





(C) 2011 Timothy T. Work. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Biodiversity monitoring is increasingly being bolstered with high resolution data derived from remote sensing such as LIDAR (Light Detection and Ranging). We derived a series of topographical variables, including slope, azimuth, ground curvature and flow accumulation from LIDAR images and compared these to captures of female carabids in pitfall traps in Eastern boreal mixedwood forests. We developed a series of species-specific logistic models predicting the proportion of females for eight dominant species, including Agonum retractum, Calathus ingratus, Platynus decentis, Pterostichus adstrictus, Pterostichus coracinus, Pterostichus pensylvanicus, Sphaeroderus nitidicollis and Synuchus impunctatus. We used these models to test three hypotheses related to how the modest topography in boreal forests could influence the availability of microhabitats and possibly potential sites for oviposition and larval development. In general, topographic features such as north facing slopes and high flow accumulation were important predictors of the proportion of females. Models derived from larger scale topography, such as hillsides or small watersheds on the order of ¼-1 ha were better predictors of the proportion of females than were models derived from finer scale topography such as hummocks and small depressions. We conclude that topography likely influences the distribution of carabids based on hydrological mechanisms rather than factors related to temperature. We further suggest based on the scale of responses that these hydrological mechanisms may be linked to the attenuation of past disturbances by wildfire and the propensity of unburned forest patches and fire skips.
boreal carabids, topography, LIDAR, remote sensing, fire skips
Inherent in many biodiversity monitoring programs is a belief that there are landscape units or other easily measurable habitat characteristics that can be consistently related to particular species or assemblages. This perspective is at the heart of coarse filter approaches to biodiversity conservation whereby stand structures and habitat features are often used as surrogates for individual or multiple species (
While links between habitat units and organisms will require validation across numerous taxa, many recent efforts have focused on the relation between carabid beetles with landscape units or stand structures (
Increasingly, larger-scale classifications such as forest inventory data, ecological landtype phases [ELTPs], and stand maps are being complemented with much higher resolution data on stand structure and underlying topography. One such example is Light Detection and Ranging [LIDAR] which provides detailed digital terrain models [DTMs] and fine scale information related to the topography of surfaces (
Surface topography could affect carabids in many ways. South-facing slopes in northern climates are often preferentially selected as oviposition sites by a variety of insects. These habitats are often attributed some sort of developmental advantage for immature stages of insects, where south facing oviposition sites may be warmer (Andreson et al. 2001;
Here we have related changes in the proportion of females for eight dominant carabid species in boreal forests to topographic variables derived from LIDAR images. We concentrated our analysis on four primary topographic elements; slope, azimuth, curvature and elevation, and one secondary element; flow accumulation derived from a high precision, LIDAR digital elevation map in order to examine three hypotheses related to microhabitat preference of female carabids.
We propose three working hypotheses which relate surface topography to the proportion of females with the underlying assumption that this proportion reflects preferred oviposition sites and initial larval habitats (
All carabids were collected using pitfall traps at the SAFE research site in the Lac Duparquet Research and Teaching Forest in northwestern Québec (
LIDAR data was acquired in full-leaf conditions on 14-16 August 2003 using an Optech ALTM 2050 sensor flown at 1 000 m above ground level. The sensor recorded the first and last returns of each laser pulse emitted at 50 000 Hz. The last returns were classified into ground and non-ground categories by the data provider using the Terrascan application from Terrasolid Ltd (Helsinki, Finland). The last returns classified as ground were interpolated and gridded at resolutions of 4, 8, 16 and 32 m to create multi-scale DTMs. Topographic variables were derived from these LIDAR DTMs and included local slope, azimuth, curvature and flow accumulation using ArcGIS 9.3 (ESRI, Redlands CA). Slopes were defined as a percentage where 100% would be equivalent to 45°. Azimuths were translated to ‘northing’ where values range from 0-180° with 0° reflecting N and 180° reflecting S, thus eliminating the E-W axis from azimuth. Curvature is defined from second-order derivatives from a 4th-order polynomial surface based on elevation. Positive values represent hummocks where as negative values represent depressions. Flow accumulation (FAC) corresponds to the number of grid cells that are connected to a target cell through monotonic increases in elevation. We recognize that this measure cannot be interpreted in the strict sense of fluids passing over impermeable surfaces, but nevertheless we feel that this provides some insight into runoff following heavy precipitation events and soil hydrology. Each of these variables was derived by placing a 3 × 3 cell grid centered on pitfall trap locations. We derived each variable at four spatial scales corresponding to the DTM’s grid cell sizes of 4, 8, 16 and 32 m. Thus topographic variables reflect increasingly larger surface areas or neighborhoods [144, 576, 2034, 9026 m2]. By changing the scale of inference on topographic variables we feel that we capture a range of perspectives reflecting ‘microsites’ to ‘seepage’.
Relating carabids to topographic parametersThe proportion of females captured for each species were related to the five topographic parameters at each scale using logistic regression. Prior to this analysis we evaluated a series of logistic-mixed models, where individual forest cohorts were treated as random effects in the model. In each case, these random variations among cohorts were extremely small, thus we simplified our approach using simple logistic models.
For each logistic model, the five topographic variables were centered to a mean of 0. We used AIC to select models that best reflected changes in the proportion of females to four scales of LIDAR derived topographic variables. Once an optimal scale was selected, the full model containing all five topographic variables was compared to four reduced models using AICc. These reduced models were chosen to reflect hypotheses related to female responses to temperature, moisture and elevation. The reduced models were 1) slope and azimuth, 2) slope, azimuth and elevation, 3) curvature and FAC, and 4) curvature, FAC and elevation. For each model, we corrected for model dispersion using the sum of squared pearson`s residuals divided by residual degrees of freedom. Models were constructed using the glm function in R 2.11.1 (
The mean and standard deviation of slope and curvature decreased with increasing scales (Table 1). Thus at larger spatial scales topography became flatter and more planar. In contrast, aspect, elevation and FAC were largely consistent across scales. Together these variables typify a boreal landscape with modest changes in slopes which are distributed in all directions and where small, ‘fine-grained’ topographic features [such as small hummocks and depressions] are the norm (Table 2). Differences in elevation across all scales were equally modest, ranging from 38-43 m.
Means and standard deviations of topographic parameters derived at four spatial scales from LIDAR.
| Parameter | 4m | 8m | 16m | 32m | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| Azimuth | 83.14 | 59.724 | 78.45 | 62.7 | 84.63 | 60.746 | 61.46 | 58.978 |
| Slope | 9.87 | 6.399 | 7.39 | 3.47 | 4.11 | 2.605 | 2.18 | 2.006 |
| Curvature | -0.07 | 0.786 | 0.02 | 0.267 | 0 | 0.041 | 0 | 0.021 |
| FAC | 12.46 | 34.021 | 10.1 | 11.079 | 17.71 | 42.16 | 15.49 | 19.46 |
| Elevation | 249.7 | 11.443 | 249.76 | 11.41 | 248.94 | 12.275 | 251.33 | 15.569 |
Absolute range of topographic parameters derived at four spatial scales from LIDAR.
| Parameter | 4m | 8m | 16m | 32m |
|---|---|---|---|---|
| Azimuth | 173.60 | 176.86 | 167.37 | 174.16 |
| Slope | 27.39 | 17.70 | 9.39 | 6.71 |
| Curvature | 3.94 | 1.25 | 0.20 | 0.14 |
| FAC | 216.00 | 49.00 | 228.00 | 79.00 |
| Elevation | 38.21 | 38.52 | 38.85 | 43.17 |
With the exception of Agonum retractum, the proportion of females was related to topographic variables derived at 16 and 32 m scales (Table 3). With the exception of Platynus decentis, the proportion of females observed was best explained by reduced models which had consistently lower AICc values than the initial full model.
AIC values for full logistic models based on azimuth, slope, curvature, flow accumulation and elevation.
| Species | 4m | 8m | 16m | 32m |
|---|---|---|---|---|
| Agonum retractum | 148.651 | 151.444 | 151.012 | 155.601 |
| Calathus ingratus | 72.711 | 73.287 | 70.220 | 73.112 |
| Platynus decentis | 126.694 | 142.549 | 138.993 | 124.737 |
| Pterostichus adstrictus | 178.912 | 178.274 | 181.027 | 175.792 |
| Pterostichus coracinus | 130.115 | 130.577 | 122.559 | 129.707 |
| Pterostichus pensylvanicus | 225.792 | 224.805 | 223.546 | 219.138 |
| Sphaeroderus nitidicollis | 133.667 | 134.729 | 131.134 | 137.884 |
| Synuchus impunctatus | 160.347 | 160.566 | 159.233 | 159.532 |
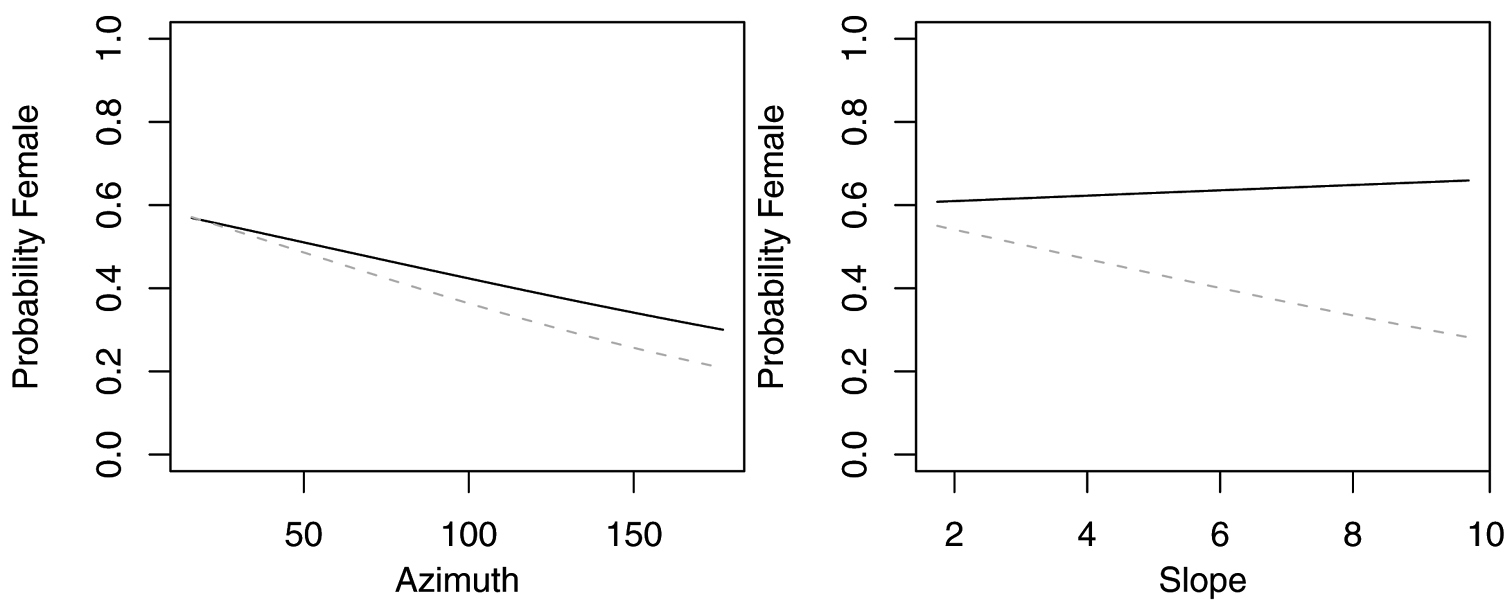
The reduced model with only azimuth and slope was the best model for Agonum retractum (Figure 1) and Calathus ingratus (Figure 2), although in both cases azimuth was the only significant predictor (Table 4). In both these cases, the proportion of females increased on more northern azimuths. The odds of finding Agonum retractum and Calathus ingratus females on northern azimuths were 1.97 (95% CI=1.144 to 3.398) and 3.083 (95% CI= 1.139 to 8.340) times greater, respectively than on southern azimuths. Reduced models for Pterostichus adstrictus and Pterostichus coracinus also included azimuth and slope but were expanded to include elevation based on AICc comparisons (Table 5). Elevation was a significant predictor for both species while proportion of Platynus adstrictus females was larger on northern azimuths (Figure 3) and proportion of Pterostichus coracinus females increased with slope (Figure 4). The odds of finding Platynus adstrictus females were 2.249 (95% CI= 1.034 to 4.892) times greater on northern azimuths than southern azimuths and 2.228 (95% CI= 1.034 to 4.798) times greater at its maximum elevation than its minimum elevation. The odds of finding Pterostichus coracinus females were 6.394 (95% CI= 1.956 to 20.897) times greater on its maximum slopes than its minimum slopes and 10.539 (95% CI= 3.290 to 33.759) times greater at its minimum elevation than its maximum elevation.
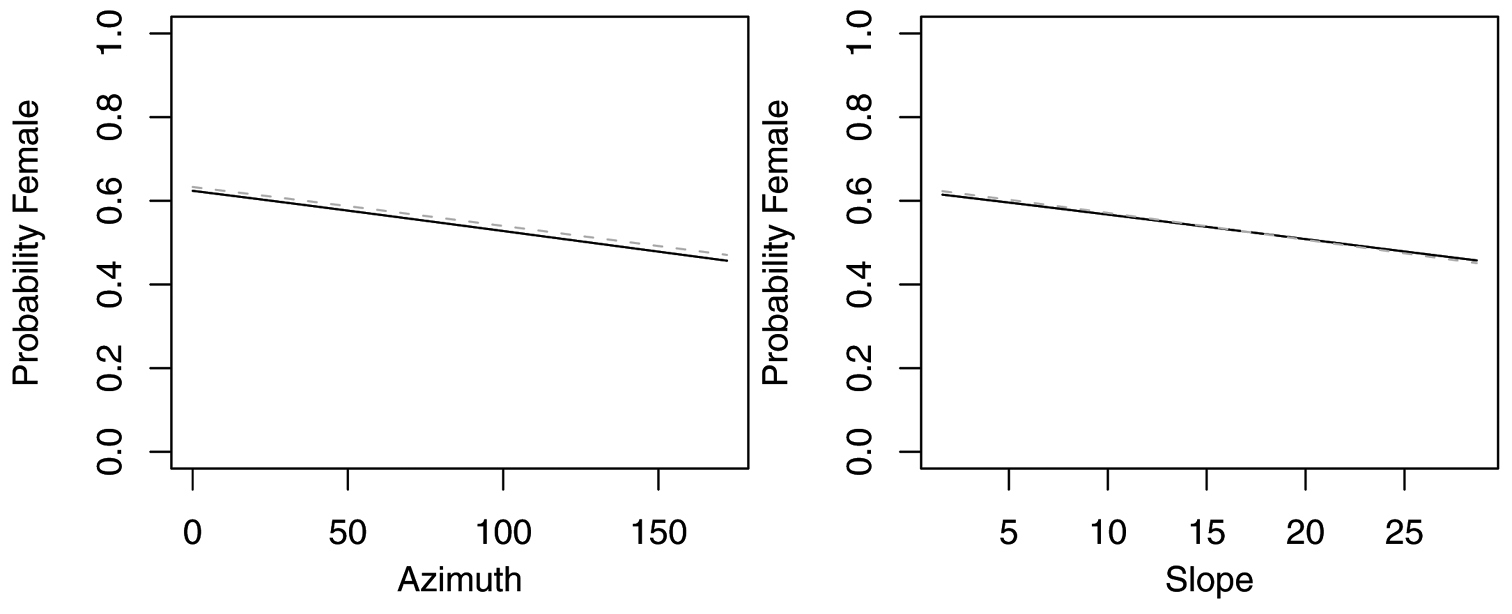
Probability of collecting female Agonum retractum with changes in azimuth and slope when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only azimuth, slope and intercept.
Probability of collecting female Calathus ingratus with changes in azimuth and slope when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only azimuth, slope and intercept.
Reduced logistic models for Agonum retractum [4m scale] and Calathus ingratus [16m scale] 1.
| Estimate | Std. Error | z value | Pr(>|z|) | |
|---|---|---|---|---|
| Agonum retractum | ||||
| (Intercept) | 0.5059 | 0.09270 | 5.45800 | 0.00000 |
| Azimuth | -0.0039 | 0.00161 | -2.44700 | 0.01440 |
| Slope | -0.0236 | 0.01562 | -1.51200 | 0.13050 |
| Calthus ingratus | ||||
| (Intercept) | 0.3904 | 0.14514 | 2.69000 | 0.00714 |
| Azimuth | -0.0070 | 0.00315 | -2.21700 | 0.02662 |
| Slope | 0.0276 | 0.06661 | 0.41500 | 0.67834 |
Table 5. Reduced logistic models for Pterostichus adstrictus [32m scale] and Pterostichus coracinus [16m scale] 1.
| Estimate | Std. Error | z value | Pr(>|z|) | |
|---|---|---|---|---|
| Pterostichus adstrictus | ||||
| (Intercept) | 0.1793 | 0.10270 | 1.74600 | 0.08080 |
| Azimuth | -0.0047 | 0.00229 | -2.04400 | 0.04090 |
| Slope | -0.1107 | 0.07034 | -1.57300 | 0.11560 |
| Elevation | 0.0186 | 0.00907 | 2.04600 | 0.04080 |
| Pterostichus coracinus | ||||
| (Intercept) | -0.3303 | 0.12082 | -2.73400 | 0.00626 |
| Azimuth | 0.0035 | 0.00239 | 1.48000 | 0.13881 |
| Slope | 0.2766 | 0.09006 | 3.07100 | 0.00214 |
| Elevation | -0.0546 | 0.01376 | -3.96500 | 0.00007 |
Probability of collecting female Pterostichus adstrictus with changes in azimuth, slope and elevation when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only azimuth, slope, elevation and intercept. Regression slopes for elevation appear near zero as total range of elevation is relatively small.
Probability of collecting female Pterostichus coracinus with changes in azimuth, slope and elevation when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only azimuth, slope, elevation and intercept. Regression slopes for elevation appear near zero as total range of elevation is relatively small.
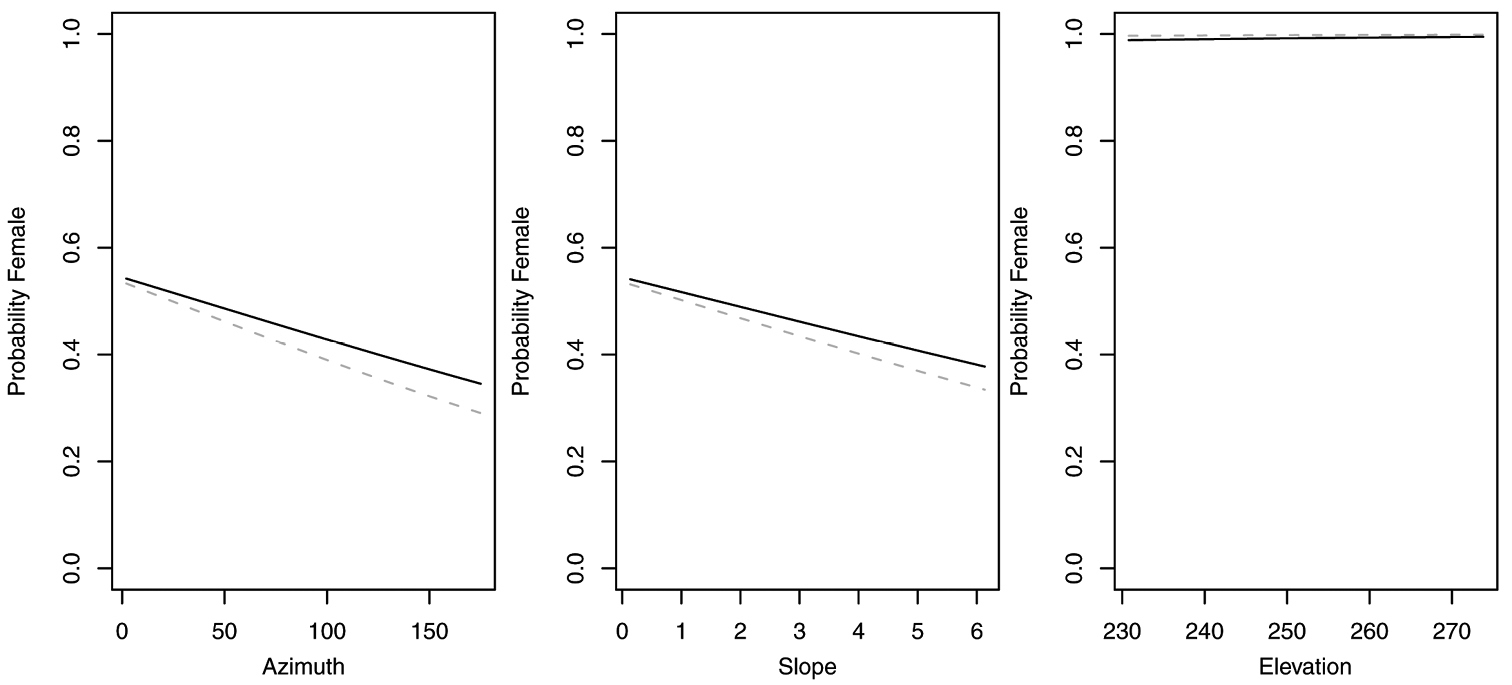
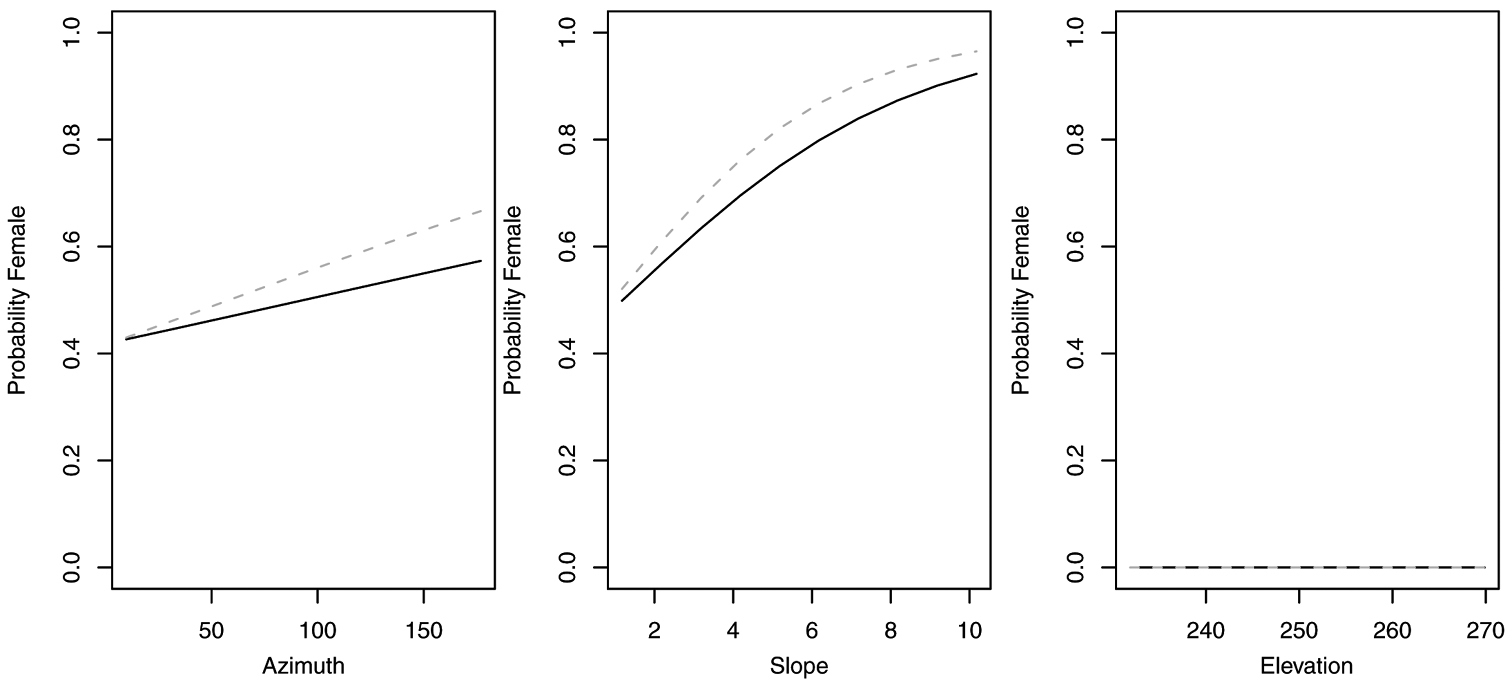
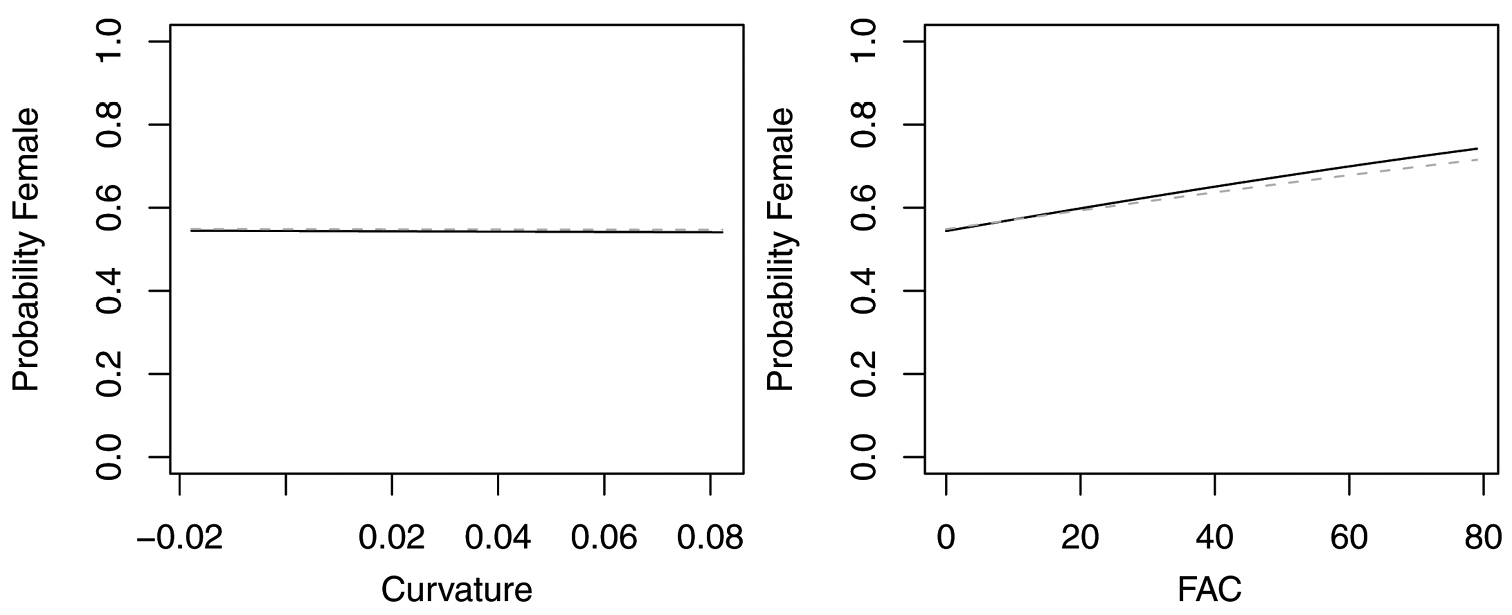
In contrast, reduced models for Synuchus impunctatus, Sphaeroderus nitidicollis and Pterostichus pensylvanicus included only curvature and FAC, rather than slope, aspect or elevation (Table 6). In the reduced model for Synuchus impunctatus, neither curvature nor FAC were significant (Figure 5). Proportion of females for Pterostichus pensylvanicus increased with increasing FAC (Figure 6). The odds of finding female Pterostichus pensylvanicus were 2.411 (95% CI= 1.126 to 5.164) times greater in areas at its maximum FAC than areas with low FAC. Proportion of females of Sphaeroderus nitidicollis increased with both increasing FAC and curvature (Figure 7). The odds of finding female Sphaeroderus nitidicollis were 3.693 (95% CI =1.190 to 11.465) greater at its maximum FAC than on its minimum FAC and 3.521 (95% CI=1.493 to 8.300) times greater in areas of its maximum curvature than minimum curvature. Platynus decentis was the only species where the full topographical model best explained changes in the proportion of females based on model comparisons (Table 7), however, like Sphaeroderus nitidicollis only FAC and curvature were significant predictors of changes in the proportion of females (Figure 8). However, unlike Sphaeroderus nitidicollis, Platynus decentis was negatively associated with both FAC and curvature. The odds of finding Platynus decentis were 22.000 (95% CI = 3.233 to 149.714) times greater on sites with minimum FAC than on sites with maximum FAC. The odds of finding Platynus decentis were 8.499225 (95% CI = 1.736651 to 41.59547) greater at areas with minimum curvature than areas with maximum curvature.
Reduced logistic models for Pterostichus pensylvanicus [32 m scale], Sphaeroderus nitidicollis and Synuchus impunctatus [both at 16m scale]1.
| Estimate | Std. Error | z value | Pr(>|z|) | |
| Pterostichus pensylvanicus | ||||
| (Intercept) | 0.1767 | 0.07114 | 2.48400 | 0.01300 |
| Curvature | -0.1532 | 1.93062 | -0.07900 | 0.93680 |
| FAC | 0.0111 | 0.00492 | 2.26500 | 0.02350 |
| Sphaeroderus nitidicollis | ||||
| (Intercept) | -0.5619 | 0.10347 | -5.43100 | 0.00000 |
| Curvature | 6.3393 | 2.87685 | 2.20400 | 0.02760 |
| FAC | 0.0057 | 0.00254 | 2.26100 | 0.02380 |
| Synuchus impunctatus | ||||
| (Intercept) | 0.6328 | 0.12900 | 4.90600 | 0.00000 |
| Curvature | 3.9788 | 3.81754 | 1.04200 | 0.29700 |
| FAC | 0.0001 | 0.00353 | 0.03000 | 0.97600 |
1P-values were corrected with 1.487152, 0.8426052 and 1.456111, respectively as dispersion parameters
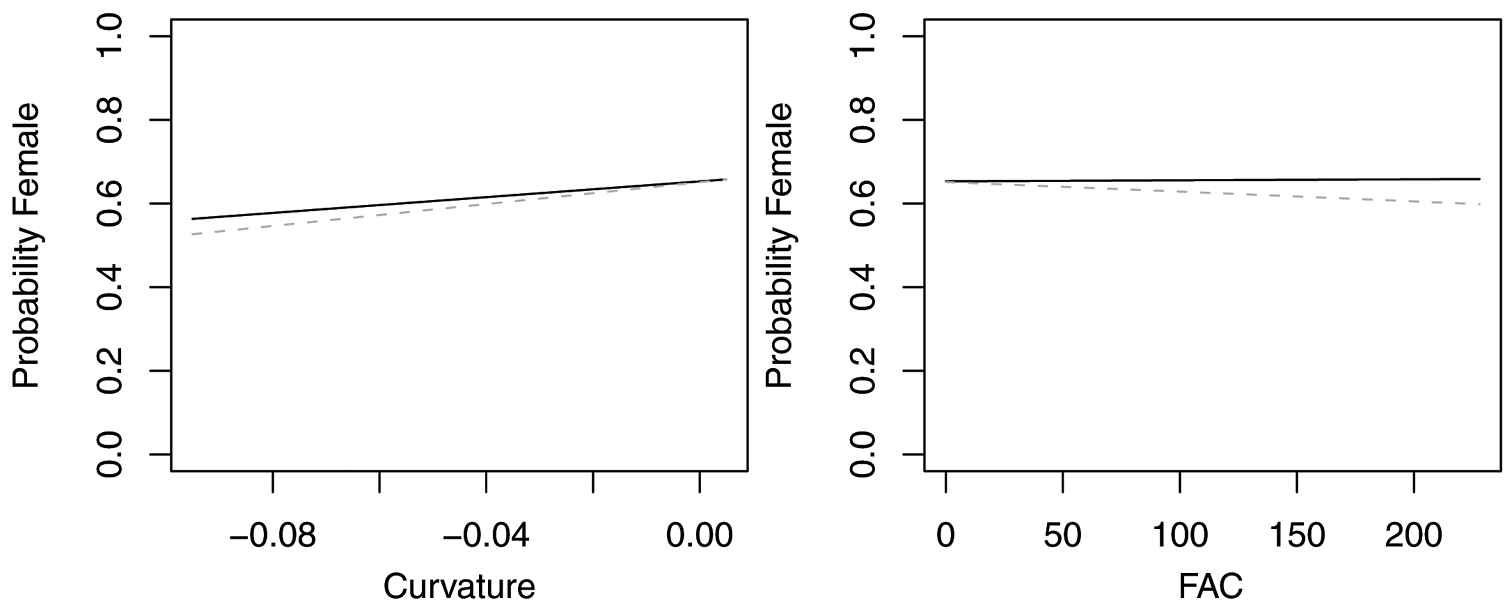
Figure 5. Probability of collecting female Synuchus impunctatus with changes in azimuth and slope when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only curvature, FAC and intercept.
Probability of collecting female Pterostichus pensylvanicus with changes in azimuth and slope when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only curvature, FAC and intercept.

Probability of collecting female Sphaeroderus nitidicollis with changes in azimuth and slope when additional variables are held constant at their respective means. Dashed lines represent model parameters derived from full logistic model. Solid lines represent model parameters of reduced logistic model including only curvature, FAC and intercept.

Probability of collecting female Platynus decentis with changes in azimuth and slope when additional variables are held constant at their respective means. Both dashed and solid lines represent model parameters derived from full logistic model. Regression slopes for elevation appear near zero as total range of elevation is relatively small.
Reduced logistic models for Platynus decentis [32 m scale]1.
| Estimate | Std. Error | z value | Pr(>|z|) | |
|---|---|---|---|---|
| (Intercept) | 0.6225 | 0.13125 | 4.74300 | 0.00000 |
| Azimuth | -0.0062 | 0.00343 | -1.79600 | 0.07252 |
| Slope | 0.0455 | 0.18603 | 0.24500 | 0.80672 |
| Curvature | -14.8768 | 5.63250 | -2.64100 | 0.00826 |
| FAC | -0.0391 | 0.01239 | -3.15900 | 0.00158 |
| Elevation | 0.0287 | 0.02251 | 1.27300 | 0.20289 |
1P-values were corrected with 1.101403 as a dispersion parameter
Our data suggest that females for all but one species are responding to topographic features on the order of 1/4-1 ha rather than highly localized microsites [i.e.<1/20 ha]. While it is tempting to further relate body size as a surrogate for dispersal ability under a hypothesis that ‘large species operate at larger scales’, our data does not support this hypothesis. While Agonum retractum, the smallest species used in our analysis, responded to the smallest scale of topographic variables, all other species were associated with larger scales [16-32 m]. To rigorously test this hypothesis, more species should be included along with a larger range of spatial scales.
Neither does our data correspond directly with either of our hypotheses related to development time or small scale hydrology. Species that oviposit preferentially with slope and aspect in order to speed or synchronize larval development are often phytophagous and strongly linked to plant phenology (
In our study site, ¼-1 ha corresponded to topographic features on the order of small hillsides or catchments rather than hummocks or depressions related to uprooted trees, large rocks or buried logs. In the eastern boreal forest, modest differences in topography are nested within a landscape where heterogeneity is maintained by large-scale, stand replacing fire. This fire regime results in a skewed distribution where there are numerous small fires [on the order of <1-10 ha] and relatively few very large fires (
The only species associated with small scale topography, Agonum retractum was positively associated with northern azimuths. Providing scale specific interpretations of azimuth, as compared to other topographic variables, is difficult as azimuth varies little over different spatial scales. Thus physical explanations related to northern azimuths such as retention of snow and thus higher humidity could also be applied to smaller scales such as the northern sides of hummocks.
Alternatively, the smaller scale responses of Agonum retractum may be a methodological artifact related to elevation errors in the LIDAR image. Elevational errors are caused in part by variations of the vertical accuracy of LIDAR under forest canopies. These errors are influenced by ground return density and by the ground/non-ground classification success. Derivatives such as slope, aspect and FAC are quite sensitive to small scale elevational errors and may be relatively noisy at high resolution. This noise would in turn greatly weaken statistical relationships to observed insect abundance. At larger scales, minute LIDAR errors cancel out leading to increased robustness of derivatives.
The fact that we are able to detect changes in the proportion of females in response to very small changes in topography demonstrates the potential of high-resolution remote sensing techniques such as LIDAR, which is currently the only 3D remote sensing instrument capable of creating accurate and high resolution topographical maps, a capacity that opens up numerous research paths. In the recent past, LIDAR-based habitats studies have focused on flora, or larger animals such as birds or mammals. We have additionally demonstrated that LIDAR DTMs can be useful for studying the habitat preferences of insects at different scales, even for a single insect family in a landscape with very mild topography.
ConclusionsWe believe that advances in remote sensing such as LIDAR will likely continue to complement biodiversity assessments rather than replace actual biological inventories as a stand-alone surrogate. While it is initially seductive to mine high resolution remote sensing data for correlations with assemblages, this seems more of a preliminary exercise in calibration of the technique than an end goal. The utility of LIDAR will be borne out when the force of this data is realized through mechanistic hypotheses related to habitat requirements of plants and animals. We have shown that topographic features such as north facing slopes and sites with high FAC are more likely to harbor female carabids in eastern boreal landscapes where topography is modest, although the current imprecision of these models clearly limits predictive applications of these models. We further demonstrate that 7 of the 8 species analyzed respond to topography consistent with the scale of fire skips and suggest that topography may be affecting the distribution of females by maintaining more mesic habitats and attenuating wildfire. This in itself is a preliminary hypothesis which assumes a strong link between distribution of females, oviposition choice and larval survivorship. However we hope that using LIDAR to identify preferred microhabitats for females will lead us to a better understanding of preferred habitats of larvae which we feel has been largely overlooked in discussions of carabids as bioindicators or in discussions of refining coarse filter approaches to conservation.
Kit O’Connor collected and identified all specimens used in this analysis. This paper was inspired through presentations and conversations during the Fetschrift presented in honor of Ross and Joyce Bell, whose contributions to carabidology has touched many of our dear colleagues. We thank the Canadian Foundation for Innovation for funding the 2003 airborne LIDAR data collection.