





(C) 2011 Ryuki Murao. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Lasioglossum (Lasioglossum) subopacum (Smith) is recorded from the Korean Peninsula for the first time. Lasioglossum (Lasioglossum) okinawa Ebmer et Maeta from Japan is ranked to a subspecies of Lasioglossum (Lasioglossum) subopacum judging from the characteristics of the male. The male of Lasioglossum (Lasioglossum) subopacum okinawa is described for the first time. Some bionomical notes of both subspecies are presented.
Hymenoptera, Halictidae, Lasioglossum, Asia, taxonomy
The halictine bee subgenus Lasioglossum s. str. Curtis, 1833 (Halictidae: Halictinae) is morphologically characterized by the second submarginal crossvein of female fore wing as strong as the first, and the female inner hind tibial spur serrate or pectinate with five or more teeth. This subgenus is mainly known from the Holarctic Region with 111 species recorded in the Palaearctic Region. Two of them, Lasioglossum (Lasioglossum) subopacum (Smith, 1853) and Lasioglossum (Lasioglossum) okinawa Ebmer et Maeta, 1999 are known to occur in Asia: the former from eastern to southeastern Asia (
This study is based on the specimens deposited in the following institutions, which are referred to in the text by the following abbreviations: ELKU, Entomological Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka, Japan; EBSU, Prof. Emeritus Yasuo Maeta’s collection, deposited in the Division of Environmental Biology, Faculty of Life and Environmental Science, Shimane University, Matsue, Japan; BPBM, Maa’s collection borrowed from the Bernice P. Bishop Museum, Honolulu, Hawaii, USA; MNHAH, the late Dr. Shoichi F. Sakagami’s collection, deposited in the Museum of Nature and Human Activities, Hyogo, Sanda, Japan; and without acronym, my private collection, now deposited in the ELKU.
Terminology and style used in the description follow
Comparative material examined. Lasioglossum (Lasioglossum) occidens (Smith, 1873): 1♂, Rifu–cho, Miyagi Pref., Honshu, Japan, 14. VIII. 1979 (K. Goukon, MNHAH, illustrated in Fig. 12); 1♂, Kusasenri, Choyo–son, Aso–gun, Kumamoto Pref., Kyushu, Japan, 11. IX. 2004 (T. Sugimoto, illustrated in Fig. 16); 1♂, Mt. Ten–zan, Kyuragi–machi, Saga Pref., Kyushu, Japan, 13. VIII. 2004 (T. Sugimoto, illustrated in Fig. 17). Lasioglossum (Lasioglossum) sakishima Ebmer et Maeta, 1999: 1♂, Yonehara, Iriomote–jima, Okinawa Pref., Ryukyus, Japan, 18. VI. 1972 (O. Tadauchi, ELKU, illustrated in Figs 13, 18); 1♂, Ohtomi, Iriomote–jima, Okinawa Pref., Ryukyus, Japan, 23. V. 2003 (T. Mita, illustrated in Fig. 19).
Taxonomyhttp://species-id.net/wiki/Lasioglossum_subopacum_subopacum
Figs 11, 20, 22This species is divided into two subspecies, one of which is newly relegated to a subspecies of Lasioglossum subopacum as treated below. The nominotypical subspecies is separated from the ssp. okinawa by only the female characteristic that propodeum, and T1 basally with dense and thick yellowish tomentose as in Figs 20, 22. In male, both subspecies cannot clearly separate. This subspecies is separated from the other Korean Lasioglossum s. str. species in having the combination of following characters: the mesoscutum reflexed upward in both sexes and densely transversely rows on medio-anterior margin in female; the female propodeum and T1 with dense yellowish tomentose; the shape of hair tufts on male S6 and the gonostylus as in Fig. 11; and the male genitalia without ventral retrorse.
China (north and southeastern areas), Taiwan, Vietnam, Philippines, and Korean Peninsula (south).
Female: March to December. Male: May to October.
In Korea, this species visited the following six species of flowering plants. Apiaceae: Angelica miqueliana. Asteraceae: Aster yomena; Sonchus arvensis. Brassicaceae: Brassica sp. Caprifoliaceae: Lonicera sp. Rosaceae: Crataegus sp.
[SOUTH KOREA] 2♀1♂, Cheju Is., 17. X. 2005 (O. Tadauchi and R. Murao), 22. X. 2005 (O. Tadauchi); 1♀, Myeong–do–am, 400–600m, Cheju city, Cheju Is., 15. IX. 1998 (O. Tadauchi, ELKU); 2♀, Pijarim Forests, Pukcheju–gun, Cheju Is., 24. IV. 1997 (O. Tadauchi and J. C. Paik, ELKU); 1♂, Pijarim, Pukcheju–gun, Cheju Is., 15. IX. 1998 (J. C. Paik, ELKU); 1♂, Kwanumsa, 500m, Cheju city, Cheju Is., 14. IX. 1998 (O. Tadauchi, ELKU); 8♀, KwangNung, Pocheon-shi, Gyeonggi–do, 19. V. 1992 (O. Tadauchi, ELKU).
[CHINA] Hunan Prov.: 1♀, Yuanling, 7. V. 1939 (T. C. Maa, BPBM); 1♀ Changteh, Yangshan, 11. X. 1938 (T. C. Maa, BPBM). Fujian Prov.: 1♀, Changting city, 3. VI. 1940 (T. C. Maa, BPBM); 12♀, Chungan, Bohea Hill, 11. VIII. 1939 (T. C. Maa, BPBM), 25. IX. 1939 (T. C. Maa, BPBM), 30. IX. 1939 (T. C. Maa, BPBM), 7. X. 1939 (T. C. Maa, BPBM), 24. IV. 1940 (T. C. Maa, BPBM), 6. V. 1940 (T. C. Maa, BPBM), 11. VIII. 1943 (T. C. Maa, BPBM); 1♀, Kienyang, Nwangkeng, 30. VII. 1943 (T. C. Maa, BPBM); 1♀, Kienyang, Liutun, 6. VII. 1942 (T. C. Maa, BPBM); 3♀ and 1♂, Shaowu city, 26. V. 1943 (T. C. Maa, BPBM), 11. IX. 1943 (T. C. Maa, BPBM), 8. XII. 1941 (T. C. Maa, BPBM), 23. XII. 1941 (T. C. Maa, BPBM, illustrated in Figs 20, 22); 1♀, Shaowu, 9. VII. 1942 (T. C. Maa, BPBM); 2♀, Shaowu, Shuipeichieh, 7. VII. 1941 (T. C. Maa, BPBM), 20. VII. 1941 (T. C. Maa, BPBM); 7♀, Shaowu, ShuiPeiKai, 16. III. 1942 (T. C. Maa, BPBM), 26. III. 1942 (T. C. Maa, BPBM), 13. V. 1943 (T. C. Maa, BPBM), 1. VI. 1943 (T. C. Maa, BPBM), V. 1945 (T. C. Maa, BPBM), 1♀, Shaowu, Tachuland, 25. IV. 1943 (T. C. Maa, BPBM); 2♀, Yungan, 19. X. 1940 (T. C. Maa, BPBM), 22. IV. 1941 (T. C. Maa, BPBM).
http://species-id.net/wiki/Lasioglossum_subopacum_okinawa
Figs 1– 10, 14, 15, 21, 23This subspecies is separated from the ssp. subopacum by the female propodeum and T1 basally with sparse and thin whitish tomentose as in Figs 21, 23. In Japan, it is closely similar to Lasioglossum (Lasioglossum) occidens (Smith, 1873) and Lasioglossum (Lasioglossum) sakishima Ebmer et Maeta, 1999. However, it is separated from the former by the mesoscutum reflexed upward in both sexes and densely transversely rows on medio-anterior margin in female, the T1 basally with whitish tomentose tufts in both sexes (Figs 6, 23), the shape of hair tufts on male S6 (Fig. 15), and the gonostylus narrowly rounded apically as in Fig 10; from the latter by the basal elevation of male labrum broadly rounded (Fig. 14), the sculpture on female mesoscutum as stated above, the shape of both hair tufts on male S6 and gonostylus. In contrast, in Lasioglossum occidens, mesoscutum flat in both sexes and densely coarsely punctate on medio-anterior margin in female, T1 without tomentose hair tufts in both sexes as in Fig. 16, hair tufts on male S6 as in Fig. 17, and gonostylus broadly rounded as in Fig. 12; in Lasioglossum sakishima, basal elevation of male labrum small and rounded as in Fig. 18, female mesoscutum reticulate–punctate on anterior margin, shape of hair tufts on male S6 as in Fig. 19, and gonostylus truncate apically as in Fig. 13.
Body length 7.0–8.7mm, wing length 6.0–7.1mm (n=5).
Color. Body black except on the following parts: mandible apical half reddish brown; tegula blackish brown translucent; tibial spur yellow; posterior margin of metasomal terga narrowly brown translucent. Wings nearly transparent; veins and pterostigma blackish brown.
Pilosity. Mostly whitish; pale brown on mesosoma dorsally; mesoscutellum and metanotum mixed with blackish brown hairs. Head with sparse short fine branched hairs, and mixed with moderately dense tomentose on lower paraocular area. Hairs on mesosoma finely branched except on the following parts: dorsal, lateral surface, and around lateral lobe of pronotum with dense tomentose. T1 (Fig. 6) basally with a pair of tomentose tufts, however sometimes disappear. Disc on T2–5 with moderately dense short and simple hairs. Basal hair bands on metasomal terga present on T2–4 or T2–3. Apical fimbriae on metasomal terga absent. Acarinarium absent. S6 with well–formed, distinctive hair tufts as in Fig. 15.
Structure. Head nearly as long as wide; head length/width ratio 1.0–1.01 (n= 5). Vertex flat medially. Distance between lateral ocelli nearly as long as that between lateral ocellus and compound eye. Frons and paraocular area with reticulate–punctate, dimly shiny. Supraclypeal area slightly convex in lateral view, dimly shiny, with reticulate–punctate; IS with distinct tessellation. Clypeus 1.5× distance between lower rim of antennal socket and upper margin of clypeus; nearly flat, with moderately dense PP; IS nearly smooth. Basal area of labrum 2× as wide as long; basal elevation weakly developed, broadly rounded; distal process absent; labral fimbriae acutely pointed at apex. Mandible edentate. Hypostomal carina moderately developed; its anterior angle obtuse. Postgena slightly depressed, with distinct lineoration. Scape length 0.5–0.6mm (n= 5), F2 2.2× F1.
Pronotal dorsolateral angle acute, moderately projecting; pronotal lateral ridge incomplete, interrupted by oblique lateral sulcus; lower portion of lateral ridge inconspicuous, narrowly rounded. Mesoscutum (Fig. 3) with oily–dull luster, dense PP excluding anteriorly, IS smooth; its anterior margin weakly reflexed upwards, with reticulate–punctate. Mesoscutellum marginally and longitudinally with dense PP. Metanotum and mesepisternum coarsely rugulose. Propodeum coarsely rugulose; propodeal dorsum (Fig. 5) 0.7× mesoscutellum, and nearly as long as metanotum; shield marginally with lateral carina that not reaching to apical margin on dorsal surface. Basitibial plate of hind leg carinate marginally. Inner hind tibial spur without distinct teeth.
Metasomal terga with oily–dull luster. T1 with weak lineolation over entire surface, medially and apically with dense PP. T2–3 with dense PP over entire surface; IS weakly lineolate over entire surface. T4–5 similar to IS of T2–3. S7–8 (Fig. 7): S7 with short and slender median process; S8 without median process. Male genitalia as in Figs 8–10. Gonobase ventral arm ring–shaped, and not connected to each other at apical ends: bottom nearly flat. Gonocoxite smooth. Gonostylus simple and flat, butter knife–like apically. Ventral retrorse lobe absent.

Male of Lasioglossum (Lasioglossum) subopacum okinawa Ebmer et Maeta 1 lateral habitus 2 head in frontal view 3 mesoscutum 4 mesosoma in lateral view 5 propodeal dorsum 6 first metasomal tergum.
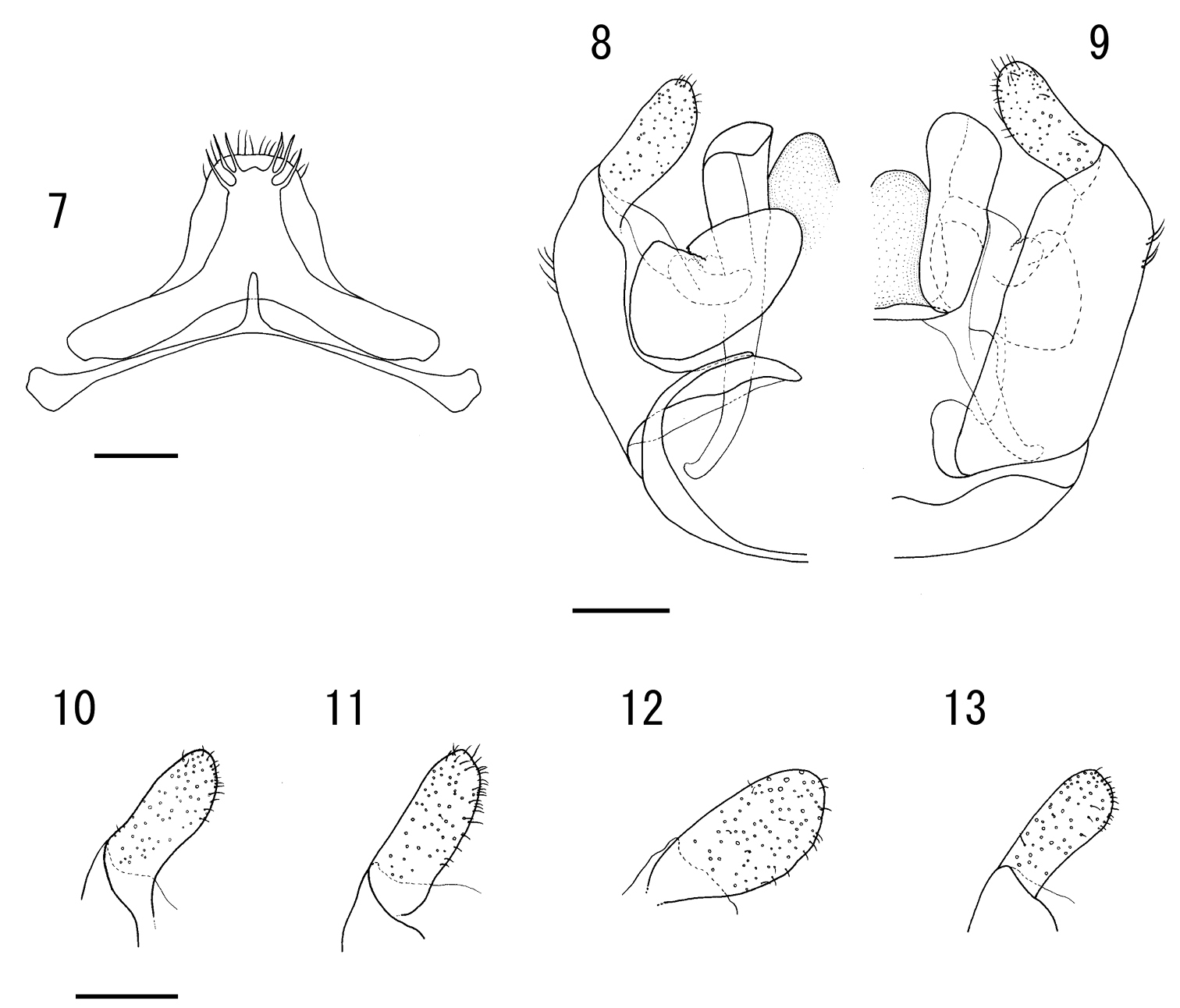
7-10 Male of Lasioglossum (Lasioglossum) subopacum okinawa Emer et Maeta 7 seventh and eighth metasomal sterna 8 genitalia in ventral view 9 genitalia in dorsal view 10–13 gonostylus of genitalia in ventral view 11 Male of Lasioglossum (Lasioglossum) subopacum subopacum (Smith) 12 Male of Lasioglossum (Lasioglossum) occidens (Smith) 13 Male of Lasioglossum (Lasioglossum) sakishima Ebmer et Maeta Scale bars: 0.2mm.
Male clypeus black over the entire surface, or with a small yellow spot on lower margin.
Japan (central Ryukyus: Amami–ôshima, Okinawa–jima, Kume–jima).
Female: April to November. Male: May to October.
This species visited the following six species of flowering plants. Apiaceae: Foeniculum vulgare. Asteraceae: Bidens pilosa var. radiata; Solidago altissima. Euphorbiaceae: Mallotus japonicus. Polygonaceae: Fallopia japonica var. japonica; Persicaria longiseta.
So far as the author’s survey in both Amami–ôshima and Okinawa–jima, this species was mainly collected from around subtropical forest at mountain areas, but in Kume–jima from open land such as cultivated area.
Holotype: ♀, Mt. Yonaha–dake, Okinawa–jima, Okinawa Pref., Japan, 7. VII. 1998 (Y. Maeta, EBSU). [JAPAN] [Ryukyus] Kagoshima Pref.: 1♀16♂, Mt. Yuwan–dake, Uken–son, Amami–ôshima, 14. X. 2004 (R. Murao, 1♂ illustrated in Fig. 1, 1♂ in Fig. 4, 1♂ in Figs 7–10, 1♂ in Fig. 15); 1♂, Mt. Yuwan–dake, alt.500m, Amami–ôshima, 17. VII. 1963 (C. M. Yoshimoto, ELKU); 2♂, Mt. Yuwan–dake, 24. VII. 1963 (Y. Hirashima, ELKU), 29. VII. 1963 (Y. Hirashima, ELKU); 8♀2♂, Shinmura, Amami–ôshima, 23. VII. 1954 (S. Ueda, ELKU; S. Miyamoto and Y. Hirashima, ELKU), 29. VII. 1954 (S. Ueda, ELKU); 1♀, Yuwan, Amami–ôshima, 31. VII. 1963 (Y. Hirashima, ELKU); 3♀, Santarou–toge, Sumiyou–son, Amami–ôshima, 15. X. 2004 (R. Murao, 1♀ illustrated in Figs 21, 23); 1♀, Yakkachi, Sumiyou–son, Amami–ôshima, 19. VII. 1933 (T. Esaki and K. Yasumatsu, ELKU). Okinawa Pref.: 3♀ (paratypes), same data as the holotype; 1♀1♂, 60–180m, Izumi, Motobu, Okinawa–jima, 22. V. 1982 (S. Ikudome, ELKU, 1♂ illustrated in Figs 2, 3, 5, 6); 1♂, Mt. Yonaha–dake, Kunigami–son, Okinawa–jima, 14. VI. 2002 (Y. Maeta, EBSU); 14♀, Hedo, Okinawa–jima, 5. IV. 1979 (K. Ohara, ELKU); 1♀, Nago, Okinawa–jima, 7. IV. 1979 (K. Ohara, ELKU); 1♀, Mt. Katsuu–dake, Nago–shi, Okinawa–jima, 3. XI. 2004 (R. Murao); 6♀3♂, Gima, Kume–jima, 27. V. 2003 (R. Murao); 3♀3♂, Zenda, Kume–jima, 27. V. 2003 (R. Murao, 1♂ illustrated in Figs 4, 14); 1♀, Nakadomari, Kume–jima, 26. V. 2003 (R. Murao).

14–15 Male of Lasioglossum (Lasioglossum) subopacum okinawa Ebmer et Maeta. 16–17 Male of Lasioglossum (Lasioglossum) occidens (Smith) 18–19 Male of Lasioglossum (Lasioglossum) sakishima Ebmer et Maeta 14, 18 labrum 16 first metasomal tergum 15, 17, 19 distal parts of metasomal sternum.

20, 22 Female of Lasioglossum (Lasioglossum) subopacum subopacum (Smith) 21, 23 Female of Lasioglossum (Lasioglossum) subopacum okinawa Ebmer et Maeta 20, 21 hairs on propodeum 22, 23 hairs on first metasomal tergum 24 Distribution of both ssp. subopacum and ssp. okinawa.
I would like to express thanks to Prof. Osamu Tadauchi (Kyushu Univ.) and Dr. Layne Westover (Kyushu Univ.) for their critical reading of early draft. I also express thanks to Emeritus Prof. Yasuo Maeta (Shimane Pref.), and Assoc. Prof. Ryoichi Miyanaga (Shimane Univ.) for loan of valuable specimens. This is a contribution from the Entomological Laboratory, Faculty of Agriculture, Kyushu University, Fukuoka (Ser. 6, No. 103).