





(C) 2010 Santiago R. Ron. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
For reference, use of the paginated PDF or printed version of this article is recommended.
Osteocephalus festae is an Amazonian species recently resurrected from a synonymy with Osteocephalus buckleyi. Because few specimens are known, its morphological variation, diagnostic characters, and distribution are poorly understood. Herein we determine its phylogenetic relationships and provide a complete taxonomic account based on recently collected specimens (adults and larvae) from nine localities in Ecuador and Peru. Osteocephalus festae is most similar to Osteocephalus verruciger from which it differs in having less tuberculate dorsal skin on males, smaller tympanum, and more tooth rows in the oral disk of larvae. A phylogeny based on mitochondrial DNA sequences, genes 12S and ND1, shows that Osteocephalus festae is closely related to Osteocephalus buckleyi, Osteocephalus mutabor and Osteocephalus verruciger. A clade consisting of Osteocephalus festae, Osteocephalus verruciger, and Osteocephalus buckleyi is characterized by stream dwelling tadpoles. Surprisingly, we found paraphyly among Ecuadorian populations of Osteocephalus buckleyi and Osteocephalus verruciger. The causes for paraphyly are unknown but in Osteocephalus buckleyi may result from the existence of cryptic species.
Andes, Amazon, Anura, morphology, Phylogeny, Osteocephalus buckleyi, Osteocephalus festae, Osteocephalus verruciger, tadpole
Osteocephalus is a genus of hylinae frogs (tribe Lophiohylini) distributed in the Amazon Basin and the Guiana Shield (
The holotype of Osteocephalus festae
is an adult female collected at “Valle Santiago”, Provincia Morona
Santiago, Ecuador. After its description, this binomial was largely
ignored until
Recently collected specimens of Osteocephalus
from nine populations from southeastern Ecuador and northeastern Peru,
one of them at a distance of ~30 km from the type locality (Fig. 1), closely resemble the holotype of Osteocephalus festae
and are morphologically and genetically distinctive from other species.
They also seem to be distinctive from the specimens ascribed to Osteocephalus festae by
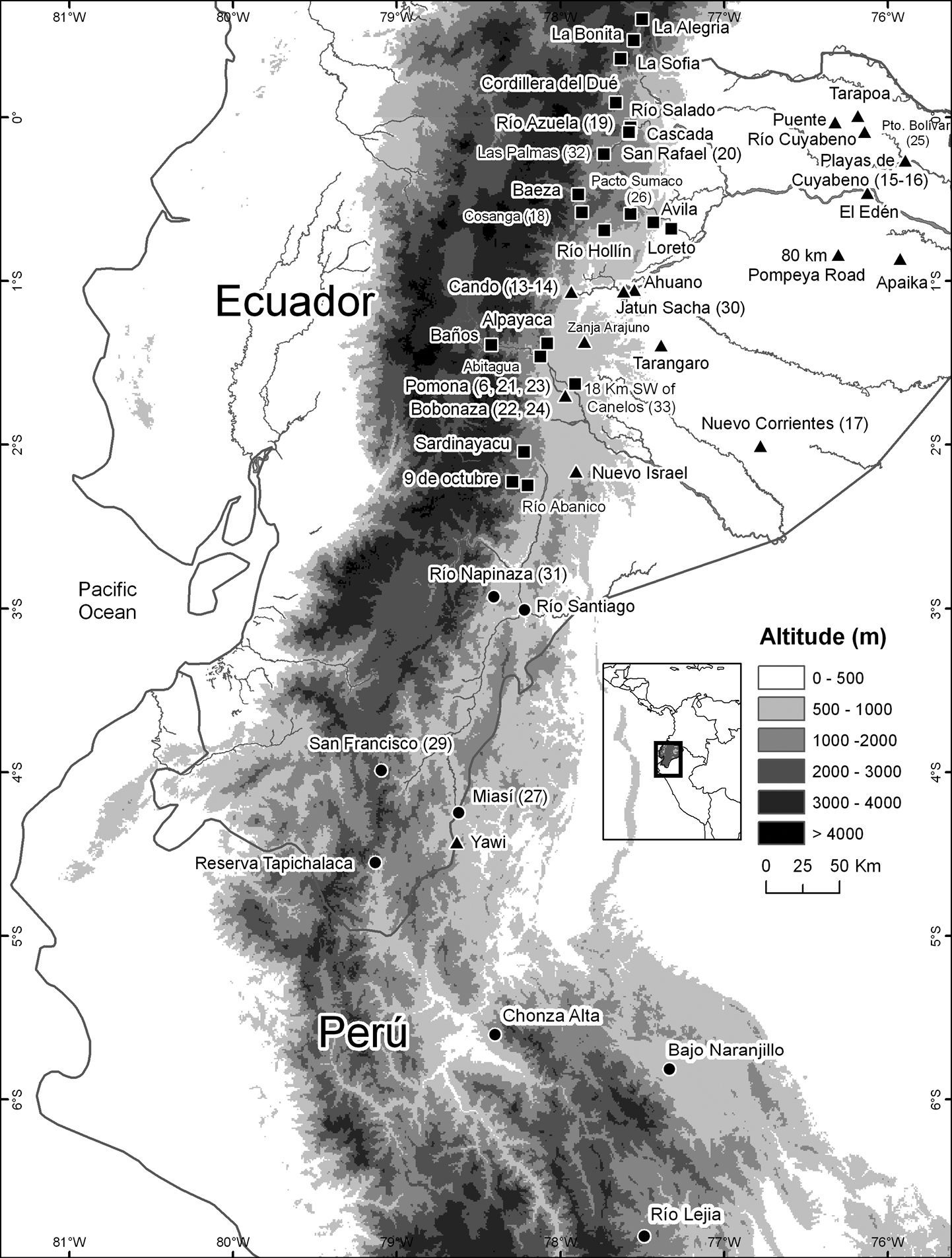
Records of Osteocephalus festae (circles), Osteocephalus verruciger (squares), and Osteocephalus buckleyi (triangles). Locality data from
Total DNA was extracted from muscle or liver tissue
preserved in 95% ethanol and tissue storage buffer using a guanidine
tiocyanate protocol. Polymerase chain reaction (PCR) was used to amplify
the mitochondrial genes 12S rRNA and ND1. We amplified one DNA fragment
for 12S and one or two overlapping fragments for ND1 using primers
listed in
A list of the samples included in the phylogenetic analyses is shown in Table 1. For the outgroup, we included sequences of Osteopilus and Hypsiboas obtained from GenBank (http://www.ncbi.nlm.nih.gov/genbank). Outgroup choice was based on phylogenies showing that Osteocephalus is most closely related to Tepuihyla and Osteopilus (
Specimens used in the phylogenetic analysis. Numbers correspond to those in the Figures 1 and 2.
| Museum No. | Species | No. | Genbank Accession No. | Reference | |
|---|---|---|---|---|---|
| 12S | ND1 | ||||
| SBH 266458 | Hypsiboas heilprini | DQ380357 | EU034080 |
|
|
| USNM 327241 | Osteopilus brunneus | DQ380382 | EU034083 |
|
|
| SBH 266457 | Osteopilus marianae | DQ380383 | EU034086 |
|
|
| SBH 191985 | Osteopilus pulchrilineatus | AY819436 | EU034087 |
|
|
| QCAZ 15981 | Osteocephalus alboguttatus | 1 | HQ600629 | HQ600596 | This study |
| LAC 2216 | Osteocephalus buckleyi | 30 | DQ380378 | EU034082 |
|
| QCAZ 14947 | Osteocephalus buckleyi | 17 | HQ600628 | HQ600595 | This study |
| QCAZ 24446 | Osteocephalus buckleyi | 13 | HQ600633 | HQ600600 | This study |
| QCAZ 24447 | Osteocephalus buckleyi | 14 | HQ600634 | HQ600601 | This study |
| QCAZ 28231 | Osteocephalus buckleyi | 25 | HQ600654 | HQ600621 | This study |
| QCAZ 28277 | Osteocephalus buckleyi | 15 | HQ600639 | HQ600606 | This study |
| QCAZ 28395 | Osteocephalus buckleyi | 16 | HQ600640 | HQ600607 | This study |
| QCAZ 32506 | Osteocephalus buckleyi | 22 | HQ600651 | HQ600618 | This study |
| QCAZ 32508 | Osteocephalus buckleyi | 24 | HQ600652 | HQ600619 | This study |
| QCAZ 37175 | Osteocephalus buckleyi | 23 | HQ600653 | HQ600620 | This study |
| QCAZ 25469 | Osteocephalus buckleyi | 21 | HQ600650 | HQ600617 | This study |
| CORBIDI 623 | Osteocephalus festae | 28 | HQ600649 | HQ600616 | This study |
| QCAZ 38420 | Osteocephalus festae | 31 | HQ600646 | HQ600613 | This study |
| QCAZ 39364 | Osteocephalus festae | 29 | HQ600648 | HQ600615 | This study |
| QCAZ 41039 | Osteocephalus festae | 27 | HQ600647 | HQ600614 | This study |
| QCAZ 20785 | Osteocephalus fuscifacies | 4 | HQ600631 | HQ600598 | This study |
| QCAZ 25603 | Osteocephalus mutabor | 6 | HQ600631 | HQ600598 | This study |
| QCAZ 28223 | Osteocephalus mutabor | 12 | HQ600638 | HQ600605 | This study |
| QCAZ 28646 | Osteocephalus mutabor | 7 | HQ600641 | HQ600608 | This study |
| QCAZ 28647 | Osteocephalus mutabor | 8 | HQ600642 | HQ600609 | This study |
| QCAZ 30926 | Osteocephalus mutabor | 11 | HQ600643 | HQ600610 | This study |
| QCAZ 40253 | Osteocephalus mutabor | 10 | HQ600644 | HQ600611 | This study |
| QCAZ 42999 | Osteocephalus mutabor | 9 | HQ600645 | HQ600612 | This study |
| QCAZ 20797 | Osteocephalus planiceps | 3 | HQ600632 | HQ600599 | This study |
| QCAZ 18230 | Osteocephalus taurinus | 2 | HQ600630 | HQ600597 | This study |
| QCAZ 15942 | Osteocephalus verruciger | 18 | HQ600659 | HQ600626 | This study |
| QCAZ 15991 | Osteocephalus verruciger | 19 | HQ600656 | HQ600623 | This study |
| QCAZ 20544 | Osteocephalus verruciger | 33 | HQ600655 | HQ600622 | This study |
| QCAZ 32032 | Osteocephalus verruciger | 20 | HQ600658 | HQ600625 | This study |
| QCAZ 41108 | Osteocephalus verruciger | 26 | HQ600660 | HQ600627 | This study |
| QCAZ 27816 | Osteocephalus yasuni | 34 | HQ600636 | HQ600603 | This study |
| QCAZ 27998 | Osteocephalus yasuni | 5 | HQ600637 | HQ600604 | This study |
For ease of comparison, we generally follow the format of
Examined specimens (listed in the type-series and Appendix I) are housed at Museo de Zoología, Pontificia Universidad Católica del Ecuador (QCAZ), the Herpetology Collection, Escuela Politécnica Nacional (EPN-H), and the collection of the División de Herpetología, Centro de Ornitología y Biodiversidad (CORBIDI).
Principal Components Analysis (PCA) and Discriminant
Function Analysis (DFA) were used to assess the degree of morphometric
differentiation between adult Osteocephalus buckleyi, Osteocephalus festae, and Osteocephalus verruciger (
Twelve morphometric variables were measured in tadpoles, following
The models with the best fit and the estimated parameters for each of four partitions for the Bayesian analyses are shown in Table 2. The Bayesian analyses of 1975 characters (1152 bp of ND1, 823 bp of 12S) resulted in a consensus tree showing strong support for an Osteocephalus clade (Fig. 2). Within Osteocephalus two clades have strong support: (1) Osteocephalus buckleyi, Osteocephalus festae, Osteocephalus mutabor, and Osteocephalus verruciger, and (2) Osteocephalus alboguttatus, Osteocephalus taurinus, Osteocephalus fuscifacies and Osteocephalus planiceps. A clade excluding Osteocephalus mutabor from (1) is weakly supported (posterior probability = 0.72).
Post burn-in averages for parameters of Bayesian analyses. Abbreviations are: AIC = Akaike information criterion, I = proportion of invariant sites, G = shape parameter of the gamma distribution of rate variation, k = ratio of transition/transversion rates.
| Rate Matrix | Base Frequency | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Partition | Best-fit model | AIC score | I | G | K | AC | AG | AT | CG | CT | GT | A | G | C | T |
| 12S | GTR + G + I | 15762 | 0.386 | 0.740 | - | 0.079 | 0.281 | 0.081 | 0.007 | 0.522 | 0.026 | 0.334 | 0.183 | 0.238 | 0.243 |
| ND1, 1st position | HKY + G + I | 14527 | 0.489 | 1.385 | 14.22 | - | - | - | - | - | - | 0.365 | 0.129 | 0.240 | 0.264 |
| ND1, 2nd position | GTR + G | 6472 | - | 0.185 | - | 0.024 | 0.589 | 0.047 | 0.042 | 0.274 | 0.020 | 0.261 | 0.096 | 0.307 | 0.334 |
| ND1, 3rd position | HKY + G + I | 3745 | 0.295 | 1.617 | 14.22 | - | - | - | - | - | - | 0.397 | 0.104 | 0.216 | 0.280 |
Phylogenetic analysis shows that both Osteocephalus buckleyi and Osteocephalus verruciger are paraphyletic relative to each other. In Osteocephalus verruciger, the population of Pacto Sumaco is more closely related to one of two clades of Osteocephalus buckleyi than to the other populations of Osteocephalus verruciger. Seven populations of Osteocephalus buckleyi are separated in two well-supported clades, one of which is embedded within Osteocephalus verruciger. In contrast, monophyly among populations of Osteocephalus mutabor and Osteocephalus festae is strongly supported. Within Osteocephalus festae, pairwise uncorrected p-genetic distances range from 0.001 (Río Napinaza vs. Miasí) to 0.014 (San Francisco vs. Río Lejía). Uncorrected p-distances between both clades of Osteocephalus buckleyi range from 0.047 to 0.060. Distances between Pacto Sumaco and the other populations of Osteocephalus verruciger range from 0.015 to 0.018.
Systematic account of Osteocephalus festaeThroughout this section, coloration refers to preserved specimens unless otherwise noted. Osteocephalus festae is a medium-sized species of Osteocephalus having the following combination of characters : (1) size sexually dimorphic; maximum SVL in males 56.1 mm, in females 84.9 mm; (2) skin on dorsum bearing tubercles in males, smooth in females; (3) skin on flanks areolate; (4) hand webbing formula II2½—3¼III3½—2IV; foot webbing formula varying as shown in Table 3 and Fig. 4; (5) dorsum brown, usually with irregular dark marks; (6) venter varying from cream to tan, with ill to well defined brown chocolate blotches; (7) narrow, cream to light brown, labial stripe confluent with similarly colored suborbital mark; (8) flanks cream to light brown with darker reticulations anteriorly and dark blotches posteriorly; (9) dermal roofing bones of the skull weakly exostosed; (10) bones green in life; (11) iris dark brown without reticulations, in life; (12) paired vocal sacs located laterally, behind jaw articulation, (13) in life, juveniles with red iris, and pale elbows, knees, and heels; (14) larvae with LTRF of 4/7 or 5/7.
Osteocephalus festae is most similar to Osteocephalus verruciger. Both species differ from other Osteocephalus by the combination of a brown iris (in life) and the presence of brown marks in the venter (in life and preservative). Osteocephalus festae differs from Osteocephalus verruciger in having: (1) less dorsal ornamentation in males (fewer and less developed dorsal tubercles in Osteocephalus festae), (2) smaller tympanum (1/5 of head length in Osteocephalus festae vs. 1/4 in Osteocephalus verruciger), and (3) more tooth rows in larvae (LTRF = 4/7 to 5/7 in Osteocephalus festae vs. 2/5 in Osteocephalus verruciger; Fig. 5). Mitochondrial DNA sequences show that Osteocephalus festae and Osteocephalus verruciger are not sister species (Fig. 2). Osteocephalus festae differs from Osteocephalus mutabor Jungfer & Hödl, 2002 in having chocolate blotches in the venter (blotches absent in Osteocephalus mutabor). It differs from most species of Osteocephalus (except Osteocephalus verruciger, Osteocephalus heyeri Lynch, 2002, and Osteocephalus subtilis
Martins & Cardoso, 1987) in having a dark brown iris in life (iris
bronze to golden with or without black straight lines or irregular
black reticulations in Osteocephalus alboguttatus (Boulenger, 1882), Osteocephalus buckleyi, Osteocephalus cabrerai (Cochran & Goin, 1970), Osteocephalus deridens Jungfer et al., 2000, Osteocephalus exophthalmus Smith & Noonan, 2001, Osteocephalus fuscifacies Jungfer et al., 2000, Osteocephalus leoniae Jungfer & Lehr, 2001, Osteocephalus leprieurii (Duméril & Bibron, 1841), Osteocephalus mutabor, Osteocephalus oophagus Jungfer & Schiesari, 1995, Osteocephalus pearsoni (Gaige, 1929), Osteocephalus phasmatus MacCulloch & Lathrop, 2005, Osteocephalus planiceps Cope, 1874, Osteocephalus taurinus Steindachner, 1862, and Osteocephalus yasuni Ron & Pramuk, 1999). Osteocephalus mimeticus (Melin, 1941) can be discriminated from Osteocephalus festae in having a black iris with golden marks. Osteocephalus festae differs from Osteocephalus subtilis and Osteocephalus heyeri in size (maximum male SVL 38.8 mm in Osteocephalus subtilis, 36.1 mm in Osteocephalus heyeri vs. 56.1 mm in Osteocephalus festae). The presence of areolate skin in the flanks, specially anteriorly, distinguish Osteocephalus festae from Osteocephalus leprieurii, Osteocephalus mutabor, Osteocephalus pearsoni, Osteocephalus planiceps, and Osteocephalus yasuni (smooth to granular skin on flanks; Trueb & Duellman, 1971). Osteocephalus taurinus has weakly areolate skin restricted to the axillary region. Osteocephalus festae further differs from Osteocephalus buckleyi and Osteocephalus cabrerai in lacking prominent tarsal tubercles (

Bayesian consensus phylogram depicting relationships within Osteocephalus. Phylogram derived from analysis of 1975 bp of mtDNA (genes ND1 and 12S). Numbers in parenthesis corresponds to those on Table 1 and Figure 1. Posterior probabilities resulting from Bayesian Markov chain Monte Carlo searches appear above branches. An asterisk represents a value of 1. The outgroup species Hypsiboas heilprini is not shown.
The holotype is an adult female with SVL = 78 mm (Fig. 3G, H; Peracca, 1904). The descriptions provided by

Dorsolateral and ventral views of Osteocephalus festae. A QCAZ 41039, adult female, SVL = 79.81 mm B Amplectant pair (not collected) from Chonza Alta, Peru C–D QCAZ 45674, subadult, SVL = 36.54 mm E–F QCAZ 38081, juvenile, SVL = 13.37 mm G–H MZUT An208 (holotype), adult female, (SVL = 78.00 mm;
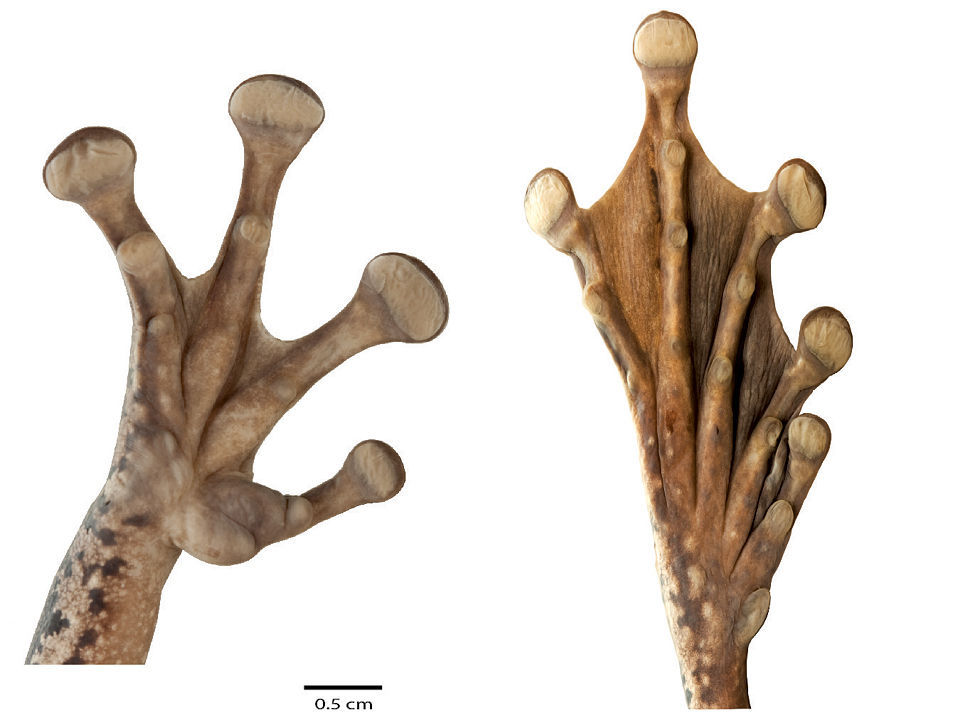
Ventral views of the right hand and foot of Osteocephalus festae. Adult female from Río Napinaza, Ecuador, SVL = 84.93 mm, QCAZ 39811. Hand and foot are shown at the same scale.
Variation in dorsal and ventral coloration of preserved specimens is shown in Figures 6 and 7. Dorsal coloration consists of a light brown to dark brown background with irregular marks. There is sexual dimorphism in dorsal tuberculation: in females the dorsum is smooth while in males it varies between having scant and ill-defined non-keratinized tubercles (most males from Río Napinaza, e.g., QCAZ 26488) to having abundant keratinized tubercles (two males from Chonza Alta, e.g., CORBIDI 758; Fig. 3B).
Ventral surfaces of preserved specimens (Fig. 7) have a cream (QCAZ 39364) to tan (QCAZ 39806) background with darker brown marks that are more distinct and abundant in females (e.g., QCAZ 39811) than in males (e.g., QCAZ 39799); a male from Río Lejía (CORBIDI 623) has an immaculate venter. Ventrally, limbs vary from brown to cream; in QCAZ 39809 and 39811 cream dots are present on hindlimbs; scant cream tubercles can be present in the external edge of the forearm (e.g., QCAZ 39804). The vent region is dark brown to brown bordered by a lighter area (cream to tan). Flanks are areolate in the anterior half and smooth posteriorly. The areolate portion is cream with dark brown reticulation; the posterior half is cream (e.g., QCAZ39810) to light brown (e.g., QCAZ 39806) with dark brown blotches.
Head shape is rounded in dorsal view and rounded (e.g., QCAZ 39803) to bluntly rounded (e.g., QCAZ 39800–01) in lateral view. Lateral head coloration varies between dark brown (QCAZ 11625) to light brown (QCAZ 39810). Except for QCAZ 39802 and 41039, there is a lighter (brown to cream) subocular mark. A tan (QCAZ 39805) to cream (QCAZ 39364) labial stripe is always present. The tympanic annulus is concealed dorsally and has lighter color than the background. Variation in hand and foot webbing is shown in Table 3. The distal subarticular tubercle on Finger IV is single in all specimens.
Variation in webbing in hand and feet of representative adults of Osteocephalus festae. Webbing formula notations follow
| Hand | Foot | |
|---|---|---|
| QCAZ 39805 (female) | I basal II2½—3¼III3½—2+IV | I1–—1–II1–—1–III1–—1–IV1–—1–V |
| QCAZ 39809 (female) | I basal II2½—3¼III3½—2+IV | I1–—1–II1–—2–III1–—2½IV2½—1–V |
| QCAZ 39799 (male) | I basal II2½—3¼III3½—2+IV | I1—1II1—1–III1–—2½IV2½—1–V |
| QCAZ 39802 (female) | I basal II2½—3¼III3½—2+IV | I1+—2–II1+—2III1—2IV2—1–V |
| QCAZ 26304 (female) | I basal II2½—3¼III3½—2–IV | I1–—1–II1–—1–III1–—1–IV1–—1–V |
| QCAZ 26488 (male) | I basal II2½—3¼III3½—2–IV | I1–—1–II1–—1–III1–—1–IV1–—1–V |
Morphometric data pertain to adults and are summarized in Table 4. In the examined series, the largest male has a SVL of 56.09 mm and the largest female 84.94 mm; mean male SVL = 49.47 mm (n = 12; SD = 5.10), mean female SVL = 67.92 mm (n = 27; SD = 10.08). Females are significantly larger than males (t = 5.52, df = 23, P < 0.001). A MANOVA on the residuals of the regressions between SVL and the other measured variables indicates lack of significant differences between sexes in size-free morphometry (F = 1.052, df = 17, P = 0.433).
Descriptive statistics for morphometric measurements of adult Osteocephalus festae. Mean ± SD is given with range below. Bold figures are averages for individuals of all populations. Abbreviations are: SVL = snout-vent length; FOOT = Foot Length; HL = Head Length; HW = Head Width; ED = Eye Diameter; TD = Tympanum Diameter; TL = Tibia Length; FL = Femur Length. All measurements are in mm.
| Species | SVL | FOOT | HL | HW | ED | TD | TL | FL |
|---|---|---|---|---|---|---|---|---|
| Males (n = 12) | 47.47 ± | 19.38 ± | 14.51 ± | 15.77 ± | 5.01 ± | 2.79 ± | 25.90 ± | 22 ± |
| Females (n = 27) | 67.91 ± | 29.79 ± | 20.31 ± | 22.60 ± | 6.25 ± | 3.72 ± | 39.13 ± | 35.35 ± |
| Chonza Alta | ||||||||
| Males (n = 4) | 43.72 ± | 17.97 ± | 14.07 ± | 13.82 ± | 4.51 ± | 2.55 ± | 24.03 ± | 22.05 ± |
| Females (n = 8) | 68.72 ± | 29.33 ± | 20.62 ± | 22.33 ± | 6.2 ± | 3.26 ± | 39.56 ± | 34.63 ± |
| Miasí | ||||||||
| Female (n = 1) | 79.81 | 36.65 | 21.93 | 26.12 | 6.98 | 3.8 | 45.73 | 42.19 |
| Río Lejia | ||||||||
| Males (n = 1) | 48.3 | 20.7 | 14.3 | 16.8 | 4.3 | 2.8 | 26.8 | 24.6 |
| Females (n = 1) | 72.6 | 30 | 21.1 | 23.2 | 6.1 | 3.7 | 41.2 | 30.4 |
| Río Napinaza | ||||||||
| Males (n = 4) | 50.82 ± | 21.08 ± | 14.62 ± | 17.4 ± | 5.66 ± | 2.96 ± | 27.67 ± | 24.88 ± |
| Females (n = 16) | 66.37 ± | 29.59 ± | 20.04 ± | 22.53 ± | 6.28 ± | 3.96 ± | 38.35 ± | 35.55 ± |
| San Francisco | ||||||||
| Female (n = 1) | 69.54 | 29.48 | 19.63 | 21.73 | 5.72 | 3.55 | 39.63 | 35.90 |
| San Carlos | ||||||||
| Males (n = 3) | 47.74 ± | 19.31 ± | 15.00 ± | 15.86 ± | 5.04 ± | 2.87 ± | 25.72 ± | 23.23 ± |
Based on digital photograph of adult female QCAZ 41039 (Fig. 3A): dorsum dark brown with irregular light brown and yellowish green marks; canthal region dark brown with yellowish green subocular mark and labial band; tympanum brown; flanks greenish brown with dark brown reticulation anteriorly and irregular dark brown marks posteriorly; dorsal surfaces of thighs and shanks dark brown with transversal brown bands bordered with light brown; dorsal surfaces of forelimbs dark brown with irregular brown marks; venter light tan with irregular brown marks; bones green; iris dark brown. Female CORBIDI 761 has a predominantly light brown dorsum with irregular brown marks; clear areas on flanks and below the eye and tympanum are light yellow.
There is significant change in coloration between juveniles and adults. The following description is based on a digital photograph of juvenile QCAZ 38081 (Fig. 3E-F). The dorsum beige with black interorbital band and two large medial ovoid black blotches; flanks dark brown; dorsal surfaces of thighs and shanks brown with cream transversal bars; dorsal surfaces of arms cream, dorsal surfaces of forearms brown with cream transversal bars; knees, elbows, and heels cream; anterior half of the venter cream, posterior half light brown; bones green; iris bright red.
Three components with eigenvalues > 1.0 were extracted from the PCA for males (Table 5). The three components accounted for 76.4% of the total variation. The highest loadings for the PCA for males were foot length and tibia length for PC I, head length for PC II, and eye diameter and tympanum diameter for PC III (Table 5). The morphometric space of Osteocephalus festae overlaps with Osteocephalus verruciger (Fig. 8) but only slightly with Osteocephalus buckleyi. There are significant differences in PC scores between Osteocephalus festae and Osteocephalus buckleyi along PC II and PC III (t = 2.46, df = 9, P = 0.035; t = 6.76, df = 13, P < 0.001, respectively) but not along PC I (t = 1.61, df = 7, P = 0.115). There are significant differences only along PC III between Osteocephalus festae and Osteocephalus verruciger, (t = 9.03, df = 9, P < 0.001).
Character loadings and eigenvalues for Principal Components (PC) I–III. The analysis was based seven morphometric variables of adult Osteocephalus. Bold figures indicate highest loadings.
| PCA Males | PCA Females | |||||
|---|---|---|---|---|---|---|
| Variable | PC I | PC II | PC III | PC I | PC II | PC III |
| Foot length | 0.523 | 0.564 | 0.242 | 0.459 | 0.202 | – 0.151 |
| Head length | – 0.243 | 0.647 | 0.121 | – 0.243 | 0.394 | 0.537 |
| Head width | 0.407 | – 0.430 | – 0.208 | 0.196 | 0.315 | – 0.680 |
| Eye diameter | – 0218 | 0.259 | – 0.673 | – 0.061 | 0.648 | – 0.125 |
| Tympanum diameter | – 0.224 | 0.004 | 0.655 | – 0.435 | 0.451 | – 0.016 |
| Tibia length | 0.455 | 0.407 | – 0.020 | 0.519 | 0.096 | 0.285 |
| Femur length | 0.441 | 0.398 | – 0.005 | 0.476 | 0.266 | 0.356 |
| Eigenvalue | 2.570 | 1.744 | 1.030 | 2.722 | 1.517 | 1.382 |
Three components with eigenvalues > 1.0 were extracted from the PCA for females (Table 5). The three components accounted for 80.3% of the total variation. The highest loadings for the PCA for females were tibia length and femur length for PC I, eye diameter and tympanum diameter for PC II, and head width and head length for PC III (Table 5). There is little overlap in the morphometric space of Osteocephalus festae with Osteocephalus verruciger and Osteocephalus buckleyi (Fig. 8). Principal Components scores are significantly different between Osteocephalus festae and Osteocephalus verruciger along PC I and PC III (t = 4.96, df = 21, P < 0.001; t = 4.91, df = 21, P < 0.001, respectively) but not along PC II (t = 0.85, df = 21, P = 0.403). Similarly, Osteocephalus festae and Osteocephalus buckleyi differed along PC I and PC III (t = 4.96, df = 21, P < 0.001; t = 4.91, df = 21, P < 0.001, respectively) but not along PC II (t = 0.85, df = 21, P = 0.403).
In the DFA classification procedure, 24 out of 25 specimens of Osteocephalus festae were classified correctly. The misclassified female (QCAZ 38420) was assigned to Osteocephalus verruciger. The multivariate analyses (PCA and DFA) show morphometric differentiation between Osteocephalus festae and the closely related Osteocephalus verruciger and Osteocephalus buckleyi.
Letters in parenthesis refer to individual
tadpoles on each lot. The following description is based on lot QCAZ
30511 of ten larvae in Stages 25 (A), 26 (B), 31 (C), 32 (D), 33
(E), 34 (F), 35 (G), 39 (H), 40 (I) and 42 (J). Tadpoles were
collected at Río Napinaza by E. E. Tapia and I. G. Tapia on October
2003. These larvae belong to the exotrophic, benthic guild as defined
by
Measurements of tadpoles of Osteocephalus festae (lot QCAZ 30511). Developmental stages, in parentheses, are defined according to
| Stage | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variable | A (25) | B (26) | C (31) | D (32) | E (33) | F (34) | G (35) | H (39) | I (40) | J (42) |
| TL | 30.05 | 29.49 | 37.22 | 35.57 | 34.75 | 34.68 | 39.81 | 40.39 | 40.63 | 40.94 |
| BL | 9.51 | 9.19 | 11.06 | 10.07 | 10.82 | 11.04 | 11.44 | 11.13 | 11.81 | 11.16 |
| BW | 5.74 | 5.68 | 7.25 | 7.05 | 7.23 | 7.10 | 8.11 | 7.50 | 8.46 | 7.12 |
| BH | 4.83 | 4.98 | 5.95 | 5.46 | 5.87 | 5.88 | 6.10 | 6.32 | 6.27 | 4.89 |
| TAL | 20.30 | 20.04 | 26.59 | 24.56 | 24.36 | 23.76 | 27.90 | 28.99 | 28.69 | 29.90 |
| ED | 1.02 | 1.30 | 1.25 | 1.28 | 1.47 | 1.10 | 1.50 | 1.41 | 1.70 | 1.47 |
| ODW | 3.50 | 3.35 | 3.90 | 4.12 | 3.80 | 4.43 | 4.13 | 4.00 | 3.94 | 3.46 |
| IOD | 4.27 | 4.34 | 5.15 | 5.40 | 5.08 | 5.45 | 5.04 | 5.26 | 5.74 | 5.02 |
| IND | 3.49 | 3.50 | 4.01 | 4.19 | 3.75 | 3.94 | 3.98 | 4.12 | 4.01 | 1.62 |
| MTH | 5.30 | 5.37 | 5.88 | 6.10 | 5.85 | 6.05 | 6.70 | 6.62 | 6.75 | 6.06 |
| TMH | 2.06 | 2.00 | 2.57 | 2.50 | 3.26 | 2.79 | 3.07 | 3.07 | 3.45 | 2.44 |
| TMW | 1.60 | 1.46 | 2.70 | 2.12 | 2.30 | 2.50 | 2.97 | 2.62 | 2.78 | 2.56 |
Tail musculature robust, decreasing in size towards tip of tail. Dorsal fin not extending onto body, slightly convex and attaining its maximum height at mid length of tail; tail tip rounded; ventral fin convex, beginning at tail-body junction and tapering gradually towards tail tip. Medial vent tube with both walls attached directly to ventral fin, opening directed posteroventrally. Limbs with subarticular patches. Dorsal body, middle body, supraorbital, infraorbital, posterior supraorbital, and posterior infraorbital lateral lines evident. No glands.
Oral disc anteroventral (Fig. 5B; average transverse width 4.35 mm; 58% of body width), not emarginate, LTRF 4/7; papillae distributed around oral disc; tooth rows complete except for medial gap in row A4; A1 = 304, A2 = 322, A3 = 311, A4 = 108 + 112; P1 = 177, P2 = 235, P3 = 234, P4 = 328, P5 = 284, P6 = 330, and P7 = 177.
In preservative, dorsum brown with darker marks between eyes; dark brown dorsolateral stripes extend from mid-body to base of tail; caudal musculature beige with brown spots (Fig 5); skin of flanks, spiracle, vent tube, fins, and around the eyes transparent; belly and fins transparent with white blotches.
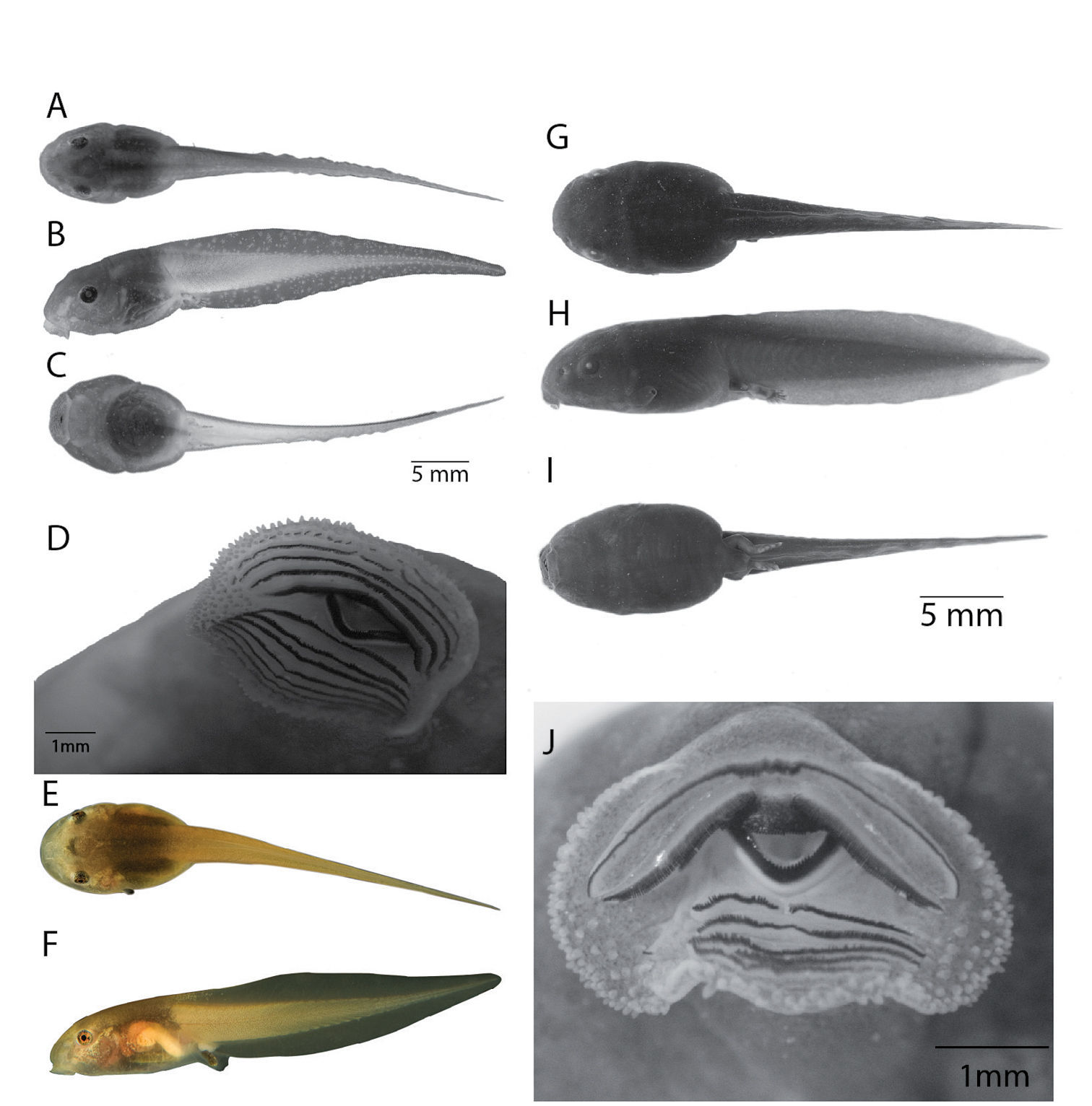
Tadpoles of Osteocephalus festae and Osteocephalus verruciger. A–D, G–J are in preservative; E–F in life. A–D: Osteocephalus festae, stage 39, QCAZ 30511; E–F: Osteocephalus festae, stage 33, QCAZ 38074; G–J: Osteocephalus verruciger, stage 36, QCAZ 36751. A, G: dorsal view; B, H: lateral view; C, I: ventral view; D, J:oral apparatus. Photos in life by SRR.

Adult Osteocephalus festae showing variation in dorsal coloration of preserved specimens. Left to right, upper row: QCAZ 39804, 39811, 39810, 39802, 39798 (females); lower row: QCAZ 39799, 26552, 26488, 26561 (males), 39364 (female). Provincia Loja and Morona Santiago, Ecuador. See Appendix I for locality data.

Ventral views of adult Osteocephalus festae showing variation in ventral coloration of preserved specimens. Specimen identity and arrangement is the same as in Figure 6.
In QCAZ 38074, 26321, 26053, 26498 and 26284 the caudal musculature is cream with brown dots; fins can have dark brown spots without white blotches (e.g., QCAZ 38074, 26321). The LTRF is the same in all stages but in stage 42, rows A2–A4 and P1–P3 have approximately half the number of teeth.
In preservative, ten tadpoles collected in Chonza Alta, Peru, lot CORBIDI-CL-10 in Stages 37 (A-B), 36 (C), 34 (D), 32 (E), 31 (F), 42 (G, H, I) and 44 (J) have dorsum brown with darker marks between the eyes; dark brown dorsolateral stripes from mid-body to base of tail; caudal musculature cream with abundant melanophores; skin of flanks, spiracle, vent tube, and fins transparent; skin around the eyes brown; belly and fins transparent with abundant melanophores. Oral disc with LTRF 5/7; papillae distributed around oral disc; tooth rows complete except for medial gap in row A5.
In life (Fig. 5E-F; QCAZ 38074, stage 33; based on digital photograph), dorsum dark brown with darker marks between the eyes; dark brown dorsolateral lines from mid-body to base of tail; tail musculature light brown with small dark brown melanophores; white dots at tail-body junction; skin is transparent ventrally in anterior half of body and ventrolaterally in the posterior half; tail musculature light brown with dark brown spots; fins transparent. Iris bronze. Live tadpoles from Chonza Alta (CORBIDI-CL-10) have dorsum and caudal musculature olive brown; skin transparent ventrally with bright brown flecks, gut visible through the skin; fins translucent brown.
Larvae of Osteocephalus verruciger differ from those of Osteocephalus festae
(in parentheses) by being smaller, having uniform dark body (tan to
cream), LTRF = 2/5 (4/7 to 5/7); and having a dorsal gap in marginal
papillae (marginal papillae complete). For comparison, larvae of Osteocephalus verruciger is shown in Figure 5 G–H. Differences were verified in 30 Osteocephalus verruciger larvae from four localities (lots QCAZ 1579, 10798, 21405 and 36751). Larvae of Osteocephalus festae have the highest number of tooth rows known among Osteocephalus (LTRF = 2/3 in Osteocephalus oophagus, 2/6 in Osteocephalus buckleyi and Osteocephalus taurinus;
Osteocephalus festaehas
been recorded at nine localities in the Ecuadorian (Loja, Morona
Santiago, and Zamora-Chinchipe provinces) and Peruvian Amazon basin
(Mariscal Cáceres and Rioja provinces). Localities with known elevation
(Río Napinaza, Miasí, San Francisco, Reserva Tapichalaca, Río Lejia,
Chonza Alta, Camñopite Bajo, and Naranjillo) range between 1000 and
2200 m of elevation. The elevation at San Francisco (2200 m) is the
highest known for Osteocephalus. Maximum airline distance between localities is 440 km. Osteocephalus festae and Osteocephalus verruciger have similar elevational ranges and seem to replace each other latitudinally in Ecuador (Fig. 1). Records of Osteocephalus verruciger from Peru (e.g.,
Most of our specimens of Osteocephalus festae
are from Río Napinaza, a river surrounded by secondary forest,
pastures and agricultural lands. At the collection site, the river has
an average width of 2.85 m and an average depth of 23 cm with fast
running water and waterfalls that reach 10 m in height (
All the specimens collected in Las Cataratas de Paraiso (Chonza Alta) and Camñopite were found at night on vegetation 50 to 300 cm above the ground, next to fast running streams. Tadpoles (CORBIDI-CL-10) were found in a rocky stream with average width of 4 m and an average depth of 30 to 40 cm with fast running water, close to the base of a waterfall. At both sites the streams are surrounded by secondary forest, pastures and agricultural lands. The specimens from Bajo Naranjillo and Río Lejia were found at night on branches 150 to 200 cm above the ground (in primary forest at Río Lejia and secondary forest surrounded by pastures at Naranjillo).
At Las Cataratas de Paraiso (Chonza Alta), on 1 December 2007, we found twelve males calling from the low vegetation and six amplectant pairs (Fig. 3B). Recently metamorphosed individuals were perching on leaves and rocks at the shore. On 4 November 2008, at the same stream, we only found one adult non-amplectant female, two adult males, several tadpoles, and 12 freshly metamorphosed juveniles on leaves and rocks (e.g., CORBIDI 1962–64). The rainy season in this region generally starts in December but during our surveys heavy rains fell since the first week of November. Three gravid females (CORBIDI 761, SVL 68.6 mm; CORBIDI 762, SVL 69.1 mm; CORBIDI 764, SVL 66.6 mm) contained 1080, 1388 and 1290 eggs respectively. A gravid female CORBIDI 624 (SVL = 71.2 mm) from Río Lejia, collected on 29 January 2008, contained 780 eggs. The color of eggs in preservative is black and brownish-cream.
Vegetation types for Ecuadorian localities (according to the classification of
Vegetation types of the Peruvian localities (according to
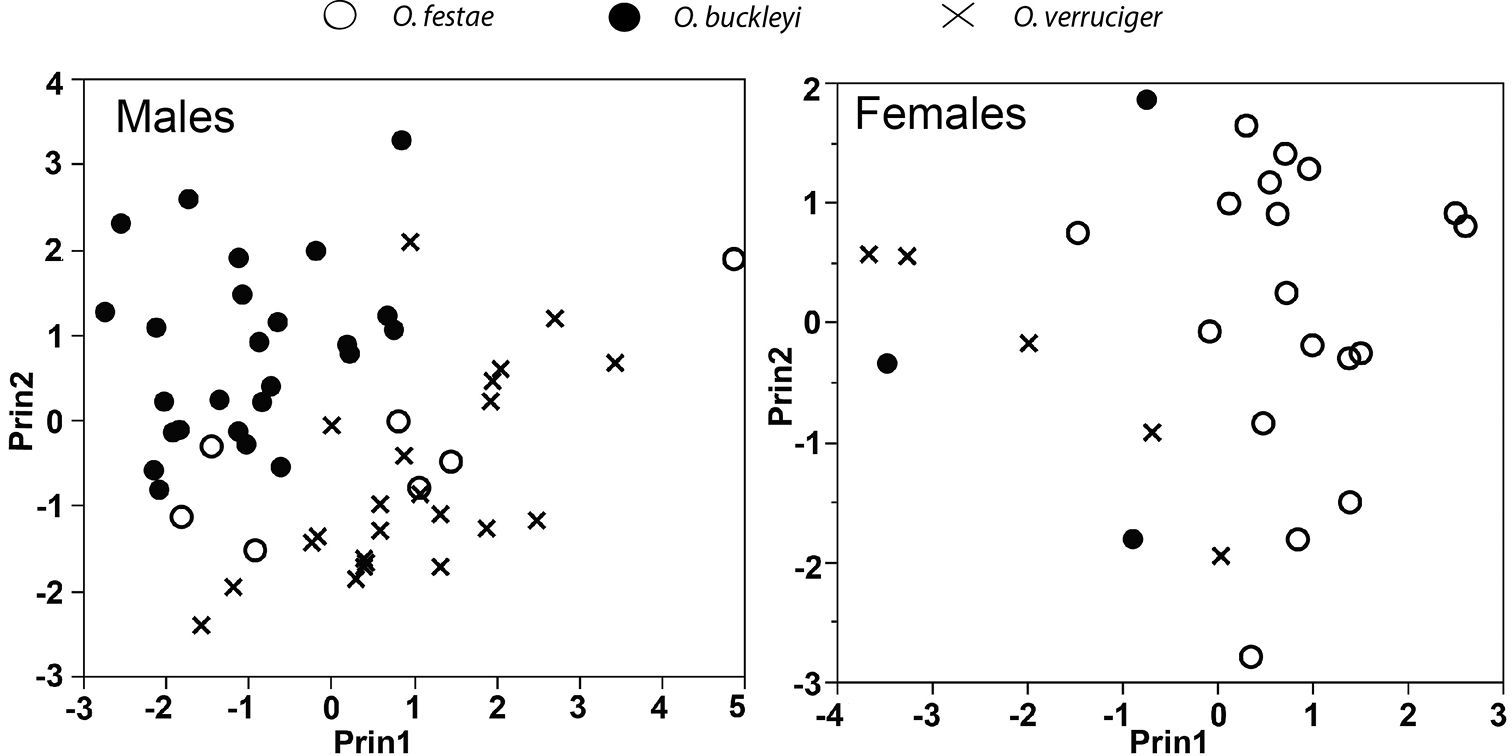
Principal components from analysis of seven size-corrected morphological variables. See Table 5 for character loadings on each component.
The phylogenetic relationships recovered by this study are consistent with the phylogenies reported by
An unexpected result in our phylogeny is the finding of paraphyly among populations of Osteocephalus buckleyi and Osteocephalus verruciger
from Ecuador. Plausible explanations include incomplete lineage
sorting, mitochondrial gene capture, and the existence of cryptic
species hidden within each taxa. In the case of Osteocephalus verruciger, the position in the phylogeny of the population that generates paraphyly, Pacto Sumaco, is weakly supported (Fig. 2) and the genetic distance between Pacto Sumaco and the other Osteocephalus verruciger populations is lower (1.5–1.8% sequence divergence) than the distances between Osteocephalus verruciger and Osteocephalus buckleyi
or between any other species pair in the phylogeny (> 2.1%). In
addition, we could not find conspicuous morphological differences
between Pacto Sumaco and the other Osteocephalus verruciger
populations. The observed pattern suggest that our mitochondrial gene
tree may not correctly reflect the history of divergence among Osteocephalus verruciger and the morphologically distinctive populations of Osteocephalus buckleyi from Hola Vida and Bobonaza. Conversely, the paraphyly among populations of Osteocephalus buckleyi has a strong support with higher genetic distances between both clades (4.7%–6.0%). Neither of these clades represent Osteocephalus cabrerai, a species morphologically similar to Osteocephalus buckleyi for which there are not confirmed records from Ecuador (
This study was supported by grants from the Secretaría Nacional de Ciencia y Tecnología del Ecuador SENACYT (PI-C08-0000470) and Pontificia Universidad Católica del Ecuador. For the loan of specimens we are indebted to A. Almendáriz. William E. Duellman and F. Andreone provided photographs of type material. Ministerio de Ambiente of Ecuador and the Dirección General Forestal y Fauna Silvestre (DGFFS) of Peru issued collection permits. For field assistance we are indebted to P. Peña, D. Salazar-Valenzuela, E. Tapia, I. G. Tapia, and Maik Dobiey. For logistic support in Peru we are indebted to Ronald Wagter. Fieldwork in Peru was supported by the fellowship Koepcke of APECO and Conservation International. Omar Torres-Carvajal helped with translations of relevant literature. Ross MacCulloch and J. J. Wiens provided helpful comments to the manuscript.
Osteocephalus buckleyi.— ECUADOR: PROVINCIA MORONA SANTIAGO: Bobonaza, 660 m (QCAZ 32506–08); PROVINCIA NAPO: Cando, 700 m (QCAZ 24446–47); Ahuano, 410 m (QCAZ 36703); Serena, Río Jatunyacu, 520 m (QCAZ 25321); PROVINCIA ORELLANA: Río Yasuní (QCAZ 7360); Puente del Río Beque, 228 m (QCAZ 43071); Río Rumiyacu, Parque Nacional Yasuní, 250 m (QCAZ 16007); Taracoa, 251 m (QCAZ 34963); Aguarico, Parque Nacional Yasuní (EPN-H 2786); PROVINCIA PASTAZA: Pomona, Fundación Hola Vida, 846 m (QCAZ 25469, 25607, 37175); Nuevo Corrientes, 250 m (QCAZ 14947); Tarangaro, 338 m (QCAZ 39073); Arajuno, Curaray, Río Maderoyacu (EPN-H 6374); PROVINCIA SUCUMBÍOS: Tarapoa (QCAZ 14948); Puerto Bolívar, 240 m (QCAZ 28231); Playas de Cuyabeno, 230 m (QCAZ 28277, 28280, 28395); intersection of the Tarapoa-Puerto Carmen road and Río Cuyabeno, 290 m (QCAZ 28427); PROVINCIA ZAMORA CHINCHIPE: Centro Shuar Yawi, 940 m (QCAZ 31016, 31032–3, 31047, 31051, 31053).
Osteocephalus deridens.—ECUADOR: PROVINCIA ORELLANA: Estación Científica Yasuní, Universidad Católica del Ecuador, Parque Nacional Yasuní (QCAZ 12556).
Osteocephalus festae.—ECUADOR: PROVINCIA LOJA: San Francisco, Arco Iris Reserve, Parque Nacional Podocarpus (3.98845º S, 79.09298º W), 2200 m (QCAZ 39364); PROVINCIA MORONA SANTIAGO: Río Napinaza, 6.6 km N from General Leonidas Plaza (Limón) in the road to Mendez (2.92665º S, 78.40701º W), 1010 m (QCAZ 26283, 26304, 26488, 26552, 26561, 32835, 38081, 38420, 39799, 39804–6, 39798–803, 39808–12); San Carlos, San Miguel and Río Oro, 600–1200 m (QCAZ 11624–26); PROVINCIA ZAMORA CHINCHIPE: Miasí Alto (4.25025º S, 78.61740º W), 1250–1300 m (QCAZ 41039); Reserva Tapichalaca (4.55004º S, 79.12914º W), 1637 m (QCAZ 45674); PERU: REGIÓN DE AMAZONAS: PROVINCIA DE BAGUA: Cataratas de Paraiso-Chonza Alta (5.60264 S, 78.3985 W), 1342 m (CORBIDI 760, 761, 762, 763, 764, 758, 759); Camñopite (5.61469 S, 78.33192 W), 1650 m (CORBIDI 1962, 1963, 1964, 1965, 2992); REGIÓN DE SAN MARTÍN: PROVINCIA MARISCAL CACERES: Río Lejia (6.83655 S, 77.48603 W), 1500 m (CORBIDI 623, 624); PROVINCIA RIOJA: Bajo Naranjillo (5.81571, 77.33668 W), 844 m (CORBIDI 3386).
Osteocephalus fuscifacies.—ECUADOR: PROVINCIA NAPO: El Tena-Talag Road, 15 km from Tena, 550 m (QCAZ 8806); PROVINCIA ORELLANA: Pompeya-Iro Road, 38 km SE from Pompeya (QCAZ 8137); Estación Científica Yasuní, Universidad Católica del Ecuador, 240 m (QCAZ 20785).
Osteocephalus mutabor.—ECUADOR: PROVINCIA NAPO: Chontapuntas, Comunidad Sumak Sacha-Pozo Yuralpa Centro 1 (QCAZ 28646–48); Huino, around the waterfall (QCAZ 30916–17, 30919–20, 30922–23, 30925–26); PROVINCIA ORELLANA: km 22 Pompeya-Iro Road, 287 m (QCAZ 42999); PROVINCIA PASTAZA: Pomona, Fundación Hola Vida, 846 m (QCAZ 25603); Cantón Santa Clara, Río Pucayacu, Colonia Mariscal Sucre (QCAZ 36935, 36946, 40253); PROVINCIA SUCUMBIOS: Puerto Bolívar, 240 m (QCAZ 28223).
Osteocephalus planiceps.—ECUADOR: PROVINCIA DE NAPO: Chontapuntas, Comunidad Sumak Sacha-Pozo Yuralpa Centro 1 (QCAZ 28648); PROVINCIA ORELLANA: Parque Nacional Yasuní, km 38 Pompeya-Iro Road, 280 m (QCAZ 5134, 14842); Estación Científica Yasuní, Universidad Católica del Ecuador, 240 m (QCAZ 14844, 20797–800); PROVINCIA SUCUMBIOS: La Selva lodge, 250 m (QCAZ 7408, 12093–95).
Osteocephalus taurinus.—ECUADOR: PROVINCIA ORELLANA: Parque Nacional Yasuní, km 97 Pompeya-Iro road, 450 m (QCAZ 5301); Estación Científica Yasuní, Universidad Católica del Ecuador, 220 m (QCAZ 9007, 10604, 14804, 14954, 24449–50); PROVINCIA SUCUMBÍOS: Reserva de Producción Faunística Cuyabeno, 220 m (QCAZ 5871–77); Puerto Bolívar, 240 m (QCAZ 27916, 27920); Zábalo, 220 m (QCAZ 27982, 28015); Chiritza-Puerto El Carmen road, intersection with Río Aguas Negras, 270 m (QCAZ 28485); Tarapoa-Puerto El Carmen road, intersection with Río Cuyabeno, 290 m (QCAZ 28435–36); PROVINCIA ZAMORA CHINCHIPE: Shaime, Nangaritza, 980 m (QCAZ 18230).
Osteocephalus verruciger.—ECUADOR: PROVINCIA MORONA SANTIAGO: Nueve de Octubre (QCAZ 32266); Bosque Protector Abanico, Morona (EPN-H 11444–45); Río Sardinayacu, Palora, Parque Nacional Sangay (EPN-H 5940–42, 5947); PROVINCIA NAPO: E of Volcán Sumaco, 1570 m (QCAZ 1560, 1562); Lago Agrio road between Cascabel 1 and 2, 1600 m (QCAZ 7783–84); Sumaco, 1800–2100 m (QCAZ 8964); Pacto Sumaco (QCAZ 10907); km 13 (Loreto-Coca road), 1324 m (QCAZ 22201); Río Hollín (QCAZ 1681, 2405); Coordillera de los Guacamayos, Cosanga-Archidona road, 1600 m (QCAZ 12206, 41108); El Reventador (QCAZ 29208); Cascada San Rafael, 1553 m (QCAZ 363, 13225, 13247, 16954, 32032–36); Cosanga, 339 m (QCAZ 15942); PROVINCIA SUCUMBIOS: Quito-Lago Agrio road, Río Azuela, 1680 m (QCAZ 15149, 15991–97, 16220, 16953, 22497, EPN-H 6341, 11987, 12105–07, 12112, 12143); La Bonita (QCAZ 3175); trail to Volcán Reventador, Gonzalo Pizarro (EPN-H 7052–53, 7059–60).
Osteocephalus yasuni.—ECUADOR: PROVINCIA SUCUMBIOS: Zábalo, 220 m (QCAZ 27998); Playas de Cuyabeno, 230 m (QCAZ 27816).