 1874 (1 male, 1 female)
Pseudoxycheila ceratoma Chaudoir, 1865 (1 male)
Cheiloxya binotata Laporte de Castelnau, 1833 (1 male)
Dromica junodi Péringuey, 1892 (2 males, 1 female)
Oxygonia gloriola Bates, 1872 (= Oxygonia simplicipennis Horn) (1 male)